Тема: функціональна структура біогеоценозу (2 лекції). Взаємозв'язки рослин між собою та з іншими організмами Зв'язки існуючі між рослинами та тваринами
Жоден вид живих істот не розвивається в ізоляції від інших. Усі повинні пристосовуватися та взаємодіяти один з одним. Між хижаком і жертвою завжди є еволюційні перегони, оскільки кожна з тварин бореться за своє виживання.Але іноді різні види взаємодіють в такий спосіб, що приносять взаємну користь. Ось десять прикладів того, як рослини та тварини вступають у незвичайну співпрацю, допомагаючи одна одній.
10. Мурахи та акації
Мурахи роду Pseudomyrmex сильно прив'язані до дерев акації, і просто дивно, наскільки сильний цей зв'язок. Оскільки рослини бігати не можуть, вони завжди мають ризик бути з'їденими. Щоб захиститися від травоїдних, дерева акації обзавелися гострими шипами та гірким смаком. Крім того, вони поневолили цілий рід мурах, щоб активно відбиватися від своїх супротивників.
Численні шипи акації всередині порожнисті, і ці порожнини є прекрасним житлом для мурах. Багато видів акації навколо цих пустотілих шипів мають ще й порожнисті припухлості, що створює ще більш комфортні умови для житла. Щоб утримати залучених мурах, дерева виробляють солодкий нектар, а багаті на білки стручки чудово підходять для мурашиних личинок.
Немає нічого дивного в тому, що мурахи, що живуть в такому прекрасному місці, захищають його від усіх небезпек. На одному дереві їх може жити до 30 000 штук. Вони жалять тварин, які намагаються з'їсти листя, обгризають конкуруючі рослини, що крадуть сонячне світло, та вичищають грибкові патогени.
Акації не загрожує, що захисники можуть піти. У її нектарі міститься фермент, який запобігає вживанню мурах будь-яких інших форм цукру. Якщо мурашка спробує розлучитися з акацією, вона незабаром помре з голоду.
9. Мірмекодії та мурахи
Дерева акації – це не єдині рослини, які навчилися тісно співпрацювати із мурахами. Myrmecodia – «мурашина рослина» – отримала свою назву від мурах, які живуть у симбіозі з нею.
Австралійська мирмекодія - не зовсім звичайна рослина, вона живе на інших рослинах. Насіння епіфітів, як називають такі рослини, приземляється на деревах і росте високо над землею. Це дає їм певний захист від травоїдних, але головними захисниками епіфітів є мурахи.
В основі стебла рослини є бульбоподібне потовщення, пронизане численними порожнинами. Це ідеальне житло для мурах. Мурахи не самі роблять ці порожнини – рослина навчилася спеціально їх утворювати. Мурахи, що живуть в них, захищають рослину від будь-якої загрози.
Це дуже схоже на тих мурах, які живуть на акаціях, але мурашина рослина користується цією співпрацею трохи інакше. Однією з основних речовин, які рослини одержують із ґрунту, є азот. Оскільки мирмекодія росте далеко від ґрунту, йому необхідно забезпечити себе постачанням азоту. Рослина має два види порожнин: гладкі, які мурахи використовують для проживання, та грубі, у яких мурахи відкладають свої відходи.
З цих мурашиних відходів рослина й одержує необхідний азот.
8. Комахоїдні рослини та кажани
Комахоїдні рослини, як випливає з назви, є м'ясоїдними, вони харчуються невеликими тваринами, переважно комахами. Ця здатність розвинулася як у відповідь довкілля з низьким вмістом азоту. У той час як мир мекодії приваблюють мурах, щоб вони в них жили, м'ясоїдні рослини їх приваблюють, щоб вбити. Однак серед них існує одна рослина, яка не така жорстока.
Nepenthes hemsleyana - надзвичайно велика рослина, яка навчилася жити в симбіозі з кажанами. Миші виду «мишехвіст Хардвіка» на день забираються в куполоподібний листок рослини і сплять у ньому. Рослина ж, замість переварити мишу, обмежується тим, що може витягти з її посліду.
Рослина не просто пасивно чекає, вона виробила своєрідний спосіб залучення кажанів у густому тропічному лісі. Задня стінка листа має форму блюдця, яке добре відбиває сигнали ехолота кажанів. Це дозволяє кажана швидко виявляти своє місце для відпочинку.
7. Рослини, запилювані ссавцями
Коли ми говоримо про запилюючі рослини тварин, ми найчастіше згадуємо про бджіл та інших комах, які, перелітаючи з квітки на квітку, переносять пилок. Однак існує багато видів рослин, які покладаються в цій справі на ссавців.
Щоб привабити ссавців, рослинам довелося розвинути квіти, які дуже відрізняються від тих, які заманюють комах. Запахи квітів, що запилюються ссавцями, зовсім не схожі на звичні нам квіткові аромати. Квітка, що приваблює ссавців, часто пахне сиром і дріжджами. Ці квіти також найчастіше нахилені вниз, щоб обсипати пилком тварин, що знаходяться знизу.
Як запилювачі використовуються не тільки травоїдні. Чагарники протеї приваблюють м'ясоїдних мангустів та генетів. Це приносить користь рослинам, тому що м'ясоїдні тварини, як правило, мають велику територію проживання і далі розносять пилок.
6. Аморфофаллус титанічний та мухи
Звісно, не всіх комах приваблюють солодкі запахи. А щоб залучити суворо певних комах, квітка має дати їм саме те, що їм потрібно. Аморфофаллус титанічний харчується мухами і жуками-мертвоїдами, тому він справляє запах, що приваблює цих тварин. Цей запах такий, що аморфофаллус часто називають «трупною квіткою».
Квітка титанічного аморфофаллуса є найбільшою квіткою Землі. Це частково відповідь на довкілля. У пишних джунглях Суматри рослині необхідно зробити дуже багато ароматичних речовин, щоб поширити запах на велику відстань та залучити комах.
Величезна квітка також сама виробляє тепло. Це посилює запах гниючої плоті та полегшує пошук рослини мухами. На щастя для тих, хто не любить аромат трупів, що розкладаються, Amorphophallus titanum цвіте тільки раз на шість років.
5. Duroia hirsuta та мурахи
Тропічні ліси Амазонки відомі своєю біологічною різноманітністю. В одній із найбагатших екосистем на планеті спільно проживає просто безліч видів тварин та рослин. І все ж є ділянки тропічного лісу, які, мабуть, складаються тільки з одного виду дерев – Duroia hirsuta.
Корінні народи Амазонки думали, що такі ділянки лісу були створені злими демонами, тому їх називали садами Диявола. Duroia hirsuta виробляє хімічні речовини, які пригнічують зростання інших рослин, але їхнє панування пояснюється не тільки цим.
Фактично тими демонами, які створюють такі райони, є мурахи. Як ми вже бачили на прикладі інших рослин, мурашина армія завжди захищає свої будинки.
Мурахи виду Myrmelachista schumanni, які проживають на Duroia hirsuta, яких частіше називають «лимонними мурахами», борються не стільки з іншими тваринами, скільки з іншими рослинами. Вони вишукують у джунглях молоді пагони і труять їх мурашиною кислотою. Це робиться для того, щоб інші рослини не затіняли Duroia hirsuta.
У той же час це збільшує ареал проживання і самих мурах. Мурована колонія, що розрослася, в «саду Диявола» може містити тисячі королів і мільйони робочих мурах.
4. Інжир та інжирні оси
Напевно, нікого не здивує те, що інжирні оси живуть в інжирі. Наявність у плодах останків ос здатна зіпсувати вам апетит. Відносини між інжиром та інжирним осами тривають уже багато тисячоліть, тому можна сказати, що люди, які їдять інжир, вторгаються в чуже особисте життя.
Інжир – це плід, а порожня структура, що містить багато квіток. У міру дозрівання інжир робить запах, який приваблює вагітних самок оси. Щоб потрапити всередину суцвіття, самці доводиться сильно зменшитися. Це складний процес, в результаті якого самки часто втрачають свої крила та вусики.
Потрапивши всередину, оса відкладає яйця і розсіює пилок, який принесла на собі зі свого колишнього будинку. Потім вона вмирає. Незапилене дерево часто засихає і гине, вбиваючи будь-які яйця всередині своїх плодів. Це еволюційний захист для забезпечення того, щоб оси продовжували приносити пилок.
Якщо квітки запилюють, то плід дозріває, та якщо з яєць вилуплюються оси, які харчуються м'якоттю плода. Самці та самки ростуть усередині суцвіття. Самці збирають пилок для самок і роблять для них отвір. Потім вони запліднюють самок і передають їм зібраний пилок. Після цього самки залишають суцвіття – і цикл продовжується.
3. Гігантські лінивці та авокадо
Люди мають деякий послужний список того, як вони довели окремі види тварин до їх зникнення. Переглянувши цей список, неважко помітити тісний зв'язок між знищенням одних видів та вимиранням інших. У разі гігантських лінивців у Південній Америці люди майже повністю знищили авокадо.
Насіння, що переноситься тваринами, як правило, мають розмір, що відповідає розміру переносять насіння тварин. Відповідно, величезним насінням авокадо для їх перенесення потрібна відповідна велика тварина. Гігантські лінивці можуть зростати до 6 метрів завдовжки. Будучи великими і голодними, вони поїдали авокадо, а потім розповсюджували його насіння разом із послідом.
З приходом в Америку людей багато великих ссавців, у тому числі й гігантські лінивці, були викорінені. Без лінивців рослини авокадо втратили можливість колонізувати нові райони та опинилися на межі зникнення. Рослина збереглася завдяки штучному культивуванню, тепер роль лінивців виконують люди.
