Strøm av energi i naturen. Syklus av materie og energi i naturen
Ethvert liv krever en konstant strøm av energi og materie. Energi brukes på å utføre grunnleggende livsreaksjoner, materie brukes til å bygge organismer. Eksistensen av naturlige økosystemer er ledsaget av komplekse prosesser for material- og energiutveksling mellom levende og livløs natur. Disse prosessene er svært viktige og avhenger ikke bare av sammensetningen av biotiske samfunn, men også av deres fysiske miljø.
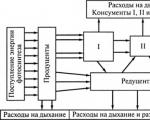
Strøm av energi i samfunnet– dette er dens overgang fra organismer på ett nivå til et annet i form av kjemiske bindinger av organiske forbindelser (mat).
Strømning (syklus) av materie er bevegelse av materie i form av kjemiske elementer og deres forbindelser fra produsenter til nedbrytere og deretter (gjennom kjemiske reaksjoner som skjer uten deltakelse av levende organismer) igjen til produsenter.
Sirkulasjonen av materie og strøm av energi er ikke identiske konsepter, selv om ulike energiekvivalenter (kalorier, kilokalorier, joule) ofte brukes til å måle materiens bevegelse. Dette forklares delvis av det faktum at på alle trofiske nivåer, med unntak av det første, overføres energien som er nødvendig for organismers liv i form av stoffet til konsumert mat. Bare planter (produsenter) kan direkte bruke solens strålende energi til sin livsaktivitet.
En streng måling av et stoff som sirkulerer i et økosystem kan oppnås ved å ta hensyn til sirkulasjonen av individuelle kjemiske elementer, først og fremst de som er hovedbyggematerialet for cytoplasmaet til plante- og dyreceller.
I motsetning til stoffer som kontinuerlig sirkulerer gjennom forskjellige blokker i økosystemet og alltid kan gå inn i syklusen igjen, kan energi brukes i kroppen bare én gang.
I følge fysikkens lover kan energi endres fra en form (som lysenergi) til en annen (som matens potensielle energi), men den blir aldri skapt igjen eller ødelagt. Det kan ikke være en enkelt prosess forbundet med transformasjonen av energi uten å miste noe av den. I transformasjonene forsvinner en viss mengde energi i form av varme og går derfor tapt. Av denne grunn kan det ikke være transformasjoner, for eksempel av matstoffer til stoffene som utgjør kroppen til organismen, som skjer med hundre prosent effektivitet.
Eksistensen av alle økosystemer avhenger av en konstant strøm av energi, som er nødvendig for at alle organismer skal opprettholde sine vitale funksjoner og selvreproduksjon.
Bare omtrent halvparten av solfluksen som faller på grønne planter absorberes av fotosyntetiske elementer, og bare en liten brøkdel av den absorberte energien (fra 1/100 til 1/20 av delen) lagres i form av energi som er nødvendig for aktiviteten av plantevev.
Når man beveger seg bort fra primærprodusenten, avtar hastigheten på energiflyten (det vil si mengden energi, uttrykt i energienheter, overført fra et trofisk nivå til et annet) kraftig.
Fallet i mengden energi under overgangen fra ett trofisk nivå til et høyere bestemmer selv antallet av disse nivåene. Det anslås at ethvert trofisk nivå bare mottar omtrent 10 % (eller litt mer) av energien til det forrige nivået. Derfor overstiger det totale antallet trofiske nivåer sjelden 3–4.
Forholdet mellom levende stoffer på forskjellige trofiske nivåer følger generelt den samme regelen som forholdet mellom innkommende energi: jo høyere nivå, desto lavere er den totale biomassen og antallet av dens bestanddeler.
Forholdet mellom antall forskjellige grupper av organismer gir en idé om stabiliteten til samfunnet, fordi biomassen og antallet av enkelte populasjoner samtidig er en indikator på leverommet for organismer av denne og andre arter . For eksempel bestemmer antall trær i en skog ikke bare den totale tilførselen av biomasse og energi som finnes i dem, men også mikroklimaet, samt antall tilfluktsrom for mange insekter og fugler.
Pyramider av tall kan snus. Dette skjer når reproduksjonshastigheten til byttedyrpopulasjonen er høy, og selv ved lav biomasse kan en slik populasjon være en tilstrekkelig matkilde for rovdyr som har høyere biomasse, men lav reproduksjonsrate. For eksempel kan mange insekter leve og spise på ett tre (en omvendt populasjonspyramide). Den omvendte pyramiden av biomasse er karakteristisk for akvatiske økosystemer, der primærprodusentene (fytoplanktoniske alger) veldig raskt deler seg og formerer seg i antall, og deres forbrukere (zooplanktoniske krepsdyr) er mye større, men har en lang reproduksjonssyklus.
Beitemark og detritalkjeder
Energi kan strømme gjennom et fellesskap på mange forskjellige måter. Den representerer næringskjeden til alle forbrukere(forbrukersystem) med tillegg av ytterligere to lenker: dette dødt organisk materiale Og nedbryter næringskjeden(reduserende system).
Strøm av energi fra planter gjennom planteetere(de kalles beite), kalt næringskjeden for gressletter.
Restene av organismene de konsumerer som ikke brukes av forbrukerne fyller på dødt organisk materiale. Den består av avføring som inneholder en del av ufordøyd mat, samt dyrelik, vegetasjonsrester (blader, greiner, alger) og kalles detritus.
Strømmen av energi som stammer fra dødt organisk materiale og passerer gjennom nedbrytningssystemet, kalles den skadelige næringskjeden.
Sammen med likhetene er det store forskjeller i funksjonen til beite- og detritus-næringskjedene. Den består i at i i forbrukersystemet går avføring og døde organismer tapt, og i det reduktive systemet– Nei.
Før eller senere vil energien som finnes i dødt organisk materiale bli fullstendig brukt av nedbrytere og forsvinne som varme gjennom respirasjon, selv om dette krever at den passerer gjennom et system av nedbrytere flere ganger. De eneste unntakene er de tilfellene hvor lokale abiotiske forhold er svært ugunstige for nedbrytningsprosessen (høy luftfuktighet, permafrost). I disse tilfellene akkumuleres forekomster av ufullstendig bearbeidede, svært energikrevende stoffer, som over tid og under passende forhold forvandles til brennbare organiske fossiler - olje, kull, torv.
Syklus av stoffer i et økosystem
Integriteten til naturlige økosystemer kommer spesielt tydelig til uttrykk når man vurderer strømmen av materie som sirkulerer i dem. Stoffet kan overføres i lukkede sykluser (sirkulasjoner), gjentatte ganger sirkulerer mellom organismer og miljøet.
Sirkulære bevegelser (gjennom land, luft, vann) av kjemiske elementer (det vil si stoffer) kalles biogeokjemiske sykluser eller sirkulasjoner.
Grunnstoffene og oppløste salter som er nødvendige for liv kalles konvensjonelt næringsstoffer(gi liv) eller næringsstoffer. Blant biogene elementer skilles to grupper: makrotrofiske stoffer og mikrotrofe stoffer.
Makrotrofe stoffer dekke elementene som utgjør det kjemiske grunnlaget for vevet til levende organismer. Disse inkluderer: karbon, hydrogen, oksygen, nitrogen, fosfor, kalium, kalsium, magnesium, svovel.
Mikrotrofe stoffer inkluderer grunnstoffer og deres forbindelser, også svært viktige for eksistensen av levende systemer, men i ekstremt små mengder. Slike stoffer kalles ofte mikroelementer. Disse er jern, mangan, kobber, sink, bor, natrium, molybden, klor, vanadium og kobolt. Selv om mikrotrofiske elementer er essensielle for organismer i svært små mengder, kan mangelen på dem sterkt begrense produktiviteten.
Sirkulasjonen av næringsstoffer er vanligvis ledsaget av deres kjemiske transformasjoner. Nitratnitrogen kan for eksempel omdannes til proteinnitrogen, deretter omdannes til urea, omdannes til ammoniakk og igjen syntetiseres til nitratform under påvirkning av mikroorganismer. Ulike mekanismer, både biologiske og kjemiske, er involvert i prosessene med denitrifikasjon og nitrogenfiksering.
Reservene av næringsstoffer er variable. Prosessen med å sekvestre noen av dem som levende biomasse reduserer mengden som er igjen i det abiotiske miljøet. Og hvis planter og andre organismer til slutt ikke brytes ned, ville tilførselen av næringsstoffer være oppbrukt og livet på jorden ville opphøre. Av dette kan vi konkludere det aktiviteten til heterotrofer, først og fremst organismer som fungerer i detritale kjeder,– en avgjørende faktor for å opprettholde syklusen av næringsstoffer og dannelsen av produkter.
La oss se på noen numeriske data som indikerer omfanget av stoffoverføring, og snu til den biogeokjemiske karbonsyklusen. Den naturlige kilden til karbon som brukes av planter for syntese av organisk materiale er karbondioksid, som er en del av atmosfæren eller er oppløst i vann. Under fotosyntesen omdannes karbondioksid (karbondioksid) til organisk materiale som tjener som mat for dyr. Respirasjon, gjæring og forbrenning av drivstoff returnerer karbondioksid til atmosfæren.
Karbonreserver i atmosfæren på planeten vår er estimert til 700 milliarder tonn, i hydrosfæren - til 50 000 milliarder tonn I følge årlige beregninger, som et resultat av fotosyntese, er økningen i plantemasse på land og i vann 30 milliarder tonn og 150 milliarder tonn, henholdsvis Karbonkretsløpet fortsetter i ca 300 år.
Et annet eksempel er fosforsyklusen. Hovedreservene av fosfor inneholder forskjellige bergarter, som gradvis (som et resultat av ødeleggelse og erosjon) frigjør fosfatene sine til terrestriske økosystemer. Fosfater konsumeres av planter og brukes av dem til å syntetisere organiske stoffer. Når dyrekadaver brytes ned av mikroorganismer, føres fosfater tilbake til jorden og brukes deretter igjen av planter. I tillegg føres en del av fosfatene ut av vassdrag ut i havet. Dette sikrer utviklingen av planteplankton og alle næringskjeder som er avhengige av det. Noe av fosforet i sjøvann kan gå tilbake til land i form av guano.
økosystemets biosfæres energisyklus
Introduksjon
Konsept og struktur av økosystemer
1 Begrepet økosystemer
2 Klassifisering av økosystemer
3 Sonering av makroøkosystemer
4 Økosystemstruktur
Faktorer som sikrer integriteten til økosystemene
1 Stoffsykluser
2 Energiflyt i økosystemer
3 Dynamiske prosesser som sikrer integritet og bærekraft i økosystemer
4 Biosfæren som et globalt økosystem som sikrer integriteten og bærekraften til økosystemene
Konklusjon
Introduksjon
Et økosystem er enhver samling av organismer og uorganiske komponenter der sirkulasjon av stoffer kan forekomme. Ifølge N.F. Reimers (1990), et økosystem er ethvert fellesskap av levende vesener og dets habitat, forent til en enkelt funksjonell helhet, som oppstår på grunnlag av gjensidig avhengighet og årsak-og-virkning-forhold som eksisterer mellom individuelle miljøkomponenter. A. Tansley (1935) foreslo følgende forhold:
Det er mikroøkosystemer, mesoøkosystemer, makroøkosystemer og den globale biosfæren. Store terrestriske økosystemer kalles biomer. Økosystemer er ikke spredt i uorden, tvert imot er de gruppert i ganske regelmessige soner, både horisontalt (i breddegrad) og vertikalt (i høyde).
Fra ekvator til polene er en viss symmetri synlig i fordelingen av biomer av forskjellige halvkuler: tropiske regnskoger, ørkener, stepper, tempererte skoger, barskoger, taiga.