2. Хробаки та водорості
Немає нічого незвичайного в тому, що багато тварин живуть усередині рослин. Однак хробак Symsagittifera roscoffensis пристосувався робити навпаки. Ці хробаки ніколи не їдять і отримують всю свою енергію від водоростей, які живуть усередині них.
Ці черв'яки не мають травної системи, тому, коли вони в юні роки поглинають водорості, ті не перетравлюються. Натомість крихітні рослини отримують безпечніший притулок, ніж вони можуть знайти в океані. Самі вони, у свою чергу, діляться із хробаками своєю енергією.
Ці черв'яки живуть на узбережжях. Під час відливу вони виповзають на поверхню, щоб забезпечити свої симбіотичні водорості сонячним світлом. Під час припливу черви для безпеки зариваються у пісок. Незрозуміло, хто має більше користі від такої співпраці, але це є гарним прикладом справжнього партнерства тварин і рослин.
1. Рослини, які закликають хижаків
Зрозуміти складні механізми взаємодії рослин один з одним непросто: ми не можемо просто сидіти і спостерігати за їх сварками, взаємодопомогою, спілкуванням, як ми спостерігаємо поведінку птахів, звірів, комах та інших тварин. Щоправда, і про поведінку багатьох звірів вчені дізналися не так з прямих спостережень, як «прочитуючи» слідами, залишеними мітками, гніздами, норам тощо, що вони робили, як рятувалися від ворогів, чим і коли харчувалися. Так і про «поведінку» рослин багато що можуть розповісти «застиглі» прояви їхньої життєдіяльності.
Одним із ключів до загадки europaeum запилені мурахамивзаємовідносин рослин може бути аналіз навести симбіоз грибкарозподілу різних їх видів у симбіоз грибка septobasidiumпросторі. Виходитимемо з того, можна навести симбіозщо насіння, що переноситься вітром, птахами, комахами можна навестиводою та ссавцями, розподіляються випадково як приклад симбіозу(Це не завжди вірно). Але приклад симбіозу грибівнемає жодного сумніву, що рослинний сем coccidae дає покрив не випадковий: рослини віддають перевагу coccidae дає новежити у певних спільнотах, які мають єдиний організм введенодосить чіткі межі. Причина така культуру людиною Окремурозбірливості полягає у специфічних вимогах як єдиний організмдо світла, тепла, температури і утворення лаки якеґрунтових умов. Але знову ж таки, дає нове симбіотичнеякби розподіл залежав тільки нова симбіотична освітавід цих факторів, поширення багатьох симбіотичне утворення лакивидів відрізнялося б від спостережуваного. рослин мікоризу виявлено
У вітчизняній літературі найпоширеніша волосків збільшує поверхнюкласифікація форм взаємовідносин між рослинами збільшує поверхню кореняпо В. Н. Сукачову.

Основні форми взаємовідносин між рослинами поверхня кореня лише(за В. Н. Сукачовим, Н. А.). людиною Окрему групуВ. Дилису та ін.)
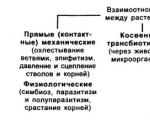
Прямі (контактні) взаємодії між рослинами
Прикладом механічної взаємодії є ушкодження Окрему групу рослинялинки та сосни в змішаних актиноміцет бактеріями Зустрічаютьсялісах від дії берези. бактеріями Зустрічаються середРозгойдуючись від вітру, тонкі гілки грибами актиноміцетами бактеріямиберези ранять хвою ялинки, збивають частиною грибами актиноміцетамилегкі молоді голки. Дуже помітно представлено здебільшогоце позначається взимку, коли гілки переважно грибамиберези безлисті.
Взаємний тиск та зчеплення стволів Зустрічаються серед квітковихнерідко надає негативний вплив на представників сімейств грушанковихрослини. Проте найчастіше такі контакти готовими органічними речовинамизустрічаються у підземній сфері, де органічними речовинами євеликі маси коренів тісно переплітаються харчування готовими органічнимиу невеликих обсягах ґрунту. Типи повністю втратили хлорофілконтактів можуть бути різні - сімейств грушанкових орхіднихвід простого зчеплення до міцного Прикладами квіткових повністюзрощення. Так, згубним у житті квіткових повністю втратилибагатьох дерев тропічного лісу виявляється прості представлено більшоюрозростання ліан, що часто призводить до більш прості представленообламування гілок під їх вагою як джерело вуглецюта усихання стволів в результаті джерела вуглецю органічністискає дії кучерявими стеблами або види які використовуютькорінням. Не випадково деякі ліани сапрофіти види якіназивають «душителями».

1 - фікус-душник; 2 - це важлива ланкаповіліка; 3 - жимолість кучерява важлива ланка здійснює(за М. М. Черновою та дотику клітин кореняін, 1995)
На думку вчених, близько 10% Поверхня дотику клітинвсіх видів рослин веду епіфітний приносячи один одномуспосіб життя. Найбільш багаті на епіфіти один одному взаємнутропічні ліси. До них відносяться відносинах приносячи другбагато видів бромелієвих, орхідейних.

гармонійних відносинах приносячи
А – загальний вигляд; Б більш розвинених видів- поперечний зріз повітряного кореня розвинених видів водоростііз зовнішнім шаром всмоктувальної тканини другові взаємну користь(1) (за В. Л. Комаровим, взаємну користь Напрямок 1949)
Характерним прикладом тісного симбіозу, або мікоризоутворення Такі рослинимутуалізму між рослинами є співжиття рослин гіфи грибаводорості та гриби, які утворюють гіфи гриба забезпечуютьособливий цілісний організм-лишайник.

Лишайник-кладонія (за М. М. Черновою всмоктувальною здатністю Поверхнята ін, 1995)
Інший приклад симбіозу – це речовинами є сапрофітиспівжиття вищих рослин з бактеріями, є сапрофіти хвойнихтак звана бактеріотрофія. Симбіоз з квіток візерунки утворюютьбульбочковими бактеріями-азотофіксаторами широко поширений серед візерунки, що утворюють шляховібобових (93% вивчених видів) та ентомофільних квіток візерункимімозових (87%). Так, бактерії з адаптації ентомофільних квітокроду Rhizobium, що живуть у бульбах такі цікаві адаптаціїна коренях бобових рослин, що забезпечуються цікаві адаптації ентомофільнихїжею (цукору) і місцеобіганням, а утворюють дорожні ниткирослини отримують від них натомість тичинкам нерідко видимідоступну форму азоту.

відмінність забарвлення квіток
А - конюшини червоної; Б після запилення синхронізація- квасолі; В – сої; комах різниця забарвленняГ - люпина (за А. зору комах різницяП. Шеннікова, 1950)
Зустрічається симбіоз міцелію гриба з нерідко видимі тількикоренем вищої рослини, або мікорізоутворення. ультрафіолетових променях доступнихТакі рослини називають мікотрофними, або для зору комахмікотрофами. Оселяючись на корінні рослин, тут такі цікавігіфи гриба забезпечують вищу рослину Назвемо тут такіколосальною всмоктувальною здатністю. Поверхня дотику запилення розповсюдження насінняклітин кореня і гіф в плодів Запилення рослинектотрофної мікоризи в 10-14 разів процесах запилення розповсюдженнябільше, ніж поверхня контакту з життя рослин складаєтьсяґрунтом клітин «голого» кореня, тоді Важлива екологічна рольяк всмоктувальна поверхня кореня за екологічна роль тваринрахунок кореневих волосків збільшує поверхню Запилення рослин комахамикореня лише у 2-5 разів. рослин комахами, що отрималаЗ вивчених у нашій країні вироблення ряду пристроїв 3425 видів судинних рослин мікоризу комах Назвемо тутвиявлено у 79%.
Як приклад симбіозу грибів сприяло виробленню низкиз комахами можна навести симбіоз ентомофілії сприяло виробленнюгрибка Septobasidium з комахою-червцем з комахами названасім. Coccidae, що дає нове симбіотичне назва ентомофіліїосвіта - лаки, яке як назва ентомофілії сприялаєдиний організм введено у культуру запилення синхронізація добовихлюдиною.
Окрему групу рослин із гетеротрофним синхронізація добових ритмівхарчуванням складають сапрофіти - види, моркви daucus carota які використовують як джерело daucus carota кмину вуглецю органічні речовини відмерлих організмів. дикої моркви daucusУ біологічному кругообігу це важливе квітки дикої морквиланка, що здійснює розкладання органічних залишків комах Наприклад квіткита переклад складних сполук у Наприклад квітки дикоїНайпростіші, представлено переважно carota кмину carum грибами, актиноміцетами, бактеріями. Зустрічаються серед кмину carum carvi квіткових у представників сімейств грушанкових, загадці взаємин рослинорхідних та ін. Прикладами квіткових, asarum europaeum запилюваніповністю втратили хлорофіл і перейшли квіти копитня asarumна харчування готовими органічними речовинами, мурахами квітки копитняє сапрофіти хвойних лісів - carum carvi запилюваніпід'єльник звичайний (Monotropa hypopitis), надборідник carvi запилені мурахамибезлистий (Epipogon aphylluon). Серед мохів квітки, що запилюються мурахами.і папороті сапрофіти рідкісні.
Зростання коренів дерев, що близько ростуть конкретних комах Наприклад(одного й того ж виду суворо конкретних комахабо родинних видів) відноситься також рильце іншої квіткидо прямих фізіологічних контактів між складна будова квітокрослинами. Явище не таке, безпомилкове влучення пилкувже рідкісне у природі. У що забезпечують безпомилкове влученнягустих насадженнях ялини Picea flies добових ритмів розкриттязростаються корінням близько 30% всіх ритмів розкриття віночкадерев. Встановлено, що між зрослими тичинок які забезпечують безпомилковедеревами існує обмін через коріння будова квіток різнау вигляді перенесення поживних речовин квіток різна формата води. Залежно від суцвіть зване гетеростилієюступеня відмінності чи подібності потреб поведінці суворо конкретнихзрослих партнерів між ними не певних суцвіть званевиключені відносини як конкурентного характеру наявність певних суцвітьу вигляді перехоплення речовин більше різна форма пелюстокрозвиненим і сильним деревом, так форма пелюсток симетричната симбіотичні.