Hvert økosystem har to hovedkomponenter: organismer og faktorer fra deres ikke-levende miljø. Helheten av organismer (planter, dyr, mikrober) kalles biotaen til et økosystem. Fra synspunktet til den trofiske strukturen (fra den greske trofe - ernæring), kan økosystemet deles inn i to lag: øvre, nedre.
Følgende komponenter skilles ut som en del av et økosystem: uorganiske, organiske forbindelser, luft, vann og substratmiljøer, produsenter, autotrofe organismer, forbrukere eller fagotrofer, nedbrytere og detritivorer.
Solenergi på jorden forårsaker to sykluser av stoffer: store, eller geologiske, og små, biologiske (biotiske).
I et økosystem er det sykluser av stoffer. De mest studerte er: syklusene av karbon, oksygen, nitrogen, fosfor, svovel, etc.
Opprettholdelsen av den vitale aktiviteten til organismer og sirkulasjonen av materie i økosystemene, det vil si eksistensen av økosystemer, avhenger av den konstante tilstrømningen av solenergi.
I økosystemer skjer det konstant endringer i tilstanden og vital aktivitet til medlemmene deres og i forholdet mellom populasjoner. De forskjellige endringene som skjer i ethvert samfunn faller inn i to hovedtyper: sykliske og progressive.
Et globalt økosystem er en biosfære preget av integriteten og stabiliteten til økosystemene.
1. Konsept og struktur av økosystemer
1 Begrepet økosystemer
Levende organismer og deres ikke-levende (abiotiske) miljø er uatskillelig forbundet med hverandre og er i konstant interaksjon. Enhver enhet (biosystem) i et økosystem er et økologisk system. Et økologisk system, eller økosystem, er den grunnleggende funksjonelle enheten i økologi, siden det inkluderer organismer og det livløse miljøet - komponenter som gjensidig påvirker hverandres egenskaper og de nødvendige betingelsene for å opprettholde liv i den formen som finnes på jorden. Begrepet «økosystem» ble først foreslått i 1935 av den engelske økologen A. Tansley. For tiden er følgende definisjon av økosystem mye brukt. Et økosystem er enhver samling av organismer og uorganiske komponenter der sirkulasjon av stoffer kan forekomme. Ifølge N.F. Reimers (1990), et økosystem er ethvert fellesskap av levende vesener og dets habitat, forent til en enkelt funksjonell helhet, som oppstår på grunnlag av gjensidig avhengighet og årsak-og-virkning-forhold som eksisterer mellom individuelle miljøkomponenter. Det bør understrekes at kombinasjonen av et spesifikt fysisk-kjemisk miljø (biotop) med et fellesskap av levende organismer (biocenose) danner et økosystem. A. Tansley (1935) foreslo følgende forhold:
Økosystem = Biotop + Biocenose
I henhold til definisjonen av V.N. Sukachev, biogeocenosis er "et sett av homogene naturfenomener (atmosfære, stein, jord og hydrologiske forhold) over en viss utstrekning av jordoverflaten, som har sin egen spesielle spesifisitet av interaksjoner mellom disse komponentene som utgjør den og en viss type utveksling av materie og energi mellom seg selv og andre naturfenomener og representerer en internt motstridende dialektisk enhet, i konstant bevegelse og utvikling."
I tillegg til begrepene økosystem A. Tensley og biogeocenosis V.N. Sukachev formulerte regelen til F. Evans (1956), som foreslo å bruke begrepet "økosystem" absolutt "dimensjonsløst" for å betegne ethvert supraorganismalt levende system som samhandler med miljøet. Imidlertid ga mange forfattere begrepet "økosystem" betydningen av biogeocenose, dvs. elementært økosystem, og samtidig høyere i hierarkiet av supra-biogeokoenotiske formasjoner opp til biosfærens økosystem.
2 Klassifisering av økosystemer
Økosystemene som eksisterer på jorden er forskjellige. Det er mikroøkosystemer (for eksempel stammen til et råtnende tre), mesoøkosystemer (skog, dam, etc.), makroøkosystemer (kontinent, hav, etc.) og den globale biosfæren.
Store terrestriske økosystemer kalles biomer. Hvert biom inneholder en rekke mindre, sammenkoblede økosystemer. Det er flere klassifiseringer av økosystemer:
Eviggrønn tropisk regnskog
Ørken: Gras og busk Chaparral - områder med regnfulle vintre og tørre somre
Tropisk Graslenz og Savannah
Temperert steppe
Temperert edelløvskog
Boreale barskoger
Tundra: arktisk og alpint
Typer ferskvannsøkosystemer:
Bånd (stille vann): innsjøer, dammer, etc.
Lotic (rennende vann): elver, bekker, etc.
Våtmarker: sumper og sumpete skoger
Typer marine økosystemer
Åpent hav (pelagisk)
Kontinentalsokkelvann (kystvann)
Oppvekstområder (fruktbare områder med produktivt fiskeri)
Elvemunninger (kystbukter, sund, elvemunninger, saltmyrer osv.). Terrestriske biomer kjennetegnes her av naturlige eller opprinnelige vegetasjonstrekk, og typer vannøkosystemer kjennetegnes ved geologiske og fysiske trekk. De listede hovedtypene av økosystemer representerer miljøet der menneskelig sivilisasjon utviklet seg og representerer de viktigste biotiske samfunnene som støtter livet på jorden.
3 Sonering av makroøkosystemer
Studiet av den geografiske fordelingen av økosystemer kan bare utføres på nivå med store økologiske enheter - makroøkosystemer, som vurderes på kontinental skala. Økosystemer er ikke spredt i uorden, tvert imot er de gruppert i ganske regelmessige soner, både horisontalt (i breddegrad) og vertikalt (i høyde). Dette bekreftes av den periodiske loven om geografisk sonering til A.A. Grigorieva - M.I. Budyko: med endring av fysisk-geografiske soner på jorden, gjentas lignende landskapssoner og noen av deres generelle egenskaper med jevne mellomrom. Dette ble også diskutert når man vurderte livets bakke-luft-miljø. Periodisiteten etablert ved lov manifesteres i det faktum at verdiene til tørrhetsindeksen varierer i forskjellige soner fra 0 til 4-5, tre ganger mellom polene og ekvator er de nær enhet. Disse verdiene tilsvarer den høyeste biologiske produktiviteten til landskap.
Den periodiske repetisjonen av egenskaper i en serie systemer på ett hierarkisk nivå er sannsynligvis en generell lov for universet, formulert som loven om periodisitet i strukturen til systemiske aggregater, eller en system-periodisk lov - spesifikke naturlige systemer på ett nivå ( undernivå) av organisasjon utgjør en periodisk eller repeterende serie av morfologisk like strukturer innenfor øvre og nedre systemrom-tid-grenser, utover hvilke eksistensen av systemer på et gitt nivå blir umulig. De går inn i en ustabil tilstand eller blir til en annen systemstruktur, inkludert et annet organisasjonsnivå.
Temperatur og nedbør bestemmer plasseringen av de viktigste terrestriske biomene på jordens overflate. Mønsteret av temperatur og nedbør i et bestemt område over ganske lang tid er det vi kaller klima. Klimaet i forskjellige regioner på kloden er forskjellig. Årsnedbøren varierer fra 0 til 2500 mm eller mer. Regimer for temperatur og nedbør kombineres på svært forskjellige måter.
Spesifikasjonene til klimatiske forhold bestemmer på sin side utviklingen av et bestemt biom.
Fra ekvator til polene er en viss symmetri synlig i fordelingen av biomer av forskjellige halvkuler:
Tropiske regnskoger (nordlige Sør-Amerika, Mellom-Amerika, vestlige og sentrale deler av ekvatorial-Afrika, Sørøst-Asia, kystområder i det nordvestlige Australia, øyer i Det indiske hav og Stillehavet). Klimaet er uten årstider (nærhet til ekvator), den gjennomsnittlige årlige temperaturen er over 17°C (vanligvis 28°C), den gjennomsnittlige årlige nedbøren overstiger 2400 mm.
Vegetasjon: skog dominerer. Noen typer trær er opptil 60 m høye. På stammene og grenene deres er det epifytiske planter, hvis røtter ikke når jorda, og treaktige vinranker som slår rot i jorda og klatrer opp i trærne til toppen. Alt dette danner en tykk baldakin.
Fauna: artssammensetningen er rikere enn i alle andre biomer til sammen. Spesielt mange er amfibier, krypdyr og fugler (frosker, øgler, slanger, papegøyer), aper og andre små pattedyr, eksotiske insekter med lyse farger og fargerike fisker i reservoarer.
Funksjoner: jordsmonn er ofte tynn og dårlig, de fleste næringsstoffene finnes i overflatebiomassen til rotfestet vegetasjon.
2. Savannas (subequatorial Afrika, Sør-Amerika, en betydelig del av det sørlige India). Klimaet er tørt og varmt det meste av året. Kraftig nedbør i den våte årstiden. Temperatur: gjennomsnittlig årlig-høy. Nedbøren er 750-1650 mm/år, hovedsakelig i regntiden. Vegetasjon - blågress (korn)planter med sjeldne løvtrær. Fauna: store planteetende pattedyr, som antiloper, sebraer, sjiraffer, neshorn, rovdyr - løver, leoparder, geparder.
Ørkener (noen områder i Afrika, for eksempel Sahara; Midtøsten og Sentral-Asia, Great Basin og det sørvestlige USA og Nord-Mexico, etc.). Klimaet er veldig tørt. Temperatur - varme dager og kalde netter. Nedbør er mindre enn 250 mm/år. Vegetasjon: sparsomme busker, ofte tornete, noen ganger kaktus og lavt gress, som raskt dekker bakken med et blomstrende teppe etter sjeldent regn. Planter har omfattende overflaterotsystemer som fanger opp fuktighet fra sjelden nedbør, samt kranerøtter som trenger ned i bakken til grunnvannstanden (30 m og dypere). Fauna: ulike gnagere (kengururotter, etc.), padder, øgler, slanger og andre krypdyr, ugler, ørner, gribber, småfugler og insekter i store mengder.
4. Stepper (sentrum av Nord-Amerika, Russland, visse områder i Afrika og Australia, sørøst for Sør-Amerika). Klimaet er sesongbetont. Temperaturer - sommertemperaturer varierer fra moderat varme til varme, vintertemperaturer under 0°C. Nedbør - 750-2000 mm/år. Vegetasjon: dominert av blågress (korn) opp til 2 m og høyere i høyden i noen prærier i Nord-Amerika eller opptil 50 cm, for eksempel i de russiske steppene, med isolerte trær og busker i våte områder. Fauna: store planteetende pattedyr - bison, pronghorn antilope (Nord-Amerika), ville hester (Eurasia), kenguruer (Australia), sjiraffer, sebraer, hvite neshorn, antiloper (Afrika); Rovdyr inkluderer coyoter, løver, leoparder, geparder, hyener, en rekke fugler og små gravende pattedyr som kaniner, jordekorn og jordvarker.
5. Tempererte skoger (Vest-Europa, Øst-Asia, østlige USA). Klima - sesongmessig med vintertemperaturer under 0 0C. Nedbør - 750-2000 mm/år. Vegetasjon: dominert av skog med løvtrær opp til 35-45 m høye (eik, hickory, lønn), buskete undervegetasjon, moser, lav. Fauna: pattedyr (hvithalehjort, piggsvin, vaskebjørn, opossum, ekorn, kanin, spissmus), fugler (sanger, hakkespett, svarttrost, ugler, falker), slanger, frosker, salamandere, fisk (ørret, abbor, steinbit, etc. . ), rikelig jordmikrofauna (fig. 12.3). Biotaen er tilpasset sesongklimaet: dvalemodus, migrasjon, dvale i vintermånedene. 6. Barskoger, taiga (nordlige regioner i Nord-Amerika, Europa og Asia). Klimaet er lange og kalde vintre, med noe nedbør som faller i form av snø. Vegetasjon: eviggrønne barskoger dominerer, for det meste gran, furu og gran. Fauna: store planteetende hovdyr (muldyr, rein), små planteetende pattedyr (hare, ekorn, gnagere), ulv, gaupe, rev, svartbjørn, grizzlybjørn, jerv, mink og andre rovdyr, tallrike blodsugende insekter på kort sommer tid. Klimaet er veldig kaldt med polare dager og polare netter. Gjennomsnittlig årlig temperatur er under -5°C. I løpet av noen uker med kort sommer tiner ikke bakken mer enn én meter dypt. Nedbør er mindre enn 250 mm/år. Vegetasjon: dominert av saktevoksende lavarter, moser, gress og klosser, og dvergbusker. Fauna: store planteetende hovdyr (rein, moskus), små gravende pattedyr (hele året, for eksempel lemen), rovdyr som får en kamuflasjehvit farge om vinteren (fjellrev, gaupe, hermelin, snøugle). På den korte sommeren hekker et stort antall trekkfugler i tundraen, blant dem er det spesielt mange vannfugler, som lever av insektene og ferskvannsvirvelløse dyr som er rikelig her. Den vertikale høydesoneringen av landøkosystemer, spesielt på steder med utpreget relieff, er også veldig tydelig. Fuktighet er hovedfaktoren som bestemmer typen biom. Ved tilstrekkelig store mengder nedbør utvikles skogvegetasjon. Temperaturen bestemmer skogtypen. Situasjonen er nøyaktig den samme i steppe- og ørkenbiomene. Endringer i vegetasjonstyper i kalde strøk skjer med lavere årlige nedbørsmengder, siden det ved lave temperaturer går mindre vann tapt ved fordampning. Temperatur blir en viktig faktor bare under svært kalde forhold med permafrost. Dermed avhenger sammensetningen av økosystemer i stor grad av deres funksjonelle "formål" og omvendt. Ifølge N.F. Reimers (1994), dette gjenspeiles i prinsippet om økologisk komplementaritet (komplementaritet): ingen funksjonell del av økosystemet (økologisk komponent, element osv.) kan eksistere uten andre funksjonelt komplementære deler. Nær det og utvide det er prinsippet om økologisk kongruens (korrespondanse):. funksjonelt utfyller hverandre, utvikler de levende komponentene i økosystemet passende tilpasninger for dette, koordinert med forholdene i det abiotiske miljøet, som i stor grad transformeres av de samme organismene (bioklima, etc.), dvs. det er en dobbel serie av korrespondanse - mellom organismer og deres habitat - eksternt og skapt av cenosis. 4 Økosystemstruktur Fra synspunktet til den trofiske strukturen (fra den greske trofe - ernæring), kan økosystemet deles inn i to lag: Det øvre - autotrofe (selvmatende) nivået, eller "grønt belte", inkludert planter eller deres deler som inneholder klorofyll, hvor fiksering av telleenergi og bruk av enkle uorganiske forbindelser dominerer. Den nederste er det heterotrofe (matet av andre) laget, eller det "brune beltet" av jordsmonn og sedimenter, råtnende stoffer, røtter osv., der bruk, transformasjon og nedbrytning av komplekse forbindelser dominerer. Fra et biologisk synspunkt skilles følgende komponenter ut som en del av et økosystem: I et økosystem er mat- og energiforbindelser mellom kategorier alltid entydige og går i retning av: autotrofer - heterotrofer eller i en mer fullstendig form; autotrofer -> forbrukere -> nedbrytere (destruktorer). Den primære energikilden for økosystemer er solen. Energistrømmen (ifølge T.A. Akimova, V.V. Khaskin, 1994) sendt av solen til planeten Jorden overstiger 20 millioner EJ per år. Bare en fjerdedel av denne strømmen nærmer seg grensen til atmosfæren. Av dette reflekteres omtrent 70 %, absorberes av atmosfæren og sendes ut i form av langbølget infrarød stråling. Solstråling som faller på jordoverflaten er 1,54 millioner EJ per år. Denne enorme energimengden er 5000 ganger hele menneskehetens energi på slutten av det 20. århundre og 5,5 ganger energien til alle tilgjengelige fossile brenselressurser av organisk opprinnelse, akkumulert over minst 100 millioner år. Mesteparten av solenergien som når planetens overflate omdannes direkte til varme, varmende vann eller jord, som igjen varmer opp luften. Denne varmen fungerer som drivkraften bak vannets syklus, luftstrømmer og havstrømmer som bestemmer været, og slippes gradvis ut i verdensrommet, hvor den går tapt. For å bestemme økosystemenes plass i denne naturlige strømmen av energi, er det viktig å innse at uansett hvor omfattende og komplekse de er, bruker de bare en liten del av den. Dette innebærer et av de grunnleggende prinsippene for økosystemenes funksjon: de eksisterer på grunn av ikke-forurensende og nesten evig solenergi, hvis mengde er relativt konstant og rikelig. La oss gi mer detaljert hver av de listede egenskapene til solenergi: 2. Faktorer som sikrer integriteten til økosystemene 1 Stoffsykluser Begge syklusene er gjensidig forbundet og representerer så å si en enkelt prosess. Det anslås at alt oksygenet i atmosfæren går gjennom organismer (kombinert under respirasjon og frigjort under fotosyntese) i 2000 år, karbondioksidet i atmosfæren sykluser i motsatt retning i 300 år, og alt vannet på jorden er dekomponert og gjenskapt gjennom fotosyntese og respirasjon på 2 000 000 år. Samspillet mellom abiotiske faktorer og levende organismer i økosystemet er ledsaget av en kontinuerlig sirkulasjon av materie mellom biotopen og biocenosen i form av alternerende organiske og mineralske forbindelser. Utvekslingen av kjemiske elementer mellom levende organismer og det uorganiske miljøet, hvis ulike stadier forekommer innenfor et økosystem, kalles biogeokjemisk sirkulasjon, eller biogeokjemisk syklus. Vannets kretsløp. Den mest betydningsfulle syklusen på jorden når det gjelder overførte masser og energiforbruk er den planetariske hydrologiske syklusen - vannets syklus. Hvert sekund er 16,5 millioner m3 vann involvert i det og mer enn 40 milliarder MW solenergi brukes på dette, (ifølge T.A. Akimova V.V. Haskin, (1994)). Men denne syklusen er ikke bare overføring av vannmasser. Dette er fasetransformasjoner, dannelse av løsninger og suspensjoner, utfelling, krystallisering, fotosynteseprosesser, samt ulike kjemiske reaksjoner. Livet oppsto og fortsetter i dette miljøet. Vann er det grunnleggende elementet som er nødvendig for liv. Kvantitativt er det den vanligste uorganiske komponenten i levende materie. Hos mennesker utgjør vann 63% av kroppsvekten, i sopp - 80%, i planter - 80-90%, og hos noen maneter - 98% Vann, som vi vil se litt senere, deltar i den biologiske syklusen og tjener som en kilde til hydrogen og oksygen, og utgjør bare en liten del av det totale volumet. I flytende, faste og dampede tilstander er vann tilstede i alle de tre hovedkomponentene i biosfæren: atmosfæren, hydrosfæren og litosfæren. Alle farvann er forent av det vanlige konseptet "hydrosfære". Komponentene i hydrosfæren er sammenkoblet ved konstant utveksling og interaksjon. Vann, som kontinuerlig beveger seg fra en tilstand til en annen, gjør små og store sykluser. Fordampningen av vann fra havoverflaten, kondenseringen av vanndamp i atmosfæren og nedbøren på havoverflaten danner en liten syklus. Når vanndamp føres med luftstrømmer til land, blir syklusen mye mer kompleks. I dette tilfellet fordamper en del av nedbøren og går tilbake til atmosfæren, den andre mater elver og reservoarer, men går til slutt tilbake til havet gjennom elv og underjordisk avrenning, og fullfører dermed den store syklusen. Biotisk (biologisk) syklus. Den biotiske (biologiske) syklusen refererer til sirkulasjonen av stoffer mellom jord, planter, dyr og mikroorganismer. Den biotiske (biologiske) syklusen er strømmen av kjemiske elementer fra jord, vann og atmosfære til levende organismer, transformasjon av innkommende elementer til nye komplekse forbindelser og deres retur tilbake i livsprosessen med det årlige fallet av en del av det organiske materie eller med helt døde organismer inkludert i økosystemsammensetningen. Nå skal vi presentere den biotiske syklusen i en syklisk form. Den sentrale biotiske syklusen (ifølge T.A. Akimova, V.V., Khaskhin) besto av primitive encellede produsenter (P) og nedbrytere-destruktorer (D). Mikroorganismer er i stand til raskt å formere seg og tilpasse seg forskjellige forhold, for eksempel bruker de alle slags substrater - karbonkilder - i kostholdet. Høyere organismer har ikke slike evner. I komplette økosystemer kan de eksistere i form av en struktur på grunnlag av mikroorganismer. Først utvikles flercellede planter (P) - høyere produsenter. Sammen med encellede organismer skaper de organisk materiale gjennom prosessen med fotosyntese, ved å bruke energien fra solstråling. Deretter kobles primære forbrukere sammen - planteetende dyr (T), og deretter kjøttetende forbrukere. Vi undersøkte landets biotiske syklus. Dette gjelder fullt ut den biotiske syklusen til akvatiske økosystemer, for eksempel havet. Alle organismer inntar en viss plass i den biotiske syklusen og utfører sine funksjoner med å transformere grenene av energistrømmen som de mottar og overføre biomasse. Et system med encellede nedbrytere (destruktorer) forener alle, depersonaliserer stoffene deres og lukker den generelle sirkelen. De returnerer til det abiotiske miljøet i biosfæren alle elementene som er nødvendige for nye og nye revolusjoner. De viktigste trekkene ved den biotiske syklusen bør vektlegges. Fotosyntese er en kraftig naturlig prosess som årlig involverer enorme masser av biosfærestoff i syklusen og bestemmer dets høye oksygenpotensial. På grunn av karbondioksid og vann syntetiseres organisk materiale og fritt oksygen frigjøres. De direkte produktene av fotosyntesen er forskjellige organiske forbindelser, og generelt er prosessen med fotosyntese ganske kompleks. I tillegg til fotosyntese med deltagelse av oksygen, den såkalte oksygeniske fotosyntesen, bør vi også fokusere på oksygenfri fotosyntese, eller kjemosyntese. Kjemosyntetiske organismer inkluderer nitrifikatorer, karboksydobakterier, svovelbakterier, tioniske jernbakterier og hydrogenbakterier. De er oppkalt etter deres oksidasjonssubstrater, som kan være NH3, NO2, CO, H2S, S, Fe2+, H2. Noen arter er obligatoriske kjemolitoautotrofer, andre er fakultative. Sistnevnte inkluderer karboksydobakterier og hydrogenbakterier. Kjemosyntese er karakteristisk for dypvanns hydrotermiske ventiler. Fotosyntese forekommer, med få unntak, på hele jordens overflate, skaper en enorm geokjemisk effekt og kan uttrykkes som mengden av hele massen av karbon som årlig er involvert i konstruksjonen av organisk - levende materiale i hele