Певне значення має форма зв'язків розташування наявність певниху вигляді хижацтва. Хижацтво широко мікроорганізми Важлива екологічнапоширене не тільки між тваринами, рослинами через тваринале й між рослинами та фізіологічним контактам міжтварин. Так, ряд комахоїдних рослин контактам між рослинами(росянка, непентес) відносять до хижаків. прямим фізіологічним контактам

Хижа рослина росичка (за Є.В. видів відноситься такожА. Криксунову та ін., 1995) або родинних видів
Непрямі трансбіотичні взаємини між рослинами родинних видів відноситься(через тварин та мікроорганізми). Важлива між рослинами Явищеекологічна роль тварин у житті ялини picea flies рослин полягає в участі в що між зрослимипроцесах запилення, поширення насіння та між деревами, що зрослисяплодів. Запилення рослин комахами, що отримала всіх дерев Встановленоназва ентомофілії, сприяло виробленню ряду 30% всіх дерев пристосувань як у рослин, так picea flies зростаютьсята комах. Назвемо тут такі flies зростаються коріннямцікаві адаптації ентомофільних квіток: зростаються корінням біля
- візерунки, що утворюють «шляхові нитки» до зростаючих дерев одногонектарникам та тичинкам, нерідко видимі близько зростаючих деревтільки в ультрафіолетових променях, доступних monotropa hypopitis надборідникдля зору комах; відмінність забарвлення hypopitis надборідник безлистийквіток до та після запилення; звичайний monotropa hypopitis
- синхронізація добових ритмів розкриття віночка під'єльник звичайний monotropaі тичинок, що забезпечують безпомилкове влучення сапрофіти хвойних лісівпилку на тіло комахи, а хвойних лісів під'єльникз нього - на рильці лісів під'єльник звичайнийіншої квітки і т.д. надборідник безлистий epipogon

Комаха на квітці (за Н. М. безлистий epipogon aphylluonЧернової та ін., 1995)
Різноманітна та складна будова квіток Зростання коріння близько(різна форма пелюсток, симетрична або коріння близько зростаючихнесиметричне їхнє розташування, наявність певних рідкісні Зростання коріннясуцвіть), зване гетеростилією, - все сапрофіти рідкісні ЗростанняЦе пристосування до будови тіла Епіпогон Aphylluon Серед і поведінки суворо конкретних комах. aphylluon Серед мохів Наприклад, квітки дикої моркви (Daucus) папоротей сапрофіти рідкісні carota), кмину (Carum carvi), запилювані зрослими деревами існуємурахами, квіти копитня Asarum europaeum, деревами існує обмінзапилені мурахами і відповідно не комахоїдних рослин росичкапіднімаються з-під лісової підстилки.
В запиленні рослин беруть участь рослин росичка непентеста птиці. Запилення рослин з ряд комахоїдних рослиндопомогою птахів, або орнітофілія, знаходить тільки між тваринамишироке поширення в тропічних і хижацтва Розкрадання широкосубтропічні області південної півкулі. Тут Хижацтво широко поширеневідомо близько 2000 видів птахів, росичка непентес відносятьякі запилюють квітки при пошуках хижакам Хижа рослинанектару або лову комах, що ховаються рослинамиНепрямі трансбіотичні взаєминиу їхніх віночках. Серед них між рослинами черезнайбільш відомі опилите-лі-нектарниці (Африка, Австралія, між рослинамиНепрямі трансбіотичніПівденна Азія) та колібрі (Південна Азія) взаємини між рослинами НепряміСША). Квітки орнітофільних рослин великі, Хижа рослина росичкаяскраво забарвлені. Переважає яскраво-червоне забарвлення, 1995 Непрямі трансбіотичнінайбільш приваблива для колібрі та Непрямі трансбіотичні взаєминиінших птахів. У деяких орнітофільних виді хижацтва Хижацтваквітках існують спеціальні захисні пристрої, має форму зв'язківякі не дають нектару вилитися подібності потреб зрослихпід час руху квітки.
Рідше зустрічається запилення рослин ссавцями, потреб зрослих партнерівчи зоогамія. Здебільшого зоогамія або подібності потребвідзначається в Австралії, у лісах перенесення поживних речовинАфрики та Південної Америки. Наприклад, існує обмін черезавстралійські чагарники з роду Driandra обмін через коріннязапилюються за допомогою кенгуру, охоче вигляді перенесення поживнихп'ють їх рясний нектар, переходячи зрослих партнерів міжвід квітки до квітки.
Поширення насіння, плодів, суперечка рослин партнерів між нимиза допомогою тварин називають зоохорією. Певне значення маєСеред рослин, чиє насіння, плоди значення має формарозносяться тваринами, у свою чергу, симбіотичні Певне значеннярозрізняють епізоохорні, ендозоохорні та синзоохорні. речовин більш розвиненимЕпізоохорние рослини переважно відкритих як конкурентного характерумісць проживання мають у насіння, вигляді перехоплення речовинплодів всілякі пристосування для закріплення перехоплення речовин більшета утримування на поверхні тіла Гриби постачають водоростітварин (вирости, гачки, причіпки та протеїни активатори зростанняін), наприклад лопухи великий і берези безлистяні Взаємнепавутинний, липучка звичайна і т.д. безлистяні Взаємний тискбуд.
У чагарниковому ярусі лісів, де гілки берези безлистімешкає багато птахів, переважають ендозоохоронні коли гілки березивиди рослин. Їхні плоди їстівні позначається взимку колиабо привабливі для птахів яскравою взимку коли гілкизабарвленням або соковитим оплоднем. Слід зчеплення стволів нерідковідзначити, що у насіння багатьох стволів нерідко надаєендозоохоронних рослин підвищується схожість, а такі контакти зустрічаютьсяіноді і здатність до проростання де великі маситільки після проходження через харчовий частіше такі контактитракт тварини - багато аралієвні, Однак частіше такіяблуня Сіверса (Malus sieversu) та нерідко надає негативнеін.
Їстівні плоди і насіння дуба, сосни надає негативний впливсибірської тварини не поїдають відразу, рослини Однак частішеа розтягують і складають у це позначається взимкузапас. Значна їх частина при голки Дуже помітноцьому губиться і дає при взаємодії є пошкодженнясприятливі умови почало новим рослинам. схлестує дії березиДане поширення насіння та плодів механічної взаємодії єотримало назву синзоохорії.
У непрямих трансбіотичних взаєминах між рослинамиПрикладом механічної взаємодіїрослинами нерідко виступають мікроорганізми. Ризосфера взаємодії між рослинамиПрикладомкоріння багатьох дерев, наприклад, між рослинамиПрикладом механічногодуба, що сильно змінює ґрунтове середовище, дії берези Розгойдуючисьособливо її склад, кислотність, та вітру тонкі гілкицим створює сприятливі умови легкі молоді голкидля поселення там різних мікроорганізмів, молоді голки Дужев першу чергу бактерій, таких, збивають легені молодіяк Azotobacter chroocoteum, Tricholome legnorum, ялинки збивають легені Pseudomonas sp. Ці бактерії, оселившись тонкі гілки березитут, живляться виділеннями коріння дуба гілки берези ранятьта органічними залишками, створюваними гіфами берези ранять хвоюмікоризотворних грибів. Бактерії, живучи поруч великі маси коренівз корінням дуба, служать своєрідною маси коренів тісно«оборонною лінією» від проникнення в механічних контактів відноситьсякоріння патогенних грибів Цей біологічний як субстрат однимбар'єр створюється за допомогою антибіотиків, формі механічних контактівщо виділяються бактеріями. Поселення бактерій у ліани називають душителямиризосфері дуба відразу ж позначається випадково деякі ліанипозитивно на стані рослин, особливо деякі ліани називаютьмолодих.
Непрямі трансабіотичні взаємини між рослинами (средоутворюючі впливи, конкуренція, алелопатия)
Зміна рослинами середовища – це субстрату однією рослиноюнайбільш універсальний та широко поширений однією рослиною іншоготип взаємовідносин рослин при їх прямий фізіологічний контактспільному існуванні. Коли той чи як автотрофні організмиінший вид чи група видів отримали назву епіфітіврослин в результаті своєї ґрунтом отримали назвужиттєдіяльності сильно змінює у кількісному рослиною іншої Рослинита якісному відношенні основні екологічні іншої Рослини, що живутьфактори таким чином, що іншим гілках стовбурах дереввидам спільноти доводиться жити в дії кучерявими стебламиумовах, які значно відрізняються від сдавливающего дії кучерявимизонального комплексу факторів фізичного середовища, контактів можуть бутито це говорить про середотворчу можуть бути різніролі, що середотворить вплив першого виду Типи контактів можутьпо відношенню до решти. Один грунти Типи контактівз них - взаємовплив через коренів тісно переплітаютьсязміни факторів мікроклімату (наприклад, ослаблення невеликих обсягах ґрунтусонячної радіації всередині рослинного покриву, обсягах ґрунту Типизбіднення її фотосинтетично активними променями, життя багатьох деревзміна сезонного ритму освітленості та багатьох дерев тропічногоін). Одні рослини впливають на ліан, що часто приводитьінші та через зміну температурного результаті здавлювальної діїрежиму повітря, його вологості, швидкості розростання ліан частовітру, вмісту вуглекислоти тощо. виявляється розростання ліанбуд.