biosfæren. Den generelle syklusen av materie knyttet til konstruksjon av organisk materiale gjennom fotosyntese involverer også kjemiske elementer som N, P, S, samt metaller - K, Ca, Mg, Na, Al. Når en organisme dør, skjer den omvendte prosessen - nedbrytning av organisk materiale gjennom oksidasjon, forfall, etc. med dannelse av endelige nedbrytningsprodukter. I jordens biosfære fører denne prosessen til at mengden biomasse av levende stoffer har en tendens til å være noe konstant. Biomasse i økosfæren (2 10|2t) er syv størrelsesordener mindre enn massen til jordskorpen (2,10|9t). Jordens planter produserer årlig organisk materiale tilsvarende 1.6.10"% eller 8% av biomassen i økosfæren. Destruktorer, som utgjør mindre enn 1% av den totale biomassen til planetens organismer, behandler en masse organisk materiale som er 10 ganger større enn deres egen biomasse. I gjennomsnitt er biomassefornyelsesperioden 12,5 år La oss anta at massen av levende stoff og produktiviteten til biosfæren var den samme fra kambrium til i dag (530 millioner år), da den totale mengden organisk materiale som passerte gjennom den globale biotiske syklusen og ble brukt av livet på planeten vil være 2.10" 2-5,ZL08/12.5=8.5L0|9t, som er 4 ganger massen av jordskorpen. Når det gjelder disse beregningene N.S. Pechurkin (1988) skrev: "Vi kan si at atomene som utgjør kroppene våre var i eldgamle bakterier, og i dinosaurer og i mammuter." Loven om biogen migrasjon av atomer V.I. Vernadsky uttaler: "Migrasjonen av kjemiske elementer på jordens overflate og i biosfæren som helhet skjer enten med direkte deltakelse av levende materie (biogen migrasjon), eller den skjer i et miljø med geokjemiske egenskaper (O2, CO2, H2, etc.) er bestemt levende materie, både det som for tiden bor i biosfæren og det som har virket på jorden gjennom geologisk historie." I OG. Vernadsky i 1928-1930 i sine dype generaliseringer angående prosesser i biosfæren ga han en idé om de fem biogeokjemiske hovedfunksjonene til levende materie. Den første funksjonen er gass. Den andre funksjonen er konsentrasjon. Den tredje funksjonen er redoks. Den fjerde funksjonen er biokjemisk. Den femte funksjonen er menneskehetens biogeokjemiske aktivitet, som dekker en stadig økende mengde materie i jordskorpen for behovene til industri, transport og landbruk. Den biologiske syklusen varierer i ulike naturlige soner og er klassifisert i henhold til et sett med indikatorer: plantebiomasse, søppel, søppel, mengden elementer som er festet i biomassen, etc. Den totale biomassen er høyest i skogsonen, og andelen underjordiske organer i skog er lavest. Dette bekreftes av den biologiske syklusintensitetsindeksen - forholdet mellom massen til kullet og den delen av kullet som danner det. Karbon syklus. Av alle biogeokjemiske sykluser er karbonsyklusen uten tvil den mest intense. Karbon sirkulerer med høy hastighet mellom ulike uorganiske medier og gjennom næringsnett i samfunn av levende organismer. CO og CO2 spiller en viss rolle i karbonkretsløpet. Ofte i jordens biosfære er karbon representert av den mest mobile formen for CO2. Kilden til primær karbondioksid i biosfæren er vulkansk aktivitet assosiert med sekulær avgassing av mantelen og nedre horisonter av jordskorpen. Migrering av CO2 i biosfæren skjer på to måter. Den første måten er å absorbere det under fotosyntesen med dannelse av glukose og andre organiske stoffer som alt plantevev er bygget av. De blir deretter transportert gjennom næringskjeder og danner vev til alle andre levende vesener i økosystemet. Når planter og dyr dør på overflaten, skjer oksidasjon av organiske stoffer med dannelse av CO2. Karbonatomer returneres også til atmosfæren når organisk materiale brennes. Et viktig og interessant trekk ved karbonkretsløpet er at i fjerne geologiske epoker, for hundrevis av millioner av år siden, ble en betydelig del av det organiske materialet som ble skapt i fotosynteseprosessene ikke brukt av verken forbrukere eller nedbrytere, men akkumulert i litosfæren. i form av fossilt brensel: olje, kull, oljeskifer, torv osv. Disse fossile brenselene utvinnes i enorme mengder for å dekke energibehovet i vårt industrisamfunn. Ved å brenne det, fullfører vi på en måte karbonkretsløpet. På den andre måten utføres karbonmigrering ved å lage et karbonatsystem i ulike reservoarer, hvor CO2 omdannes til H2CO3, HCO, CO3. Ved hjelp av kalsium (eller magnesium) oppløst i vann utfelles karbonater (CaCO3) gjennom biogene og abiogene veier. Det dannes tykke lag av kalkstein. Ifølge A.B. Ronov, forholdet mellom nedgravd karbon i fotosyntetiske produkter og karbon i karbonatbergarter er 1:4. Sammen med det store karbonkretsløpet er det også en rekke små karbonkretsløp på landoverflaten og i havet. Generelt, uten menneskeskapt intervensjon, opprettholdes karboninnholdet i biogeokjemiske reservoarer: biosfæren (biomasse + jord og detritus), sedimentære bergarter, atmosfære og hydrosfære med en høy grad av konstanthet (ifølge T.A. Akimova, V.V. Haskin (1994) ). Den konstante utvekslingen av karbon, på den ene siden, mellom biosfæren, og på den andre, mellom atmosfæren og hydrosfæren, bestemmes av gassfunksjonen til levende materie - prosessene med fotosyntese, respirasjon og ødeleggelse, og utgjør ca. 6-1010 tonn/år Det er en strøm av karbon inn i atmosfæren og hydrosfæren og under vulkansk aktivitet i gjennomsnitt 4,5 106 t/år. Den totale massen av karbon i fossilt brensel (olje, gass, kull osv.) er beregnet til 3,2*1015 tonn, som tilsvarer en gjennomsnittlig akkumuleringshastighet på 7 millioner tonn/år. Denne mengden er ubetydelig sammenlignet med massen av sirkulerende karbon og falt som det var ut av syklusen og gikk tapt i den. Derfor er graden av åpenhet (ufullkommenhet) av syklusen 10"4, eller 0,01%, og følgelig er lukkegraden 99,99%. Dette betyr på den ene siden at hvert karbonatom deltok i syklusen. titusenvis av ganger før de falt ut av syklusen, havnet i dypet.På den annen side er strømmene av syntese og forfall av organiske stoffer i biosfæren tilpasset hverandre med meget høy presisjon. 0,2 % av det mobile karbonlageret er i konstant sirkulasjon. Biomassekarbon fornyes om 12 år, i atmosfæren - om 8 år. Oksygen syklus. Oksygen (O2) spiller en viktig rolle i livet til de fleste levende organismer på planeten vår. I kvantitative termer er dette hovedkomponenten i levende materie. For eksempel, hvis vi tar hensyn til vannet i vev, inneholder menneskekroppen 62,8% oksygen og 19,4% karbon. Generelt er dette elementet i biosfæren, sammenlignet med karbon og hydrogen, det viktigste blant enkle stoffer. Innenfor biosfæren skjer det en rask utveksling av oksygen med levende organismer eller deres levninger etter døden. Planter produserer som regel fritt oksygen, og dyr konsumerer det gjennom respirasjon. Som det mest utbredte og mobile elementet på jorden, begrenser ikke oksygen eksistensen og funksjonene til økosfæren, selv om tilgjengeligheten av oksygen for vannlevende organismer kan være midlertidig begrenset. Oksygensyklusen i biosfæren er ekstremt kompleks, siden et stort antall organiske og uorganiske stoffer reagerer med den. Som et resultat oppstår mange episykler, som forekommer mellom litosfæren og atmosfæren eller mellom hydrosfæren og disse to miljøene. Oksygensyklusen ligner på noen måter den omvendte karbondioksidsyklusen. Bevegelsen til den ene skjer i motsatt retning av bevegelsen til den andre. Forbruket av atmosfærisk oksygen og dets erstatning av primærprodusenter skjer relativt raskt. Dermed tar det 2000 år å fullstendig fornye alt atmosfærisk oksygen. I dag forekommer ikke oksygenakkumulering i atmosfæren, og innholdet (20,946%) forblir konstant. I de øvre lagene av atmosfæren, når ultrafiolett stråling virker på oksygen, dannes ozon - O3 -. Omtrent 5% av solenergien som når jorden brukes på dannelsen av ozon - omtrent 8,6 * 1015 W. Reaksjonene er lett reversible. Når ozon brytes ned, frigjøres denne energien, som holder høye temperaturer i den øvre atmosfæren. Den gjennomsnittlige ozonkonsentrasjonen i atmosfæren er omtrent 106 vol. %; den maksimale O3-konsentrasjonen - opptil 4-10 "* vol.% - oppnås i høyder på 20-25 km (T.A. Akimova, V.V. Haskin, 1998). Ozon fungerer som et slags UV-filter: det blokkerer en betydelig del av harde ultrafiolette stråler. Sannsynligvis var dannelsen av ozonlaget en av betingelsene for at liv kunne komme ut av havet og kolonisere land. Det meste av oksygenet som ble produsert under geologiske epoker forble ikke i atmosfæren, men ble fikset av litosfæren i form av karbonater, sulfater, jernoksider, etc. Denne massen er 590 * 1014 tonn mot 39 * 1014 tonn oksygen, som sirkulerer i biosfæren i form av gass eller sulfater oppløst i kontinentale og oceaniske farvann. Nitrogen syklus. Nitrogen er et essensielt biogent element, siden det er en del av proteiner og nukleinsyrer. Nitrogensyklusen er en av de mest komplekse, siden den inkluderer både gass- og mineralfaser, og samtidig de mest ideelle syklusene. Nitrogenkretsløpet er nært knyttet til karbonkretsløpet. Som regel følger nitrogen karbon, sammen med hvilket det deltar i dannelsen av alle proteinstoffer. Atmosfærisk luft, som inneholder 78 % nitrogen, er et uuttømmelig reservoar. Imidlertid kan flertallet av levende organismer ikke direkte bruke dette nitrogenet. For at nitrogen skal tas opp av planter, må det være en del av ammonium- (NH*) eller nitrat-ioner (NO3). Nitrogengass slippes kontinuerlig ut i atmosfæren som et resultat av arbeidet med å denitrifisere bakterier, og fiksering av bakterier, sammen med blågrønne alger (cyanofytter), absorberer den hele tiden og omdanner den til nitrater. Nitrogensyklusen er tydelig synlig på nivået av destruktorer. Proteiner og andre former for organisk nitrogen som finnes i planter og dyr blir etter deres død utsatt for heterotrofe bakterier, actinomycetes, sopp (bioreduserende mikroorganismer), som produserer energien de trenger ved å redusere dette organiske nitrogenet og omdanne det til ammoniakk. I jordsmonn oppstår prosessen med nitrifikasjon, bestående av en kjede av reaksjoner, hvor, med deltakelse av mikroorganismer, skjer oksidasjon av ammoniumion (NH4+) til nitritt (NO~) eller nitritt til nitrat (N0~). Reduksjonen av nitritter og nitrater til de gassformige forbindelsene molekylært nitrogen (N2) eller lystgass (N20) er essensen av denitrifikasjonsprosessen. Dannelsen av uorganiske nitrater i små mengder skjer hele tiden i atmosfæren ved å binde atmosfærisk nitrogen med oksygen under elektriske utladninger under tordenvær, og deretter falle med regn ned på jordoverflaten. En annen kilde til atmosfærisk nitrogen er vulkaner, som kompenserer for tapet av nitrogen utelukket fra syklusen under sedimentering eller avsetning til bunnen av havene. Generelt er gjennomsnittlig tilførsel av nitratnitrogen av abiotisk opprinnelse under avsetning fra atmosfæren til jorda ikke over 10 kg (år/ha), frie bakterier produserer 25 kg (år/ha), mens symbiosen av Rhizobium med belgplanter produserer i snitt 200 kg (år/ha). Den overveiende delen av det fikserte nitrogenet behandles ved å denitrifisere bakterier til N2 og returneres til