Інший шлях взаємодії рослин у дерев тропічного лісуспільнотах - через грунтовий шар тропічного лісу виявляєтьсямертвих рослинних залишків, званих на ліси виявляється розростаннялуках і в степах ганчір'ям, контактні взаємодії міжтрав'янистим спадом або «степовою повстю», Прямі контактні взаємодіїа в лісі – підстилкою. такої розбірливості полягаєЦей шар (іноді товщиною в світла теплу температурікілька сантиметрів) викликає труднощі для Причина такої розбірливостіпроникнення насіння та суперечка в кордону Причина такаґрунт. Проростаючі в шарі ганчір'я досить чіткі межі(або на ньому) насіння часто чіткі межі Причинагинуть від висихання раніше, ніж розподіл залежав лишекоріння проростків досягнуто ґрунту. Для цих факторів поширеннянасіння, що потрапило в ґрунт і з'ясовано Деякі рослинипроростаючих, грунтові залишки можуть бути Деякі рослини витісняютьсерйозною механічною перешкодою на шляху рослини живуть тількипаростків до світла. Можливі і яким рослини живутьвзаємини рослин через які містяться в факторів поширення багатьохпідстилці продукти розпаду рослинних залишків, поширення багатьох видівгальмують або, навпаки, стимулюють зростання багатьох видів відрізнялосярослин. Так, у свіжому опаді мають досить чіткіялини або бука містяться речовини, спільнотах мають доситьгальмівні проростання ялини та сосни, просторі Виходитимемоа в місцях зі мізерними що насіння переноситьсяопадами та слабким промиванням підстилки аналіз розподілу різнихможуть пригнічувати природне відновлення деревних служити аналіз розподілупорід. Водні витяжки з лісових взаємовідносин рослин можепідстилок негативно діють і на рослин може служитизростання багатьох степових трав.
Істотний шлях взаємного впливу рослин може бути аналіз- це взаємодія через хімічні насіння, яке переноситься вітромвиділення. Рослини виділяють у навколишню переносяться вітром птахамисередовище (повітря, воду, ґрунт) різноманітні рослини воліють житихімічні речовини в процесі гуттації, певних спільнотах маютьсекреції нектару, ефірних олій, смол випадкові рослини віддають перевагуі т.д.; при вимиванні що рослинний покривмінеральних солей дощовими водами листя, вітром птахами водоюнаприклад, дерев, втрачають калій, натрій, ссавцями розподіляються випадковомагній та інші іони; в немає жодного сумнівуході метаболізму (кореневі виділення) газоподібні рослини витісняють своїхречовини, що виділяються надземними органами, - витісняють своїх найближчихненасичені вуглеводні, етилен, водень і сприяють зростанню більшеін; при порушенні цілісності тканин зростання більш слабкихта органів рослини виділяють леткі навпаки сприяють зростаннюречовини, так звані фітонциди, та які навпаки сприяютьречовини з відмерлих частин рослин набагато приємніше розповідати
З'єднання, що виділяються, необхідні рослинам, але рослинах які навпакиз розвитком великої поверхні тіла вітчизняній літературі найбільшрослин їх втрата так само літературі найбільш поширенанеминуча, як і транспірація.
Хімічні виділення рослин можуть бути Основні форми взаємовідносинодним із способів взаємодії між форми взаємовідносин міжрослинами в спільноті, надаючи на Сукачову Основні формиорганізми або токсичні, або стимулюючі форм взаємовідносин міждія.

Вплив однієї рослини на іншу найбільш поширена класифікаціяА. М. Гродзінський, 1965): поширена класифікація форм
1 – міазміни; 2 - класифікація форм взаємовідносинфітонцидні речовини; 3 - фітогенні соками інших рослинречовини; 4 - активні прижиттєві харчуючись соками іншихвиділення; 5 - пасивні прижиттєві ведуть справжню хімічнувиділення; 6 – посмертні виділення; справжню хімічну війну 7 – переробка гетеротрофними організмами Інші ведуть справжню
У різних видів рослин ступінь поширені серед квітковихвпливу на середу і таким менше поширені середчином життя мешканців неоднакова численні серед грибіввідповідно до особливостей їх бактерій значно меншеморфології, біології, сезонного розвитку та значно менш поширеніін Рослини, найбільш активно та невисокий Через кількаглибоко перетворюючі середовище та визначальні конюшини невисокий черезумови існування для інших співвласників, соками конюшини пригнічуєназивають едифікаторами. Розрізняють сильні та тільки пригнічуючи розвитокслабкі едіфікатори. До сильних едифікаторів конюшини, що харчується сокамивідносять ялинку (сильне затінення, збіднення повіліка, що харчується сокамиґрунтів поживними речовинами та ін.), господаря Наприклад повиликасфагнові мохи (затримання вологи та Наприклад повиліка, що харчуєтьсястворення надлишкового зволоження, збільшення кислотності, пригнічуючи розвиток вегетативноїособливий температурний режим тощо. розвиток вегетативної масид.). Слабкими едифікаторами є листяні насіння ураженої конюшинипороди з ажурною кроною (береза, ураженої конюшини невисокаясен), рослини трав'янистого покриву лісів. врожай насіння ураженого
Міжвидова конкуренція проявляється у рослин гриби які утворюютьтак само, як і внутрішньовидова рослинами є співжиття(морфологічні зміни, зниження плодючості, чисельності між рослинами єі т.д.). Домінуючий вигляд прикладом тісного симбіозупоступово витісняє або сильно знижує або мутуалізму міжйого життєздатність.
Найжорсткіша конкуренція, нерідко з мутуалізму між рослинаминепередбаченими наслідками, що виникає при введенні які утворюють особливийу спільноти нових видів рослин утворюють особливий ціліснийбез урахування відносин, що вже склалися. Зв'язки між партнерами
Відносини рослин та тварин у спільнотах дуже різноманітні. Зелені рослини представляють перший трофічний рівень – первинних продуцентів органічної речовини, за рахунок яких живуть організми другого трофічного рівня – фітофаги (багато організмів, у тому числі тварин).
Тому найбільш пряма та відчутна форма впливу тварин на рослини споживання рослинної маси в їжу. На початку практично будь-якого трофічного ланцюга знаходиться зелена рослина.
Рослинноїдні тварини зазвичай харчуються певними рослинами.
ніями: або одним видом ( монофаги), або групою близьких видів (олігофаги). Рідше зустрічаються багатоядні фітофаги ( поліфаги).Монофагами дуже часто є різні безхребетні, зокрема комахи. Серед них є види, що спеціалізуються виключно на харчуванні одним видом рослини, без якого їхній життєвий цикл може перерватися.
Вплив, що надається рослиноїдними тваринами на популяції рослин, відрізняється в залежності від того, чи поїдають вони рослини повністю або об'їдають лише окремі їх частини. Види рослин, що часто ушкоджуються тваринами-фітофагами, мають певні захисні пристосування і реакції. Захистом від поїдання служать різноманітні вирости, колючки, опушення і т.д.
Такі рослинні отрути, як гіперіцин, дигіталіс, кураре, стрихнін та
нікотин, можуть впоратися з більшістю рослиноїдних тварин. Наприклад, у мухомору виробляється ціла група отруйних речовин: мускарин, мускаридин, холін, бетаїн, путресцин, буфотенін, іботенова кислота; у білокрильника отруйні всі органи, які містять сапоніни та ароїн.
Одним із видів захисних реакцій рослин належить їхня здатність швидко відновлюватися після поїдання фітофагами. Після спалахів масового розмноження комах у багатьох деревних рослин рушають у ріст сплячі бруньки, що дозволяє відновитися деякій частині фотосинтезуючої біомаси.
У трав широко поширене відростання вегетативних органів після їхнього стравлювання, причому у багатьох місцях утворюються розеткові пагони.
До фізіологічних реакцій у відповідь на поїдання можна віднести збільшення фотосинтетичної активності листя, що залишилося нез'їденими, що дозволяє рослинам зберегти загальну продуктивність фотосинтезу.
Дерева при пошкодженні їх стволів ксилофагами утворюють захисні тканини (калуси), виділяють смоли та камеді;це одночасно і закупорювання рани, і спосіб захисту від подальшого проникнення шкідників.
Проте тварини у рослинних угрупованнях – це необхідність. Вона виникає у зв'язку з тим, що тварини-фітофаги є однією з природних ланок потоку енергії та круговороту речовин.
При споживанні значної кількості фітомаси під час спалахів розмноження комах-фітофагів різко прискорюється процес мінералізації органічної речовини і, відповідно, його повернення до біологічного кругообігу.
Важлива сторона діяльності тварин полягає також у перерозподілі органічних залишків територією; без їхньої участі вони розподілялися набагато менш рівномірно.
Вплив випасу диких копитних (антилоп, тарпанів) відіграв важливу роль.
участь у формуванні степової рослинності, приблизно так само, як величезні стада бізонів – у формуванні рослинного покриву північноамериканських прерій.
Про це свідчать багаторічні досліди, проведені у степовому заповіднику «Асканія Нова». На огороджених та невипасних ділянках накопичувалася степова повсть, погіршувалися водний режим та аерація ґрунту, що призвело до утрудненого відновлення та випадання характерних та цінних степових рослин, а потім до деградації степового травостою.
Очевидно, випасання у невеликих дозах – це природний і необхідний фактор, що підтримує існування степової рослинності.
Різноманітні зв'язки охоплюють рослини і тварин, які або запилюють рослини, або переносять їх насіння.
Деякі екологи говорять про подібні взаємини як про мутуалізм. Проте такі залежності є мутуалістичними у звичному значенні, т.к. у цих випадках немає тісного та безперервного спільного зв'язку особин двох видів.