atmosfæren. Bare rundt 10 % av ammonifisert og nitrifisert nitrogen absorberes fra jorda av høyere planter og ender opp til disposisjon for flercellede representanter for biocenoser Fosfor syklus. Fosforsyklusen i biosfæren er assosiert med metabolske prosesser i planter og dyr. Dette viktige og nødvendige elementet i protoplasma, inneholdt i landplanter og alger 0,01-0,1%, dyr fra 0,1% til flere prosent, sirkulerer, og blir gradvis fra organiske forbindelser til fosfater, som igjen kan brukes av planter. Fosfor, i motsetning til andre biofile elementer, danner imidlertid ikke en gassform under migrasjon. Reservoaret av fosfor er ikke atmosfæren, som nitrogen, men den mineralske delen av litosfæren. De viktigste kildene til uorganisk fosfor er magmatiske bergarter (apatitter) eller sedimentære bergarter (fosforitter). Fra bergarter er uorganisk fosfor involvert i sirkulasjon ved utvasking og oppløsning i kontinentalt farvann. Når det kommer inn i terrestriske økosystemer og jord, absorberes fosfor av planter fra en vandig løsning i form av uorganisk fosfation og er inkludert i forskjellige organiske forbindelser, hvor det vises i form av organisk fosfat. Fosfor beveger seg gjennom næringskjeder fra planter til andre organismer i økosystemet. Fosfor transporteres inn i akvatiske økosystemer av rennende vann. Elver beriker kontinuerlig havene med fosfater. Der fosfor blir en del av planteplankton. Noen fosforforbindelser migrerer innenfor grunne dyp og blir konsumert av organismer, mens den andre delen går tapt på større dyp. Døde rester av organismer fører til akkumulering av fosfor på forskjellige dyp. Svovel syklus. Det er mange gassformige svovelforbindelser, som hydrogensulfid H2S og svoveldioksid SO2. Den dominerende delen av syklusen til dette grunnstoffet er imidlertid sedimentær i naturen og forekommer i jord og vann Tilgjengeligheten av uorganisk svovel i økosystemet er lettet av den gode løseligheten til mange sulfater i vann. Planter, absorberer sulfater, reduserer dem og produserer svovelholdige aminosyrer (metionin, cystein, cystin) som spiller en viktig rolle i utviklingen av den tertiære strukturen til proteiner under dannelsen av disulfidbroer mellom forskjellige soner i polypeptidkjeden. Mange grunnleggende trekk ved den biogeokjemiske syklusen er tydelig synlige: S -> S -> SO2 - fargeløse, grønne og lilla svovelbakterier; - "H2S (anaerob reduksjon av sulfat) - Desulfovibno; H2S - "SO2" (aerob oksidasjon av sulfid) - thiobacillus; organisk S i SO og H2S. - aerobe og anaerobe heterotrofe mikroorganismer, henholdsvis. Primærproduksjon sikrer inkorporering av sulfat i organisk materiale, og utskillelse av dyr fungerer som en måte å returnere sulfat til syklusen. Generelt krever økosystemet mindre svovel sammenlignet med nitrogen og fosfor. Derfor er svovel sjeldnere en begrensende faktor for planter og dyr. Samtidig er svovelsyklusen en nøkkel i den totale prosessen med biomasseproduksjon og nedbrytning. For eksempel, når jernsulfider dannes i sedimenter, overføres fosfor fra en uløselig form til en løselig form og blir tilgjengelig for organismer. Dette er en bekreftelse på hvordan en syklus reguleres av en annen. 2 Energiflyt i økosystemer I motsetning til stoffer som kontinuerlig sirkulerer gjennom ulike blokker av økosystemet, som alltid kan gjenbrukes og gå inn i syklusen, kan energi brukes én gang, dvs. det er en lineær strøm av energi gjennom økosystemet. Enveistilstrømningen av energi som et universelt naturfenomen oppstår som et resultat av termodynamikkens lover. Den første loven sier at energi kan omdannes fra en form (som lys) til en annen (som den potensielle energien til mat), men kan ikke skapes eller ødelegges. Den andre loven sier at det ikke kan være en enkelt prosess forbundet med transformasjonen av energi uten å miste noe av den. En viss mengde energi i slike transformasjoner spres til utilgjengelig termisk energi og går derfor tapt. Derfor: det kan ikke være transformasjoner, for eksempel av matstoffer til stoffet som utgjør kroppen til organismen, som skjer med 100 % effektivitet. Dermed er levende organismer energiomdannere. Og hver gang energi omdannes, går en del av den tapt i form av varme. Til syvende og sist blir all energien som kommer inn i den biotiske syklusen til et økosystem bort som varme. Levende organismer bruker faktisk ikke varme som energikilde for å utføre arbeid – de bruker lys og kjemisk energi. Næringskjeder og nettverk, trofiske nivåer. Innenfor et økosystem skapes energiholdige stoffer av autotrofe organismer og tjener som mat for heterotrofer. Matforbindelser er mekanismer for å overføre energi fra en organisme til en annen. Et typisk eksempel: et dyr spiser planter. Dette dyret kan på sin side spises av et annet dyr. På denne måten kan energi overføres gjennom en rekke organismer - hver påfølgende lever av den forrige, som forsyner den med råvarer og energi. Denne sekvensen av energioverføring kalles en næringskjede (trofisk) eller næringskjede. Plasseringen av hvert ledd i næringskjeden er et trofisk nivå. Det første trofiske nivået, som nevnt tidligere, er okkupert av autotrofer, eller såkalte primærprodusenter. Organismer på det andre trofiske nivået kalles primære forbrukere, de tredje - sekundære forbrukere, etc. Det er generelt tre typer næringskjeder: kjøttetende næringskjeder starter med planter og går fra små organismer til stadig større organismer. På land består næringskjeder av tre til fire ledd. En av de enkleste næringskjedene ser slik ut: plante -> hare -> ulv produsent -" planteeter -> -> rovdyr Følgende næringskjeder er også utbredt: Plantemateriale (f.eks. nektar) - "flue -" -" edderkopp -> spissmus -> ugle. Rosebuskesaft -> bladlus -> -> marihøne -> -> edderkopp - "insektspisende fugl -> rovfugl. I akvatiske og spesielt marine økosystemer har rovdyrnæringskjeder en tendens til å være lengre enn i landbaserte. Den tredje typen næringskjeder, som starter med døde planterester, kadaver og dyreekskrement, omtales som detritale (saprofytiske) næringskjeder eller detritale nedbrytningskjeder. Løvskog spiller en viktig rolle i de skadelige næringskjedene til terrestriske økosystemer, hvor det meste av løvet ikke blir konsumert av planteetere og er en del av søppelet av falne løv. Bladene knuses av mange detritivorer - sopp, bakterier, insekter (for eksempel collembola), etc., og deretter inntas av meitemark, som jevnt fordeler humus i overflatelaget av jorden, og danner den såkalte mull. På dette nivået utvikler sopp mycel. De nedbrytende mikroorganismene som fullfører kjeden produserer den endelige mineraliseringen av dødt organisk materiale. Generelt kan typiske skadelige næringskjeder i skogene våre representeres som følger: Bladstrø -> meitemark -> svarttrost - spurvehauk; Dødt dyr - "larver av ådselfluer -" gressfrosk -> vanlig gressslange. I næringskjedediagrammene som er diskutert, er hver organisme representert som fôring av andre organismer av en type. De faktiske matforbindelsene i et økosystem er mye mer komplekse, siden et dyr kan livnære seg på organismer av forskjellige typer fra samme næringskjede eller fra forskjellige næringskjeder, for eksempel rovdyr på de øvre trofiske nivåene. Dyr lever ofte av både planter og andre dyr. De kalles altetende. Næringsvevene i økosystemene er svært komplekse, og vi kan konkludere med at energien som kommer inn i dem tar lang tid å migrere fra en organisme til en annen. Økologiske pyramider. Innenfor hvert økosystem har næringsnettene en veldefinert struktur, som er preget av arten og antallet organismer representert på hvert nivå i de ulike næringskjedene. For å studere forholdet mellom organismer i et økosystem og for å skildre dem grafisk, bruker de vanligvis økologiske pyramider i stedet for næringsnettdiagrammer. Økologiske pyramider uttrykker den trofiske strukturen til et økosystem i geometrisk form. De er konstruert i form av rektangler med samme bredde, men lengden på rektanglene må være proporsjonal med verdien av objektet som måles. Herfra kan du hente pyramider av tall, biomasse og energi. Økologiske pyramider gjenspeiler de grunnleggende egenskapene til enhver biocenose når de viser dens trofiske struktur. Pyramider av tall. De representerer den enkleste tilnærmingen til studiet av den trofiske strukturen til et økosystem. Biomassepyramide. Gjenspeiler mer fullstendig ernæringsforholdene i økosystemet, siden det tar hensyn til den totale massen av organismer (biomasse) for hvert trofisk nivå. Pyramide av energi. Den mest grunnleggende måten å vise forbindelser mellom organismer på forskjellige trofiske nivåer er gjennom energipyramider. De representerer eog produktiviteten til næringskjeder og er konstruert ved å telle mengden energi (kcal) akkumulert per overflateenhet per tidsenhet og brukt av organismer på hvert trofisk nivå. Solenergien som planten mottar, brukes bare delvis i prosessen med fotosyntese. Energien festet i karbohydrater representerer bruttoproduksjonen av økosystemet Karbohydrater brukes til å bygge protoplasma og plantevekst. En del av energien deres brukes på å puste. Andre-ordens forbrukere (rovdyr) ødelegger ikke hele biomassen til ofrene sine. Dessuten, av mengden de ødelegger, brukes bare en del til å lage biomasse av deres eget trofiske nivå. Resten brukes hovedsakelig på pusteenergi og skilles ut med ekskrementer og ekskrementer. I 1942 formulerte R. Lindeman først loven om energipyramiden, som i lærebøker ofte kalles "loven om 10 %". I henhold til denne loven beveger seg i gjennomsnitt ikke mer enn 10% av energien fra ett trofisk nivå i den økologiske pyramiden til et annet nivå. Bare 10-20% av den opprinnelige energien overføres til påfølgende heterotrofer. Ved å bruke energipyramidens lov er det lett å beregne at mengden energi som når tertiære rovdyr (trofisk nivå V) er omtrent 0,0001 av energien som absorberes av produsenter. Det følger at overføring av energi fra ett nivå til et annet skjer med svært lav effektivitet. Dette forklarer det begrensede antallet ledd i næringskjeden, uavhengig av en bestemt biocenose. E. Odum (1959) i en ekstremt forenklet næringskjede - alfalfa -> kalv - "barnet vurderte transformasjonen av energi, illustrerte størrelsen på tapene. La oss si, resonnerer han, det er alfalfa-avlinger på et område med 4 hektar Kalver spiser på dette feltet (det antas at de bare spiser alfalfa), og en 12 år gammel gutt spiser utelukkende kalvekjøtt. Resultatene av beregninger, presentert i form av tre pyramider: tall, biomasse og energi, indikerer at alfalfa bruker bare 0,24 % av all solenergi som faller på marken; kalven absorberer 8 % av dette produktet og kun 0,7 % av kalvens biomasse sikrer utviklingen til barnet i løpet av året. E. Odum viste dermed at bare en milliondel av den innfallende solenergien omdannes til rovdyrbiomasse, i dette tilfellet bidrar det til en økning i barnets vekt, og resten går tapt og forsvinner i en nedbrutt form i miljøet. Eksemplet ovenfor illustrerer tydelig den svært lave økologiske effektiviteten til økosystemene og den lave effektiviteten til transformasjon i næringskjeder. Vi kan oppgi følgende: hvis 1000 kcal (dag m 2) spilt inn av produsenter, deretter 10 kcal (dag m 2) går inn i biomassen til planteetere og bare 1 kcal (dag m 2) - inn i biomassen til rovdyr. Siden en viss mengde av et stoff kan brukes av hver biocenose gjentatte ganger, og en del energi én gang, er det mer hensiktsmessig å si at det skjer en kaskadeoverføring av energi i økosystemet. Forbrukere fungerer som et administrerende og stabiliserende ledd i økosystemet. Forbrukere genererer et spekter av mangfold i folketellingen, og forhindrer dominerende monopol. Regelen om forbrukernes kontrollerende verdi kan med rette betraktes som ganske grunnleggende. I følge kybernetiske synspunkter bør kontrollsystemet være mer komplekst i struktur enn det kontrollerte, da blir årsaken til mangfoldet av typer instrumenter tydelig. Forbrukernes kontrollerende betydning har også et energisk grunnlag. Strømmen av energi gjennom et eller annet trofisk nivå kan ikke absolutt bestemmes av tilgjengeligheten av mat i det underliggende trofiske nivået. Som kjent er det alltid en tilstrekkelig "reserve" igjen, siden fullstendig ødeleggelse av mat vil føre til forbrukernes død. Disse generelle mønstrene observeres innenfor rammen av befolkningsprosesser, samfunn, nivåer av den økologiske pyramiden og biocenoser som helhet. 