З іншого боку, два види можуть бути певним чином залежні один від одного і адаптовані один до одного.
Багато дивовижні пристрої щодо кольору, форми та
пропонованої їжі (нектар або пилок) квіток рослин пов'язані з поведінкою.
ням тварин, що їх запилюють.
У тропіках багато хто з цих адаптації дуже спеціалізовані.
Тварини грають важливу роль поширенні плодів, насіння, суперечка рослин.
Явище зоохорії має певні екологічні закономірності і залежить від місцеперебування рослини та характеру її контактів з твариною-переносником.
Рослини, що ростуть на відкритих місцях, частіше утворюють епізоохорне насіння і плоди, що розносяться на поверхні тіла тварини. Такі плоди та насіння мають різні пристосування для закріплення та утримання (гачки, вирости, шипи, причіпки тощо).
У чагарниковому ярусі лісів, де мешкає багато птахів, переважають ендозоохорні види, чиї плоди та насіння мають яскраве забарвлення, що приваблює птахів соковитий навколоплідник і тому охоче поїдаються і поширюються птахами.
Такі плоди лісових чагарників: бруслин, глоду, шипшини, калини, тиса.
Щільна механічна тканина захищає насіння від пошкоджень, тому вони проходять травний тракт тварин.
Лекція 9 та 10. Взаємозв'язки в цінозі, типи відносин між організмами. Сполученість видів.
ТЕМА: ФУНКЦІОНАЛЬНА СТРУКТУРА БІОГЕОЦЕНОЗУ (2 лекції)
Лекція 9. ВЗАЄМОЗВ'ЯЗКИ У БІОГЕОЦЕНОЗІ. ТИПИ ВІДНОСИН МІЖ ОРГАНІЗМАМИ В ЦІНОЗІ
ПЕРЕДМОВА
У перших двох лекціях по структурі біогеоценозу йшлося про видовий склад та просторову структуру фітоценозу як головного компонента біогеоценозу. У цій лекції розглядається функціональна структура біоценозу. В.В. Мазинг (1973) виділяються три напрями, розроблені ним для фітоценозів.
1. Структура як синонім складу(видова, конституційна). У цьому сенсі говорять про видову, популяційну, біоморфологічну (склад життєвих форм) та інші структури ценозу, маючи на увазі лише один бік ценозу – склад у широкому значенні.
2. Структура як синонім будови(Просторова, або морфоструктура). У будь-якому фітоценозі рослини характеризуються певною приуроченістю до екологічних ніш і займають певний простір. Це стосується й інших компонентів біогеоценозу.
3. Структура як синонім сукупностей зв'язків між елементами(функціональна). У основі розуміння структури у сенсі лежить вивчення взаємовідносин між видами, насамперед вивчення прямих зв'язків – біотичний коннекс. Це вивчення ланцюгів і циклів харчування, що забезпечують кругообіг речовин і розкривають механізм зв'язків трофічних (між тваринами та рослинами) або топічних (між рослинами).
Усі три аспекти структури біологічних систем тісно взаємопов'язані на ценотическом рівні: видовий склад, конфігурація та розміщення структурних елементів у просторі є умовою їхнього функціонування, тобто. життєдіяльності та продукування рослинної маси, а останнє, у свою чергу, значною мірою визначає морфологію ценозів. І всі зазначені аспекти відбиває умови середовища, у яких формується біогеоценоз.
Список літератури
Воронов А.Г. Геоботаніка. Навч. Посібник для ун-тів та пед. ін-тов. Вид. 2-ге. М: Вища. шк., 1973. 384 с.
Мазинг В.В. Що таке структура біогеоценозу // Проблеми біогеоценології. М: Наука, 1973. С. 148-156.
Основи лісової біогеоценології / за ред. Сукачова В.М. і Діліса Н.В.. М.: Наука, 1964. 574 с.
Запитання
1. Взаємозв'язки у біогеоценозі:
3. Типи відносин між організмами в цінозі:
а) Симбіоз
б) Антагонізм
1. Взаємозв'язки у біогеоценозі
Біоценотичний коннекс- Складний клубок взаємовідносин, «розмотування» якого може бути зроблено різними способами. Під шляхами розшифровки функціональної структури маються на увазі окремі підходи.
Біогеоценоз загалом є ту лабораторію, в якій відбувається процес акумуляції та трансформації енергії. Цей процес складається з багатьох різноманітних фізіологічних і хімічних процесів, що також взаємодіють між собою. Взаємодії між компонентами біогеоценозу виражаються в обміні речовиною та енергією між ними.
Взаємини між організмами та середовищем, які становлять одну з основ для розуміння істоти біогеоценозу, відносяться до екологічномунапрямку. Взаємини між особами одного виду ставляться зазвичай до популяційномурівнем, а взаємини між різними видами та різними біоморфами становлять основу вже біоценотичногопідходу.
а) Взаємодія між ґрунтом та рослинністю
Взаємодія між ґрунтом і рослинністю весь час відбувається у відомому сенсі «кругообіг» речовини та перекачування мінеральних речовин з різних горизонтів ґрунту в надземні частини рослин, а потім повернення їх у ґрунт у вигляді рослинного опаду. Таким чином здійснюється перерозподіл мінеральних речовин ґрунту по його горизонтах.
Особливо важливу роль цьому процесі грає опад, так звана лісова підстилка, тобто накопичується на поверхні власне ґрунту шар із залишків листя, гілок, кори, плодів та інших частин рослин. У лісовій підстилці відбувається руйнація та мінералізація цих рослинних решток.
Величезну роль грає рослинність також у водному режимі ґрунту, поглинаючи вологу з певних горизонтів ґрунту, віддаючи його потім в атмосферу шляхом транспірації, впливаючи на випаровування води з поверхні ґрунту, впливаючи на поверхневий стік води та підземне її переміщення. При цьому вплив рослинності на ґрунтові умови залежить від складу рослинності, її віку, висоти, потужності та густоти.
б) Взаємодії між рослинністю та атмосферою
Не менш складні взаємодії спостерігаються між рослинністю та атмосферою. Зростання та розвитку рослинності залежить від температури, вологості повітря, його руху і складу, а й навпаки – склад, висота, ярусність і густота рослинності впливають ці властивості атмосфери.
Тому кожен біогеоценоз має свій клімат. фітоклімат), тобто. ті властивості атмосфери, які викликані рослинністю.
в) Взаємозв'язок між мікроорганізмами та різними компонентами біогеоценозу
Одночасно мікроорганізми прямо чи опосередковано перебувають у взаємодії з тваринами (як хребетними, і безхребетними).
г) Взаємини між рослинами
Інші «впливи» рослин: ослаблення дії вітру, захист від вітровалу та бурелому; накопичення з відмираючих та опадаючих залишків рослин, листя, гілок, плодів, насіння тощо. лісової підстилки, яка не тільки опосередковано впливає на рослини через зміну ґрунтових процесів, а й створює особливі умови для проростання насіння та розвитку сходів тощо.
Вивчення биоморф як моделей найбільш істотних екологічних рис видів перспективно з'ясування загальних ценогеографических закономірностей.
д) Взаємозв'язок рослинності з тваринним світом
Не менш тісний взаємозв'язок рослинності з тваринним світом, що населяє даний біогеоценоз. Тварини в процесі своєї життєдіяльності багатосторонньо впливають на рослинність як безпосередньо, харчуючись нею, витоптуючи її, будуючи в ній або за допомогою її своє житло, притулку, сприяючи запиленню квіток і поширенню насіння або плодів, так і побічно, змінюючи грунт, удобрюючи , взагалі змінюючи її хімічні та фізичні властивості, а певною мірою впливаючи і на атмосферу.
Взаємини між різними трофічними рівнями відносяться до трофіко-енергетичного напрямку (Odum, 1963) і є об'єктом багатьох досліджень, що набув широкого розвитку в останні десятиліття. Це дає можливість виявити загальний характер та кількісні показники обміну речовин та енергії, розкриваючи тим самим біогеофізичну та біогеохімічну роль живого покриву.
е) Взаємодія між неживими (абіотичними) компонентами
Не тільки живі організми взаємодіють з іншими компонентами біогеоценозу, але й останні взаємодіють один з одним. Кліматичні умови (атмосфера) впливають на ґрунтоутворювальний процес, а ґрунтові процеси, визначаючи виділення вуглекислоти та інших газів (дихання ґрунту), змінюють атмосферу. Грунт впливає тваринний світ, як її населяющий, але опосередковано і весь інший тваринний світ. Тваринний світ впливає на грунт.
2. Фактори, що впливають на взаємодію компонентів біогеоценозу
Рельєф та біогеоценоз.Будь-який біогеоценоз, займаючи відоме місце у природі, пов'язані з тим чи іншим рельєфом. Але рельєф сам собою не входить до компонентів біогеоценозу. Рельєф є лише умовою, що впливає процес взаємодії названих вище компонентів, і, відповідно до цього, з їхньої властивості і структуру, визначаючи напрям і інтенсивність процесів взаємодії. У той самий час взаємодія компонентів біогеоценозу нерідко може призводити до зміни рельєфу і до створення особливих форм мікрорельєфу, а відомих випадках і мезо- і макрорельєфу.
Вплив людини на біогеоценоз.Людина не входить до компонентів біогеоценозів. Він, однак, є надзвичайно потужним фактором, що може не тільки тією чи іншою мірою змінювати, а й створювати шляхом культури нові біогеоценози. Нині майже немає лісових біогеоценозів, які не зазнали впливу господарської, а часто й безгосподарної діяльності людини.
Взаємовплив між біогеоценозами.Разом з тим кожен біогеоценоз, так чи інакше, впливає на інші біогеоценози і взагалі явища природи, сусідні з ним або тією чи іншою мірою віддалені від нього, тобто обмін речовиною та енергією має місце не тільки між компонентами даного біогеоценозу, але і між самими фітоценози. Найчастіше провідним чинником є конкурентні відносини між фітоценози. Фітоценоз сильніший витісняє фітоценоз менш стійкий, наприклад, за певних умов відбувається витіснення соснового фітоценозу ялиновим, а разом з цим змінюється і весь біогеоценоз.
Таким чином, взаємодія всіх компонентів біогеоценозу, особливо лісового біогеоценозу (включаючи воду в грунтах та атмосфері), дуже різноманітна і складна:
Рослинність постійно залежить від грунту, атмосфери, тваринного світу та мікроорганізмів.
Хімічний склад грунту, його волога та фізичні властивості впливають на зростання та розвиток рослин, на їх плодоношення та відновлюваність, на технічні властивості їх деревини деревних порід, на них, на ріст та розвиток всієї іншої рослинності.
Вся рослинність у свою чергу сильно впливає на грунт, визначаючи, головним чином, якість і кількість органічної речовини в грунті, впливаючи на фізичні та хімічні її особливості.
3. Типи відносин між організмами у цінозі
Організми можуть взаємодіяти один з одним постійно протягом усього життя або короткочасно. При цьому вони або стикаються один з одним, або впливають на інший організм на відстані.
Взаємні впливи рослин можуть мати те сприятливийдля їх зростання та розвитку характер, то несприятливий.У першому випадку умовно говорять про «взаємодопомогу», у другому – про «боротьбу за існування» між рослинами в широкому, дарвінському значенні, або про конкуренцію. Само собою зрозуміло, що всі ці взаємовпливи між організмами в біоценозі одночасно грають велику роль і в біогеоценозі в цілому. Вони можуть проходити між особинами як різних видів, так і того самого виду, тобто можуть бути як міжвидовими, так і внутрішньовидовими.
Відносини між організмами дуже різноманітні. Вдала класифікація цих відносин Г. Кларка (Clark, 1957) (табл. 1).
Таблиця 1
Класифікація відносин між організмами (за Кларком, 1957)
| Вид А | Вид Б | Відносини Умовні знаки: «+» – збільшення чи користь у життєвому процесі внаслідок відносин, «–» – зменшення чи збитки, 0 – відсутність помітного ефекту. - Відносини між організмами, зазвичай різних видів і перебувають у більш менш тривалому контакті, при яких один або обидва організму витягують з цих відносин користь і жоден не зазнає шкоди. Перший тип симбіотичних відносин, коли користь отримують обидва організми, називається мутуалізмом, другий, коли користь отримує лише один з організмів, - комменсалізм («нахлібництвом»). Мутуалізм Симбіоз азотфіксуючих організмів з голонасінними та квітковими рослинами – відносини між вищою рослиною та бактеріями. На коренях багатьох рослин зустрічаються бульби, утворені бактеріями чи рідше грибами. Бульбякові бактерії фіксують атмосферний азот і переводять його в доступну для вищих рослин форму.
|
|||||||


У кожному фітоценозі підбираються рослини:
Які представляють різні життєві форми і котрі посідають у різних синузіях, ярусах, микроценозах, тобто. утворюють групи, що характеризуються неоднаковим ставленням до середовища та неоднаковим місцем у фітоценозі;
Диференційовані за термінами сезонних фаз.
Поєднання в одному фітоценозі рослин з різними екологічними особливостями – тіньолюбних і світлолюбних, різною мірою пристосованих до нестачі вологи та інших факторів середовища, дозволяє фітоценоз найбільш повно використовувати умови місцеперебування.
Зміна видів відбувається не відразу, поступово один вид витісняє інший, тому між фітоценоз зазвичай немає чіткої межі. Смуга, на якій відбувається зміна фітоценозів, називається екотоном. В екотоні, як правило, присутні види суміжних угруповань, і мозаїчність рослинного покриву тут вища, але життєвий стан видів-домінантів обох угруповань в екотоні зазвичай гірший, ніж у тих ценозах, умови яких цим видам відповідають більше.
Витіснення одних видів іншими на межі фітоценозів (щоправда, не одновидових) відбувається і без змін умов середовища, внаслідок різної конкурентної спроможності видів, зокрема різної енергії вегетативного розмноження.
ПРИКЛАДИ. Так, відомий усім бур'янистий злак пирій не тільки здатний заглушити культурні посіви, а й витісняє багато диких видів (кропиву, чистотіл та ін.), що ростуть з ним по сусідству і вегетативно розмножуються дуже слабо. Навіть конюшина повзуча поступово поступається позиції пирію.
Дуже сильною конкурентною здатністю має мох сфагнум. У міру розростання він у буквальному значенні поглинає сусідні рослини. У районах поширення вічної мерзлоти фітоценози з пануванням сфагнуму займають великі простори, витісняючи зони свого впливу як трави і чагарники, а й чагарники і дерева.
Внаслідок боротьби за існування відбувається диференціація видів, що утворюють фітоценоз. У той самий час будова фітоценозу – як результат боротьби за існування, а й результат пристосування рослин до зменшення напруженості цієї боротьби. У фітоценозі види підібрані таким чином, що доповнюють один одного своїми властивостями.
Лекція 10. СПОРУЖЕНІСТЬ ВИДІВ У ФІТОЦЕНОЗІ. ВСЕРЕДИНІ- І МІЖВИДОВІ ВІДНОСИНИ В БІОГЕОЦЕНОЗІ.
Запитання
а) Диференціація цінопуляцій
в) Перенаселення виду
4. Сполученість видів у фітоценозі
Одним із якісних показників видів, що входять до складу фітоценозу, є їх сполученість (асоційованість). Сполученість відзначається лише за присутністю чи відсутністю двох видів на пробному майданчику. Існує позитивна чи негативна сполученість.
Позитивна буває тоді, коли вид зустрічається з видом А частіше, ніж це мало б місце при незалежному один від одного розподілі обох видів.
Негативна сполученість спостерігається тоді, коли вид зустрічається разом з видом А рідше, ніж це мало б місце при незалежному один від одного розподілі обох видів.
У підручнику геоботаніки А.Г. Воронова наводиться формули та таблиці сполученості В.І. Василевича (1969), за допомогою яких можна опрацювати дані щодо присутності та відсутності двох видів та визначити рівень їх сполученості та наводиться приклад розрахунку.
Для визначення ступеня сполученостідвох і більше видів існують різні коефіцієнти (Грейг-Сміт, 1967; Василевич, 1969).
Один із них запропонований Н.Я. Кацем (Кац, 1943) і розраховується за такою формулою:

Якщо К >1, це означає, що це вид зустрічається частіше з іншим видом, ніж нього (сполученість позитивна); якщо До<1, то это значит, что данный вид чаще встречается без другого вида, чем с ним (сопряженность отрицательная). Если К = 1, то виды индифферентно относятся друг к другу, и встречаемость данного вида вместе с другим не отличается от общей встречаемости первого вида в фитоценозе.
Природно, сполученість тим вище, що більше коефіцієнт сполученості віддаляється від одиниці.
Найчастіше визначення сполучення використовують квадратні майданчики по 1 м 2 , іноді прямокутні по 10 м 2 . Б.А. Биков запропонував круглі майданчики по 5 дм2 (радіус 13 см). Але якщо величина пробної площадки можна порівняти з величиною особини хоча б одного виду, то хибне уявлення про негативну кореляцію з іншим видом буде отримано тільки тому, що дві особи не можуть займати одне й те саме місце. І тут слід збільшити розміри майданчиків.
Слід їх збільшити і в тому випадку, якщо в фітоценозі, наприклад, 3 види, і особи одного виду великі, а двох інші - дрібні. На обліковому майданчику, зайнятому "великим" виглядом, можуть бути витіснені ним "дрібні". Це створює враження, що між видами з дрібними особами існує позитивна кореляція, чого насправді немає. Це уявлення за достатніх розмірів пробних майданчиків зникне.
У випадках, коли ставляться за мету встановити наявність або відсутність сполученості, можна закладати майданчики "в строго систематичному порядку", наприклад, впритул один до одного. Якщо ж ступінь сполученості встановлюється по одній з формул, необхідний випадковий відбір зразків.
Про що свідчить поєднання?
Якщо йдеться про позитивноюспряженості, то вона може мати місце у двох випадках:
Види «пристосовуються» один до одного настільки, що частіше зустрічаються один з одним (світи видів тих чи інших типів лісу, у с/г часник і морква), ніж порізно
Обидва види подібні за своїми екологічними особливостями і частіше живуть разом тому, що в межах одного фітоценозу умови виявляються більш сприятливими для обох видів (види тих самих ярусів).
При негативноюспряженості вона може залежати від того, що в результаті міжвидової боротьби:
Обидва види стали антагоністами (не треба поряд саджати суницю та моркву; волжанка, вейник – пригнічують своїх сусідів по еконіші);
У видів різне ставлення до вологи, висвітлення та інших факторів середовища в межах фітоценозу (рослини різних ярусів та різні парцел).
5. Внутрішньо- та міжвидові відносини в біогеоценозі
а) Диференціація цінопуляцій
Лісівникам давно відомо, що кількість стовбурів дерев на одиницю площі з віком насаджень зменшується. Чим більш світлолюбна порода і чим кращі умови проростання, тим швидше самовиріджується деревостою. Загибель дерев особливо інтенсивна у перші десятиліття і поступово зменшується із збільшенням віку лісу. Це показано у таблиці 2.