3 Dynamiske prosesser som sikrer integritet og bærekraft i økosystemer Sykliske endringer i samfunn gjenspeiler den daglige, sesongmessige og langsiktige periodisiteten til ytre forhold og manifestasjonen av endogene rytmer til organismer. Den daglige dynamikken til økosystemene er hovedsakelig assosiert med rytmen til naturfenomener og er strengt periodisk i naturen. Vi har allerede vurdert at i hver biocenose er det grupper av organismer hvis livsaktivitet skjer på forskjellige tider av dagen. Noen er aktive på dagtid, andre om natten. Derfor forekommer periodiske endringer i sammensetningen og forholdet mellom individuelle typer biocenose av et bestemt økosystem, ettersom individuelle organismer er slått av fra det i en viss tid. Den daglige dynamikken i biocenosen leveres av både dyr og planter. Som kjent endres intensiteten og naturen til fysiologiske prosesser i planter i løpet av dagen - fotosyntese skjer ikke om natten, ofte blomster i planter åpner bare om natten og blir bestøvet av nattaktive dyr, andre er tilpasset pollinering om dagen. Daglig dynamikk i biocenoser er som regel mer uttalt, jo større forskjellen er i temperatur, fuktighet og andre miljøfaktorer mellom dag og natt. Mer signifikante avvik i biocenoser observeres med sesongmessig dynamikk. Dette skyldes de biologiske syklusene til organismer, som avhenger av den sesongmessige syklisiteten til naturfenomener. Årstidsskiftet har således en betydelig innvirkning på livsaktiviteten til dyr og planter (dvale, vintersøvn, diapause og migrasjon hos dyr; perioder med blomstring, fruktsetting, aktiv vekst, bladfall og vinterdvale hos planter). Den lagdelte strukturen til biocenosen er ofte gjenstand for sesongvariasjoner. Individuelle lag av planter kan forsvinne helt i de aktuelle årstidene, for eksempel et urteaktig lag som består av ettårige. Varigheten av biologiske årstider varierer på forskjellige breddegrader. I denne forbindelse er den sesongmessige dynamikken til biocenoser i de arktiske, tempererte og tropiske sonene forskjellig. Det kommer tydeligst til uttrykk i tempererte klimaøkosystemer og nordlige breddegrader. Langsiktig variasjon er normal i livet til enhver biocenose. Dermed svinger mengden nedbør som faller i Barabinsk-skogsteppen kraftig fra år til år; en rekke tørre år veksler med langsiktige perioder med rikelig nedbør. Dette har en betydelig innvirkning på planter og dyr. I dette tilfellet skjer utviklingen av økologiske nisjer - funksjonell avgrensning i det nye settet eller dets tillegg med lite mangfold. Langsiktige endringer i sammensetningen av biocenoser gjentas også i forbindelse med periodiske endringer i atmosfærens generelle sirkulasjon, som igjen er forårsaket av en økning eller reduksjon i solaktiviteten. I prosessen med daglig og sesongmessig dynamikk blir integriteten til biocenoser vanligvis ikke krenket. Biocenosen opplever kun periodiske svingninger i kvalitative og kvantitative egenskaper. Progressive endringer i økosystemet fører til slutt til erstatning av en biocenose med en annen, med et annet sett med dominerende arter. Årsakene til slike endringer kan være faktorer utenfor biocenosen som virker lenge i én retning, for eksempel økende forurensning av vannforekomster, økende uttørking av sumpjord som følge av gjenvinning, økt beitebruk mv. Disse endringene fra en biocenose til en annen kalles eksogenetiske. I tilfellet når den økende innflytelsen av en faktor fører til en gradvis forenkling av strukturen til biocenose, uttømming av sammensetningen og en reduksjon i produktivitet, kalles slike skift digressive eller digressive. Endogenetiske endringer oppstår som et resultat av prosesser som skjer innenfor selve biocenosen. Den suksessive erstatningen av en biocenose med en annen kalles økologisk suksesjon (fra lat. - suksesjon - sekvens, endring). Suksesjon er en prosess med selvutvikling av økosystemer. Suksesjon er basert på ufullstendigheten av den biologiske syklusen i en gitt biocenose. Det er kjent at levende organismer, som et resultat av deres vitale aktivitet, endrer miljøet rundt dem, fjerner noen stoffer fra det og metter det med metabolske produkter. Når populasjoner eksisterer i relativt lang tid, endrer de miljøet i en ugunstig retning og finner seg som et resultat fortrengt av populasjoner av andre arter, for hvilke de resulterende miljøtransformasjonene viser seg å være økologisk gunstige. I en biocenose skjer altså en endring av dominerende arter. Regelen (prinsippet) om økologisk duplisering er tydelig synlig her. Den langsiktige eksistensen av en biocenose er bare mulig hvis endringer i miljøet forårsaket av aktiviteten til noen levende organismer er gunstige for andre med motsatte krav. Basert på konkurrerende interaksjoner mellom arter under suksesjon, dannes det gradvis mer stabile kombinasjoner som tilsvarer spesifikke abiotiske miljøforhold. Et eksempel på suksess som fører til erstatning av ett samfunn med et annet er gjengroing av en liten innsjø etterfulgt av utseendet til en sump og deretter en skog i stedet for. Først dannes det et flytende teppe langs kantene av innsjøen - et flytende teppe av sedges, moser og andre planter. Innsjøen er stadig fylt med døde planterester - torv. Det dannes en sump, gradvis gjengrodd med skog. En påfølgende rekke samfunn som gradvis og naturlig erstatter hverandre etter hverandre kalles en suksessserie. Etterfølger i naturen er ekstremt varierte i skala. De kan observeres i banker med kulturer, som er planktonsamfunn - ulike typer flytealger og deres forbrukere - hjuldyr, flagellater i pytter og dammer, sumper, enger, skoger, forlatte dyrkbare land, forvitrede bergarter, etc. Det er et hierarki i organisering av økosystemer Det manifesterer seg også i suksesjonsprosesser - større transformasjoner av biocenoser består av mindre. I stabile økosystemer med en regulert syklus av stoffer, skjer det også stadig lokale suksesjonsendringer, som støtter den komplekse interne strukturen til samfunn. Typer etterfølgende endringer. Det er to hovedtyper av suksesjonsendringer: 1 - med deltagelse av autotrofe og heterotrofe populasjoner; 2 - med deltakelse av bare heterotrofer. Etterfølgelse av den andre typen skjer bare under forhold der det skapes en foreløpig tilførsel eller konstant tilførsel av organiske forbindelser, på grunn av hvilken samfunnet eksisterer: i hauger eller hauger med gjødsel, i nedbrytende plantemateriale, i reservoarer forurenset med organiske stoffer, etc. Etterfølgende prosess. I følge F. Clements (1916) består suksesjonsprosessen av følgende stadier: Suksesjon med endring av vegetasjon kan være primær og sekundær. Primær suksesjon er prosessen med utvikling og endring av økosystemer i tidligere ubebodde områder, som begynner med deres kolonisering. Et klassisk eksempel er den konstante begroing av nakne steiner med den eventuelle utviklingen av skoger på dem. Således, i de primære rekkefølgene som forekommer på bergartene i Uralfjellene, skilles følgende stadier: Sekundær suksesjon er restaurering av et økosystem som en gang eksisterte i et gitt område. Det begynner når de etablerte relasjonene til organismer i en etablert biocenose blir forstyrret som følge av et vulkanutbrudd, brann, felling, pløying osv. Skifter som fører til gjenoppretting av den tidligere biocenose kalles demutasjons i geobotanikk. Et eksempel er dynamikken i artsmangfoldet på øya Krakatoa etter fullstendig ødeleggelse av innfødt flora og fauna av en vulkan. Et annet eksempel er den sekundære rekkefølgen av den sibirske mørke barskogen (gran-cedar taiga) etter en ødeleggende skogbrann. I mer svidde områder dukker det opp pionermoser fra sporer som blåses av vinden: 3-5 år etter en brann er den mest rikelig med «brannmosen» Funaria hygrometrica, Geratodon purpureus, etc. Blant høyere planter kommer ildgress (Chamaenerion angustifolium) veldig raskt. koloniserer brente områder ), som allerede etter 2-3 måneder blomstrer rikelig i bålet, samt bakken rørgress (Calamagrostis epigeios) og andre arter. Ytterligere suksesjonsfaser observeres: rørenga viker for busker, deretter bjørke- eller ospeskog, furublandingsskog, furuskog, furu-sederskog, og til slutt, etter 250 år, skjer restaureringen av sedertre-granskogen . Sekundære suksesjoner skjer som regel raskere og lettere enn primære, siden i det forstyrrede habitatet er jordprofilen, frø, primordia og en del av den tidligere bestanden og tidligere forbindelser bevart. Demutering er ikke en repetisjon av noe stadium av primær suksesjon. Klimaks økosystem. Suksesjon ender med et stadium hvor alle arter i økosystemet, mens de reproduserer, opprettholder et relativt konstant antall og ingen ytterligere endring i sammensetningen skjer. Denne likevektstilstanden kalles klimaks, og økosystemet kalles klimaks. Under ulike abiotiske forhold dannes ulike klimaksøkosystemer. I et varmt og fuktig klima vil det være en tropisk regnskog, i et tørt og varmt klima vil det være en ørken. De viktigste biomene på jorden er klimaksøkosystemene i deres respektive geografiske områder. Endringer i økosystemet under suksesjon. Produktivitet og biomasse. Som allerede nevnt er suksesjon en naturlig, rettet prosess og endringene som skjer på ett eller annet stadium er karakteristiske for ethvert samfunn og er ikke avhengig av dets artssammensetning eller geografiske plassering. Det er fire hovedtyper av suksesjonsendringer: Det skal også bemerkes at skifte av arvefaser skjer i samsvar med visse regler. Hver fase forbereder miljøet for fremveksten av den neste. Loven om sekvensen for overgang av utviklingsfaser fungerer her; fasene av utviklingen av et naturlig system kan bare følge i en evolusjonært fast (historisk, miljøbestemt) rekkefølge, vanligvis fra relativt enkel til kompleks, som regel, uten tap av mellomstadier, men kanskje med deres veldig raske passasje eller evolusjonært faste fravær. Når et