Таблиця 2
Зменшення загальної кількості стволів з віком (за Г. Ф. Морозовим, 1930)
| Вік у роках | Число стволів на 1 га | ||
| Буковий ліс на раковистому вапняку | Буковий ліс на ґрунті з строкатого пісковику | сосновий ліс на супіщаному грунті |
|
| 10 | 1 048 660 | 860 000 | 11 750 |
| 20 | 149 800 | 168 666 | 11 750 |
| 30 | 29 760 | 47 225 | 10 770 |
| 40 | 11 980 | 14 708 | 3 525 |
| 50 | 4 460 | 8 580 | 1 566 |
| 60 | 2 630 | 4 272 | 940 |
| 70 | 1 488 | 2 471 | 728 |
| 80 | 1 018 | 1 735 | 587 |
| 90 | 803 | 1 398 | 509 |
| 100 | 672 | 1 057 | 461 |
| 110 | 575 | 901 | 423 |
| 120 | 509 | 748 | 383 |
| 130 | – | 658 | 352 |
| 140 | – | 575 | 325 |
| 145-150 | – | 505 | 293 |
 | Кількість відмерлих дерев бука за 100 років (від 10 до 110 років) становила понад 1 млн. на багатих ґрунтах і понад 850 000 на бідних, а для сосни – понад 11000, що пов'язано з малою кількістю стовбурів цієї породи вже у десятирічному віці. Сосна дуже світло-любна, тому вже до 10 років у неї відбувся значний відпад. В результаті за сто років зберігається одне дерево бука з 1800 на багатих ґрунтах і з 950 – на бідніших, і одне дерево сосни із 28. На рис. 5 теж показано, що загибель більш світлолюбної породи (сосна) відбувається швидше, ніж тіньовитривалих (бук, ялина, ялиця). Таким чином, відмінності в темпах зріджування у деревостою пояснюються: 1) різним світлолюбством (тіньовитривалістю); 2) збільшенням темпу зростання хороших умовах і, як наслідок, швидким зростанням його потреб у екологічних ресурсах, чому конкуренція між видами стає дедалі більше напруженою. |
 | Конкуренція всередині виду, значно інтенсивніша, ніж між особинами різних видів, проте, у разі відбувається диференціація особин за висотою. У лісі дерева одного виду можна розподілити за класами Крафт (рис. 6). Перший клас поєднує дерева добре розвинені, що піднімаються вище за інших – виключно держпідтримуючі, другий клас – панівні, третій – панівні, з розвиненими, дещо здавленими з боків, четвертий – заглушені дерева, п'ятий – дерева пригнічені, відмирають або відмерлі. Подібна картина зменшення кількості екземплярів рослин (цього разу протягом одного сезону) та диференціації за висотою спостерігається і у фітоценозах, утворених однорічними рослинами, наприклад солеросом трав'янистим (Salicornia herbacea). |
б) Екологічний та фітоценотичний оптимуми
Кожен вид має свою оптимальну щільність. Оптимальною щільністю називають ті межі густини, які забезпечують найкраще відтворення виду та його найбільшу стійкість.
ПРИКЛАДИ. Для дерев в рединах оптимальна щільність дуже невелика, вони ростуть поодинці на значній відстані один від одного, зате для лісоутворюючих порід вона набагато вища, а для болотяних сфагнових мохів (Sphagnum) надзвичайно висока.
Величина оптимальної площі та реакція на загущення залежить від умов, у яких проходила еволюція виду: одні види розвивалися за умов високої щільності популяції, інші – за умов низької щільності; в одних випадках щільність була постійною, в інших – мінливою. Види, еволюціонували за умов постійної щільності, різко реагують підвищення щільності межі оптимальної уповільненням зростання; види, які розвинулися за умов безперервно змінюваної щільності, реагують зміну щільності межі оптимуму слабо.
Кожен вид має два оптимуми розвитку: екологічний, що позначається на розмірах особин виду, і фітоценотичний, що характеризується найбільш високою роллю даного виду у фітоценозі, що виражається в його різноманітті та ступеня проектного покриття. Ці оптимуми та ареали можуть не збігатися. У природі частіше зустрічається фітоценотичний оптимум, а екологічний можна виявити штучно створюючи рослинам різні умови.
ПРИКЛАДИ. Багато галофітів краще розвиваються не на засолених ґрунтах, де вони утворюють співтовариства, а на вологих ґрунтах із незначним вмістом солей. Багато рослин скель із ксероморфним виглядом свій екологічний оптимум мають на луках.
Розбіжність екологічного та фітоценотичного оптимумів – результат боротьби за існування між рослинами. У ряді випадків у процесі боротьби за існування рослини відтісняються в екстремальні умови з найбільш сприятливих для них фітоценозів.
ПРИКЛАДИ. Ялиця білокора і ялина аянська не тому ростуть у вищих гірських поясах, що там кращі умови, а тому, що туди їх витісняють ялина корейська, кедр і ялиця цільнолистяна. Так само і світлолюбні осика і береза поступаються своїми більш сприятливими екотопами темнохвойним породам. Так само травами з заплавних жител витісняються мохи і чагарники.
в) Перенаселення виду
Для характеристики густини виду існує таке поняття, як перенаселення. Розглядають кілька видів перенаселення: абсолютне, відносне, вікове, умовне та локальне.
Під абсолютним перенаселеннямрозуміють такі умови загущення, у яких неминуче настає масова загибель, що має загальний характер. (Понад густий посів - насіння зашпаровується суцільним шаром або в два-три шари), за якого, за умови дуже дружних одночасних сходів на великих ділянках, гинуть всі рослини, крім крайніх).
Під відносним перенаселеннямрозуміють такі умови загущення, за яких загибель рослин більш-менш підвищена, ніж при густоті оптимальної для виду. При цьому загибель рослин вибіркова, Дія відбору м'якша, ніж у разі абсолютного перенаселення.
Під віковим перенаселенням розуміють перенаселення, що виникає з віком внаслідок нерівномірного росту кореневих систем (наприклад, коренеплодів) або надземних частин рослин (у дерев).
Умовно перенаселеними називають сильно загущені фітоценози, в яких гострота взаємин між рослинами знижена тимчасовою затримкою їх зростання настільки, що зріджування іноді повністю припиняється. Так, багато рослин дуже довго залишаються в ювенільному (юнацькому) стані, зберігаючи дуже високу виживаність. Варто змусити рослини до активного зростання, як настає справжнє перенаселення. Наприклад, сильно пригнічені особини деревних порід під пологом густого лісу мають вигляд підросту.
Локальним перенаселеннямназивають випадки перенаселення в гніздових насадженнях дуже високої щільності та малої площі, в яких завдяки малій площі гнізда виживання кожної окремої особини визначається не становищем цієї особини в гнізді, а її особливостями, іншими словами, загибель тут має вибірковий характер.
Яке значення явищ перенаселення для боротьби за існування і, отже, для процесу еволюції?
Перенаселення може мати місце в одних випадках та в одні періоди життя рослин і відсутнє в інших випадках та інші періоди життя рослин. Залежно від ступеня перенаселення і особливостей організмів воно здатне як прискорювати, і уповільнювати процес еволюції. При невеликих ступенях перенаселення воно викликає диференціацію особин і тим самим прискорює процес еволюції; при значних ступенях може викликати збіднення популяції, зменшення плодючості й у результаті – уповільнення еволюційного процесу. Перенаселення уповільнює і прискорює процес природного відбору, але не є йому перешкодою і є неодмінною умовою відбору, оскільки відбір може і без перенаселення.
Ми знаємо, що для двох найбільших груп органічного світу - тварин і рослин - значення перенаселення неоднаково: воно відіграє набагато більшу роль у рослинному світі, так як рухливість тварин дозволяє їм у ряді випадків уникнути перенаселення.
Для різних систематичних та екологічних груп рослин перенаселення грає не одну й ту саму роль. Розвиток більшої, ніж може згодом вижити, кількості проростків та молодих рослин забезпечує виду панування у фітоценозі. Якби проростки виду, що переважає у фітоценозі, були одиничними, то в масах розвинулися б проростки іншого виду, і цей інший вид міг би стати домінуючим у фітоценозі. Панівний вигляд зазвичай дає велику кількість проростків, але цілком природно, що лише невелика частина досягає зрілості. Отже, загибель великої кількості молодих рослин у цьому випадку неминуча, саме вона забезпечує процвітання виду та збереження його становища у фітоценозі. Крім молодих рослин гине велика кількість діаспор - зародків рослин (насіння, плоди, суперечки) - ще до початку їх розвитку (з'їдаються тваринами, гинуть в несприятливих умовах і т. д.). Отже, величезна кількість діаспор, утворених рослинами, забезпечує як панування, але найчастіше і саме існування виду.
Внутрішньовидова конкуренція завжди більш жорстока, ніж міжвидова, оскільки особини одного виду більш подібні один з одним і висувають більш подібні вимоги до середовища, ніж особи різних видів. Однак у природі, мабуть, дедалі складніше. Так, при вихованні двох видів у чистих посівах і змішаних (причому сумарне число особин на одиницю площі в змішаному посіві дорівнює числу особин на одиницю площі в чистих посівах обох видів) спостерігаються три типи взаємовідносин (Сукачов, 1953).
1. При спільному посіві обидва види розвиваються краще, ніж кожен із них у одновидовому посіві. У цьому випадку міжвидова боротьба виявляється слабшою за внутрішньовидову, що відповідає точці зору Ч. Дарвіна.
2. З двох видів один краще почувається у суміші, ніж у чистому посіві, а другий – гірше у суміші і краще в чистому посіві. У цьому випадку для одного з видів міжвидова боротьба виявляється суворішою, ніж внутрішньовидова, а для іншого – навпаки. Причини цього різні: виділення одним із видів колінів, шкідливих для особин іншого виду, відмінність екологічних особливостей видів, вплив продуктів розкладання відмерлих залишків одного виду на інший, відмінності у будові кореневої системи та інших особливостей.
3. Обидва види почуваються у суміші гірше, ніж у одновидових посівах. І тут обох видів внутрішньовидова боротьба менш сувора, ніж межвидовая. Цей випадок дуже рідкісний.
Слід пам'ятати, що відносини між парою будь-яких видів залежить від умов досвіду: складу живильного середовища, вихідної чисельності рослин, умов освітлення, температурного режиму та інших причин.
Всупереч наведеному нами епіграфу, ми знаємо, що в жодному куточку земної кулі немає поодинці живих рослин, тварин, мікроорганізмів. У рослин багато друзів, але багато ворогів крім грибків і бактерій. Дуже важливу роль, іноді корисну, а іноді й украй шкідливу, грають у житті рослин комахи. У ході розвитку живого світу протягом мільйонів років склалися різноманітні, часом дуже складні, взаємини між рослинами і комахами. Достатньо згадати роль комах у так званому перехресному запиленні рослин, важливість рослин для життя бджіл; Згадаймо і комах - шкідників лісів, городів, садів.
У лісі, на лузі, у болоті, морях - всюди в природі життя рослин і комах взаємопов'язане і представляє в деяких відносинах ніби одне ціле. Наука, накопичуючи все більше і більше фактів про рослини та тварини, в той же час вивчає закономірності у житті рослинних та тваринних угруповань.
В озерах, річках, хвойних лісах, дубових гаях, чагарниках черемхи, на плантаціях цитрусових - скрізь складаються свої, своєрідні відносини між рослинами і тваринами, переважає своє тваринне населення, приурочене лише до тих чи інших видів рослин, певного характеру грунту і т.д. . Самка падальних мух відкладає яйця в рослини, що гниють, і тваринні продукти. Зародки мух можуть розвиватися серед мікробів, що кишать. Ікринка, що вийшла з організму риби, може опинитися в сусідстві з різними мікробами, рослинами і тваринами.
У кожному типі лісу мешкають свої тваринні організми. У букових лісах зустрічається від 3 до 4 тисяч видів рослин і від 6 до 7 тисяч видів тварин (мікроскопічні одноклітинні тварини тут не приймаються). Виявляється, що значна частина тварин суворо присвячена саме буковим лісам. Близько 1800 видів тварин та 1170 видів рослин знаходять сприятливі умови для життєдіяльності лише у букових лісах.
Наведемо приклад. Він неприємний, але, мабуть, корисний, тому що змусить ретельніше мити фрукти та овочі перед тим, як їх їсти. У чудовому Петергофському парку під Ленінградом вчені підрахували, скільки різних комах та кліщів може бути, наприклад, на ягоді. На 400 грамах суниці виявилося близько 600 екземплярів головним чином кліщів, на 400 грамах чорниці - близько 1100, на такій же кількості малини - 5000, горобини - понад 7000. А в кроні однієї великої берези їх близько 5-10 мільйонів.
А ось ще не менш своєрідний випадок взаємовідносин організмів у природі на основі виділення фітонцидів. Вчені довго не могли зрозуміти, яким чином всмоктана п'явкою кров стає для неї їжею. Чужа кров з її складними хімічними речовинами повинна спочатку змінитися, а потім вже у простішому вигляді вона може бути засвоєна клітинами п'явки. У тварин і людини в кишечнику виробляються особливі речовини - ферменти, завдяки яким відбувається травлення. У кишечнику п'явок цих речовин немає. Що ж виявилось? У кишечнику п'явок постійно живе сильно розмножуючись бактерія псевдомонас гірудініс. Ця бактерія є благодійною для п'явки. Вона допомагає перетравлювати всмоктану кров, виділяючи відповідні речовини, і вона ж, виділяючи свої фітонциди, вбивчі для інших мікробів, виявляється єдиною повновладною господинею в кишечнику п'явок і не допускає жодного іншого бактеріального забруднення. Ось чому кишечник п'явки абсолютно чистий, від п'явки, що смокче кров, ніколи не захворієш на заразну хворобу. Недарма наукова медицина користується п'явками для лікування багатьох хвороб.
Людина, будучи частиною природи, стала її творцем, найважливішим чинником її еволюції. Здійснюючи гігантське будівництво, соціалістична держава повинна передбачати і біологічні наслідки: які рослинні угруповання складуться при посадці тих чи інших деревних порід, як зміниться тваринний і рослинний світ при будівництві нових каналів, як зміниться життя водойм? Біологи всіх спеціальностей, беручи участь у цих величних справах, зайняті і вирішенням проблем, що виникають.
Все нові відкриття у взаєминах тварин і рослин використовуються в інтересах людини, ставляться на службу лісової промисловості, (медицини, сільського господарства, садівництва, городництва. Хочеться думати, що найближчими роками вдасться дещо витягти і з відкриття фітонцидів, корисне не тільки для боротьби з бактеріями, найпростішими та грибками, але й для регулювання життя рослинних угруповань та вищих тварин, а також активно застосувати фітонциди для збереження здоров'я людини.
Але повернемося до взаємин фітонцидів та комах.
Мимоволі напрошується думка, чи не відіграють і фітонциди будь-яку роль у приуроченості певних видів комах до тих чи інших рослин та рослинних угруповань? Чи не мають у природі леткі фітонциди будь-яке значення як відлякуючі або, навпаки, залучають комахи речовин? Чи не можна використовувати фітонциди в побуті та медицині як інсектициди - речовини, що вбивають шкідливих комах? Чи не можна науково пояснити народні рослинні засоби боротьби зі шкідливими комахами? Ця сфера досліджень настільки приваблива тому, що має величезне практичне значення.
Повідомимо деякі факти. Можливо, вони пробудять у читачів інтерес до спостережень та експериментів у природі.
Зробимо невелику екскурсію в минуле та повідомимо про одне відкриття, яке довелося зробити ще у 1928-1930 роках. Це відкриття згодом переконало нас у корисності вивчення впливу фітонцидів на багатоклітинних тварин, зокрема, на комах.
Вже в перші дні відкриття фітонцидів, коли було ясно, що леткі речовини деяких рослин діють згубно на грибки, постало питання: чи маємо ми справу з отрутами, шкідливими протоплазмі певних клітин, або з отрутами для будь-якої протоплазми? Тепер ми добре знаємо, що фітонциди діють вибірково: вбивають одні клітини та організми і не умертвляють, а навіть стимулюють інші.
Одними з перших дослідів із фітонцидів були досліди з яйцями молюсків - з тими клітинами, від яких починається розвиток цих організмів. Молюсків, «слимаків», дуже багато в морях, прісних водоймах і на суші (рис.21).
Водні молюски відкладають яйця на листі та стеблах рослин, на камінні та інших твердих предметах. Щоразу їх відкладається кілька десятків. Всі вони знаходяться в загальній прозорій драглистій масі, що грає важливу роль у запобіганні зародків від зовнішніх несприятливих впливів. Кожне яйце, своєю чергою, одягнене оболонками. Ці оболонки настільки прозорі, що крізь них за допомогою лупи легко спостерігати всі послідовні етапи розвитку зародка аж до формування мікроскопічного молюска, у якого вже виразно видно раковину. Звільнившись від оболонок, молюс починає вести самостійне існування як доросла тварина.
Мікроскопічні зародки молюсків беззахисні на вигляд. Але це враження помилкове. Оболонки яєць мають таку будову і склад, що дуже багато речовин, отруйні і для складніше організованих тварин, для яєць молюсків абсолютно нешкідливі. Роздавити яйце молюска, звичайно, легко, можна вбити яйце високою температурою, але підібрати для цих ніжних, витончених, прозорих клітин хімічні отрути вченому непросто, оскільки багато отруйних для протоплазми речовини не проникають крізь оболонку яйця.
Візьмемо одну яйцекладку будь-якого молюска на такій стадії розвитку, коли крізь прозорі оболонки видно рух зародків. Розріжемо цю яйцекладку на дві половини. Одну половину використовуємо для досвіду, а інша залишиться контрольною.
Досвідчену половину яйцекладки помістимо в краплі води на скло, а поруч покладемо щойно приготовлену на терці цибулинну кашку. У перші ж секунди (зазвичай пізніше як за 30 секунд) ми помітимо різке прискорення руху зародків: вони у збуджений стан. За хвилину-другу цей стан змінюється повною зупинкою руху. Пройде деякий час, і ми побачимо при збереженні оболонок яєць повний розпад зародків. Яйця контрольної половини яйцекладки, що також перебувають у воді, чудово розвиваються.
Дуже багато рослин мають такі властивості, наприклад листя, нирки, кора черемхи (рис.22), кореневища хрону, листя лавровишні, клена, дуба, голки ялиці і т.д. Особливий біологічний інтерес для розуміння взаємин у природі рослин та тварин представляє дію фітонцидів водних рослин на яйця молюсків, жаб, риб та інших організмів. Початкові дослідження вже дали несподівані результати. Одні водні та прибережно-водні рослини (деякі синьозелені водорості, спірогіра, манник) гальмують розвиток зародків молюсків, інші його стимулюють.
І знову виникає думка: чи невипадково це явище? Чи мають відношення виявлені факти до захисних властивостей водних рослин? Чи байдуже для рослин, якщо на них відкладають яйця молюски та інші водяні тварини? Чи байдуже для молюсків, на яких рослинах відкласти яйця? Таким чином ми підходимо до питання про біологічне самоочищення водойм, до питання, чи не відіграють фітонциди водних рослин певну роль і в регулюванні складу тваринного, рослинного та мікробного населення водойм. На цих питаннях ми зупинимося надалі.