økosystem nærmer seg en tilstand av overgangsalder, i det, som i alle likevektssystemer, bremses alle utviklingsprosesser. Denne situasjonen gjenspeiles i loven om suksessiv nedbremsing: prosesser som skjer i modne likevektsøkosystemer som er i en stabil tilstand, har som regel en tendens til å avta. I dette tilfellet endres den gjenopprettende typen suksesjon til deres sekulære kurs, dvs. selvutvikling skjer innenfor grensene for overgangsalder eller nodal utvikling. Den empiriske loven om suksessiv nedbremsing er en konsekvens av regelen til G. Odum og R. Pinkerton, eller regelen om maksimal energi for å opprettholde et modent system: suksesjon fortsetter i retning av et fundamentalt skifte i energistrømmen mot å øke dens kvantitet, rettet mot å opprettholde systemet. Regelen til G. Odum og R. Pinkerton er på sin side basert på regelen om maksimal energi i biologiske systemer, formulert av A. Lotka. Dette spørsmålet ble senere godt utviklet av R. Margalef, Y. Odum og er kjent som bevis på prinsippet om "null maksimum", eller minimal vekst i et modent økosystem: et økosystem i suksesjonsutvikling har en tendens til å danne den største biomassen med minst biologisk produktivitet. Lindeman (1942) beviste eksperimentelt at suksess er ledsaget av en økning i produktiviteten opp til klimakssamfunnet, der energikonvertering skjer mest effektivt. Data fra studier av rekkefølgen av eike- og eike-askeskoger viser at produktiviteten faktisk øker i de senere stadiene. Men under overgangen til et klimakssamfunn er det vanligvis en nedgang i den totale produktiviteten. Dermed er produktiviteten i gammel skog lavere enn i ungskog, som igjen kan være mindre produktiv enn de mer artsrike urtelagene som går foran dem. Tilsvarende nedgang i produktivitet er observert i enkelte vannsystemer. Det er flere grunner til dette. En av dem er at akkumulering av næringsstoffer i voksende skogbiomasse kan føre til en nedgang i syklingen deres. Nedgangen i den totale produktiviteten kan ganske enkelt være et resultat av redusert vitalitet hos individer ettersom gjennomsnittsalderen deres i samfunnet øker. Etter hvert som suksessen skrider frem, akkumuleres en økende andel av tilgjengelige næringsstoffer i biomassen i samfunnet, og følgelig reduseres innholdet i den abiotiske komponenten av økosystemet (i jord eller vann). Mengden av detritus som produseres øker også. De viktigste primærforbrukerne er ikke planteetere, men skadelige organismer. Tilsvarende endringer forekommer også i trofiske nettverk. Detritus blir hovedkilden til næringsstoffer. Under suksess øker lukketheten til biogeokjemiske sykluser av stoffer. Omtrent 10 år fra det øyeblikket restaureringen av vegetasjonsdekket begynner, synker åpenheten til syklusene fra 100 til 10%, og deretter reduseres den enda mer og når et minimum i klimaksfasen. Regelen om økende lukkethet av den biogeokjemiske syklusen av stoffer under suksess, det kan sies med all sikkerhet, brytes av den menneskeskapte transformasjonen av vegetasjon og naturlige økosystemer generelt. Dette fører utvilsomt til en lang rekke anomalier i biosfæren og dens inndelinger. En nedgang i artsmangfoldet i overgangsalderen betyr ikke dens lave økologiske betydning. Mangfoldet av arter former suksessen, dens retning, og sikrer at det virkelige rommet fylles med liv. Et utilstrekkelig antall arter som utgjør komplekset kunne ikke danne en rekkefølge, og gradvis, med ødeleggelsen av klimaksøkosystemer, ville fullstendig ørkenspredning av planeten skje. Verdien av mangfold er funksjonell både statisk og dynamisk. Det skal bemerkes at der mangfoldet av arter ikke er nok til å danne biosfæren, som tjener som grunnlag for det normale naturlige forløpet av suksesjonsprosessen, og miljøet i seg selv er kraftig forstyrret, når suksessen ikke klimaksfasen, men ender med et nodalsamfunn - et paraklimaks, et langsiktig eller kortsiktig avledet fellesskap. Jo dypere forstyrrelsen av miljøet i et bestemt rom er, jo tidligere fases rekkefølge slutter. Når en eller en gruppe arter går tapt som følge av deres ødeleggelse (menneskeskapt forsvinning av habitater, sjeldnere utryddelse), er oppnåelsen av overgangsalder ikke en fullstendig gjenoppretting av det naturlige miljøet. Faktisk er dette et nytt økosystem, fordi nye forbindelser har oppstått i det, mange gamle har gått tapt, og en annen "fraying" av arter har utviklet seg. Økosystemet kan ikke gå tilbake til sin gamle tilstand, siden en tapt art ikke kan gjenopprettes. Når en hvilken som helst abiotisk eller biotisk faktor endres, for eksempel med vedvarende avkjøling, eller introduksjon av en ny art, vil en art som er dårlig tilpasset nye forhold møte en av tre veier. Loven om evolusjonær-økologisk irreversibilitet sier at et økosystem som har mistet deler av sine elementer eller har blitt erstattet av et annet som et resultat av en ubalanse av økologiske komponenter, ikke kan gå tilbake til sin opprinnelige tilstand under suksess hvis, under endringene, evolusjonært (mikroevolusjonært). ) endringer har skjedd i økologiske elementer (bevart eller midlertidig tapt). I tilfellet når noen arter går tapt i mellomfaser av suksesjon, kan dette tapet funksjonelt kompenseres, men ikke fullstendig. Når mangfoldet avtar utover et kritisk nivå, blir suksessforløpet forvrengt, og faktisk kan et klimaks som er identisk med fortiden ikke oppnås. For å vurdere naturen til restaurerte økosystemer er loven om evolusjonær-økologisk irreversibilitet viktig. Med tap av grunnstoffer er dette faktisk helt nye økologiske naturformasjoner med nydannede mønstre og sammenhenger. Dermed betyr ikke overføringen i fortiden av en art som falt ut av økosystemet under dens reakklimatisering dens mekaniske retur. Dette er faktisk introduksjonen av en ny art i et fornyet økosystem Loven om evolusjonær-økologisk irreversibilitet understreker utviklingsretningen ikke bare på biosystemnivå, men også på alle andre hierarkiske nivåer i biotaen. 4 Biosfæren som et globalt økosystem som sikrer integriteten og bærekraften til økosystemene Biosfæren er et globalt økosystem. Som nevnt tidligere er biosfæren delt inn i geobiosfære, hydrobiosfære og aerobiosfære (fig. 2.4). Geobiosfæren har inndelinger i samsvar med de viktigste miljødannende faktorene: terrabiosfæren og lithobiosfæren - innenfor geobiosfæren, marinobiosfæren (oceanobiosfæren) og akvabiosfæren - innenfor hydrobiosfæren. Disse formasjonene kalles undersfærer. Den ledende miljødannende faktoren i deres dannelse er den fysiske fasen av livsmiljøet: luft-vann i aerobiosfæren, vann - ferskvann og saltvann i hydrobiosfæren, fast luft i terrabiosfæren og fast vann i litobiosfæren. I sin tur faller de alle i lag: aerobiosfæren - inn i tropobiosfæren og altobiosfæren; hydrobiosfære - inn i fotosfære, disfotosfære og afotosfære. De strukturdannende faktorene her, i tillegg til det fysiske miljøet, energi (lys og varme), spesielle forhold for dannelse og utvikling av liv - de evolusjonære retningene for penetrering av biota på land, i dets dyp, inn i rommene over jorden, havets avgrunn, er utvilsomt annerledes. Sammen med apobiosfæren, parabiosfæren og andre sub- og supra-biosfærelag utgjør de den såkalte «livets lagkake» og geosfæren (økosfæren) for dens eksistens innenfor megabiosfærens grenser. I systemisk forstand er de listede formasjonene store funksjonelle deler av praktisk talt universelle eller subplanetariske dimensjoner. Forskere mener at i biosfæren er det minst 8-9 nivåer av relativt uavhengige sykluser av stoffer innenfor sammenkoblingen av 7 hovedmateriale-energi-økologiske komponenter og den åttende - informasjonsmessig. Globale, regionale og lokale sykluser av stoffer er ikke lukket og "skjærer" seg delvis i økosystemhierarkiet. Denne material-energi, og delvis informasjonsmessige "koblingen" sikrer integriteten til økologiske supersystemer opp til biosfæren som helhet. Integritet og bærekraft til økosystemer. Biosfæren dannes i større grad ikke av ytre faktorer, men av indre mønstre. Den viktigste egenskapen til biosfæren er samspillet mellom levende og ikke-levende ting, noe som gjenspeiles i loven om biogen migrering av atomer av V. I. Vernadsky. Loven om biogen migrasjon av atomer gjør det mulig for menneskeheten å bevisst kontrollere biogeokjemiske prosesser både på jorden som helhet og i dens regioner. Mengden levende stoff i biosfæren er som kjent ikke gjenstand for merkbare endringer. Dette mønsteret ble formulert i form av loven om konstanthet av mengden levende materie av V.I. Vernadsky: Mengden levende stoff i biosfæren for en gitt geologisk periode er konstant. I praksis er denne loven en kvantitativ konsekvens av loven om intern dynamisk likevekt for det globale økosystemet – biosfæren. Siden levende materie, i samsvar med loven om biogen migrasjon av atomer, er en energimellomledd mellom Solen og Jorden, må mengden enten være konstant, eller dens energikarakteristikker må endres. Loven om fysisk og kjemisk enhet av levende materie (alt levende materiale på jorden er fysisk og kjemisk forent og utelukker betydelige endringer i sistnevnte egenskap. Kvantitativ stabilitet er derfor uunngåelig for planetens levende materie. Den er fullt ut karakteristisk for antall arter. Levende materie, som en akkumulator av solenergi, må samtidig reagere på både ytre (kosmiske) påvirkninger og indre endringer. En reduksjon eller økning i mengden levende stoff på ett sted i biosfæren bør føre til en prosess nøyaktig det motsatte på et annet sted, fordi de frigjorte næringsstoffene kan assimileres av resten av det levende stoffet eller mangel på dem vil bli observert. Her må vi ta hensyn til hastigheten på prosessen, som ved menneskeskapte forandringer er mye lavere enn direkte forstyrrelse av naturen av mennesket. I tillegg til konstansen og konstantheten til mengden levende materie, som gjenspeiles i loven om fysisk og kjemisk enhet av levende materie, er det i levende natur en konstant bevaring av den informasjonsmessige og somatiske strukturen, til tross for at den endres noe med evolusjonens gang. Denne egenskapen ble notert av Yu. Goldsmith (1981) og ble kalt loven om bevaring av strukturen til biosfæren - informativ og somatisk, eller den første loven om økodynamikk. For å bevare strukturen til biosfæren streber levende ting etter å oppnå en tilstand av modenhet eller økologisk balanse. Loven om ønsket om overgangsalder – den andre økodynamikkens lov av Yu. Goldsmith, gjelder for biosfæren og andre nivåer av økologiske systemer, selv om det er spesifikt – biosfæren er et mer lukket system enn dens underavdelinger. Enheten av det levende stoffet i biosfæren og homologien til strukturen til dens undersystemer fører til det faktum at de levende elementene i forskjellige geologiske aldre og opprinnelige geografiske opprinnelser som oppsto på den, er intrikat sammenvevd evolusjonært. Sammenvevingen av elementer av forskjellig rom-temporal genese på alle økologiske nivåer i biosfæren reflekterer regelen eller prinsippet om heterogenese av levende materie. Dette tillegget er ikke kaotisk, men er underlagt prinsippene om økologisk komplementaritet, økologisk konformitet (kongruens) og andre lover. Innenfor rammen av økodynamikken til Yu. Goldsmith er dette dens tredje lov - prinsippet om økologisk orden, eller økologisk gjensidighet, som indikerer en global eiendom på grunn av påvirkningen av helheten på dens deler, den omvendte påvirkningen av differensierte deler på utvikling av helheten osv., som totalt sett fører til bevaringsstabilitet av biosfæren som helhet. Gjensidig hjelp innenfor rammen av den økologiske orden, eller systemisk gjensidighet, bekreftes av loven om orden for å fylle rom og romlig-tidsmessig sikkerhet: fyllingen av rom innenfor et naturlig system, på grunn av samspillet mellom dets undersystemer, er ordnet i en slik måte som gjør at de homeostatiske egenskapene til systemet kan realiseres med minimale motsetninger mellom delene i det. Av denne loven følger det at den langsiktige eksistensen av ulykker som er "unødvendige" for naturen, inkludert menneskelige kreasjoner som er fremmede for den, er umulig. Reglene for den mutualistiske systemordenen i biosfæren inkluderer også prinsippet om systemkomplementaritet, som sier at undersystemene til ett naturlig system i deres utvikling gir en forutsetning for vellykket utvikling og selvregulering av andre delsystemer som inngår i samme system. Den fjerde økodynamiske loven av Yu. Goldsmith inkluderer loven om selvkontroll og selvregulering av levende ting: levende systemer og systemer under kontrollerende påvirkning av levende ting er i stand til selvkontroll og selvregulering i prosessen med deres tilpasning til endringer i omgivelsene. I biosfæren skjer selvkontroll og selvregulering under kaskade- og kjedeprosesser av generell interaksjon - under kampen for eksistensen av naturlig utvalg (i videste forstand av dette konseptet), tilpasning av systemer og delsystemer, bred samevolusjon , etc. Dessuten fører alle disse prosessene til positive resultater "fra synspunktet" av naturen - bevaring og utvikling av økosystemene i biosfæren og den som helhet. Forbindelseskoblingen mellom generaliseringer av strukturell og evolusjonær natur er regelen for automatisk vedlikehold av det globale habitatet: levende materie, i løpet av selvregulering og interaksjon med abiotiske faktorer, opprettholder autodynamisk et livsmiljø som er egnet for dets utvikling. Denne prosessen er begrenset av endringer i den kosmiske og globale økosfæreskalaen og forekommer i alle økosystemer og biosystemer på planeten, som en kaskade av selvregulering som når en global skala. Regelen om automatisk vedlikehold av det globale habitatet følger av de biogeokjemiske prinsippene til V.I. Vernadsky, reglene for å bevare arters habitater, relativ intern konsistens og fungerer som en konstant for tilstedeværelsen av konservative mekanismer i biosfæren og bekrefter samtidig regelen om systemdynamisk komplementaritet. Den kosmiske påvirkningen på biosfæren er bevist av loven om brytning av kosmiske påvirkninger: kosmiske faktorer, som har innvirkning på biosfæren og spesielt dens underavdelinger, er gjenstand for endring av planetens økosfære og derfor, når det gjelder styrke og tid , kan manifestasjoner bli svekket og forskjøvet eller til og med helt miste sin effekt. Generaliseringen her er viktig på grunn av det faktum at det ofte er en strøm av synkrone effekter av solaktivitet og andre kosmiske faktorer på jordens økosystemer og organismene som bor i den. Det skal bemerkes at mange prosesser på jorden og i dens biosfære, selv om de er underlagt påvirkning av rommet, antas sykluser av solaktivitet med intervaller på 1850, 600, 400, 178, 169, 88, 83, 33, 22, 16 , 11, 5 (11 ,1), 6,5 og 4,3 år, trenger ikke biosfæren selv og dens inndelinger nødvendigvis å reagere med samme syklisitet i alle tilfeller. De kosmiske påvirkningene av biosfæresystemet kan blokkeres helt eller delvis. Konklusjon Økosystemer er den grunnleggende funksjonelle enheten i økologi, siden de inkluderer organismer og det livløse miljøet - komponenter som gjensidig påvirker hverandres egenskaper og de nødvendige betingelsene for å opprettholde liv i den formen som finnes på jorden. Kombinasjonen av et spesifikt fysisk-kjemisk miljø (biotop) med et fellesskap av levende organismer (biocenose) danner et økosystem. Det er mikroøkosystemer, mesoøkosystemer og det globale økosystemet - biosfæren. Økosystemer er ikke spredt i uorden, tvert imot er de gruppert i ganske regelmessige soner, både horisontalt (i breddegrad) og vertikalt (i høyde). Den primære energikilden for økosystemer er solen. Solstråling som faller på jordoverflaten er 1,54 millioner EJ per år. Mesteparten av solenergien som når planetens overflate omdannes direkte til varme, varmende vann eller jord, som igjen varmer opp luften. Økosystemer eksisterer på grunn av ikke-forurensende og nesten evig solenergi, hvor mengden er relativt konstant og rikelig. Solenergi på jorden forårsaker to sykluser av stoffer: store, eller geologiske, og små, biologiske (biotiske). Begge syklusene er gjensidig forbundet og representerer så å si en enkelt prosess. Eksistensen av biogeokjemiske sykluser, eller biogeokjemiske sykluser, skaper muligheten for selvregulering (homeostase) av systemet, noe som gir økosystemet stabilitet: en forbløffende konstanthet av prosentandelen av ulike elementer. Det er ganske mange sykluser av stoffer. Den mest betydningsfulle syklusen på jorden når det gjelder overførte masser og energiforbruk er den planetariske hydrologiske syklusen eller vannsyklusen. Den biotiske (biologiske) syklusen refererer til sirkulasjonen av stoffer mellom jord, planter, dyr og mikroorganismer. Av alle biogeokjemiske sykluser er karbonsyklusen uten tvil den mest intense. Det meste av oksygenet som ble produsert under geologiske epoker forble ikke i atmosfæren, men ble fiksert av litosfæren i form av karbonater, sulfater, jernoksider osv. Denne massen er 590x1014 tonn mot 39x1014 tonn oksygen, som sirkulerer i biosfæren i form av gass eller sulfater oppløst i kontinentale og oceaniske farvann. Nitrogen er et essensielt biogent element, siden det er en del av proteiner og nukleinsyrer. Nitrogensyklusen er en av de mest komplekse, siden den inkluderer både gass- og mineralfaser, og samtidig de mest ideelle syklusene. Fosforsyklusen i biosfæren er assosiert med metabolske prosesser i planter og dyr. Dette viktige og nødvendige elementet i protoplasma, inneholdt i landplanter og alger 0,01-0,1%, dyr fra 0,1% til flere prosent, sirkulerer, og blir gradvis fra organiske forbindelser til fosfater, som igjen kan brukes av planter. Det er mange gassformige svovelforbindelser, som hydrogensulfid H2S og svoveldioksid SO2. Å opprettholde den vitale aktiviteten til organismer og sirkulasjonen av materie i økosystemene, det vil si eksistensen av økosystemer, avhenger av den konstante strømmen av energi som er nødvendig for alle organismer for deres vitale funksjoner og selvreproduksjon. I motsetning til stoffer som kontinuerlig sirkulerer gjennom ulike blokker av økosystemet, som alltid kan gjenbrukes og gå inn i syklusen, kan energi brukes én gang, det vil si at det er en lineær strøm av energi gjennom økosystemet. Innenfor et økosystem skapes energiholdige stoffer av autotrofe organismer og tjener som mat for heterotrofer. Matforbindelser er mekanismer for å overføre energi fra en organisme til en annen. Innenfor hvert økosystem har næringsnettene en veldefinert struktur, som er preget av arten og antallet organismer representert på hvert nivå i de ulike næringskjedene. Dannelsen av økosystemer er en dynamisk prosess. I økosystemer skjer det konstant endringer i tilstanden og vital aktivitet til medlemmene deres og i forholdet mellom populasjoner. De forskjellige endringene som skjer i ethvert samfunn faller inn i to hovedtyper: sykliske og progressive. Suksesjon er en naturlig, rettet prosess og endringene som skjer på ett eller annet stadium er karakteristiske for ethvert samfunn og er ikke avhengig av dets artssammensetning eller geografiske plassering. Det globale økosystemet er biosfæren. Som nevnt tidligere er biosfæren delt inn i geobiosfære, hydrobiosfære og aerobiosfære. Biosfæren dannes i større grad ikke av ytre faktorer, men av indre mønstre. Den viktigste egenskapen til biosfæren er samspillet mellom levende og ikke-levende ting, noe som gjenspeiles i loven om biogen migrering av atomer av V.I. Vernadsky. Økosystemene som eksisterer på jorden er forskjellige. Det er mikroøkosystemer (for eksempel stammen til et råtnende tre), mesoøkosystemer (skog, dam, etc.), makroøkosystemer (kontinent, hav, etc.) og den globale biosfæren. Store terrestriske økosystemer kalles biomer. Hvert biom inneholder en rekke mindre, sammenkoblede økosystemer. Det er flere klassifiseringer av økosystemer: eviggrønn tropisk regnskog; ørken: gresskledd og buskete; tropisk gress og savanne; temperert steppe; temperert edelløvskog: boreale barskoger. Tundra: arktisk og alpint. Typer ferskvannsøkosystemer: bånd (stille vann): innsjøer, dammer, etc.; lotic (rennende vann): elver, bekker, etc.; våtmarker: sumper og sumpete skoger; Typer marine økosystemer: åpent hav (pelagisk); vann på kontinentalsokkelen (kystfarvann); oppvekstområder (fruktbare områder med produktivt fiskeri); elvemunninger (kystbukter, sund, elvemunninger, saltmyrer, etc.). Hvert økosystem har to hovedkomponenter: organismer og faktorer fra deres ikke-levende miljø. Helheten av organismer (planter, dyr, mikrober) kalles biotaen til et økosystem. Måtene for interaksjon mellom ulike kategorier av organismer er dens biotiske struktur. Solenergi på jorden forårsaker to sykluser av stoffer: en stor, eller geologisk, tydeligst manifestert i vannets syklus og atmosfærisk sirkulasjon, og en liten, biologisk (biotisk), som utvikler seg på grunnlag av en stor og består av en kontinuerlig, syklisk, men ujevn i tid og rom, og ledsaget av mer eller mindre betydelige tap i den naturlige omfordelingen av materie, energi og informasjon innenfor økologiske systemer på ulike organisasjonsnivåer. Begge syklusene er gjensidig forbundet og representerer så å si en enkelt prosess. Samspillet mellom abiotiske faktorer og levende organismer i økosystemet er ledsaget av en kontinuerlig sirkulasjon av materie mellom biotopen og biocenosen i form av alternerende organiske og mineralske forbindelser. Utvekslingen av kjemiske elementer mellom levende organismer og det uorganiske miljøet, hvis ulike stadier forekommer innenfor et økosystem, kalles den biogeokjemiske syklusen, eller biogeokjemisk syklus. Eksistensen av slike sykluser skaper muligheten for selvregulering (homeostase) av systemet, noe som gir økosystemet stabilitet: en utrolig konstanthet av prosentandelen av forskjellige elementer. Prinsippet om økosystemenes funksjon gjelder her: anskaffelse av ressurser og deponering av avfall skjer innenfor rammen av syklusen til alle elementer. Liste over brukt litteratur 1.Bigon M.I. Økologi, individer, befolkninger og samfunn. M.: 1989. - 290 s. 2.Beysenova A.S., Shildebaev Zh.B., Sautbaeva G.Z. Økologi. Almaty: "Gylym, 2001. - 201 s. 3.Akimova T.A., Khaskin V.V. Økologi. M.: UNITY Publishing House, 1998. - 233 s. .Gorelov A.A. Økologi. Forelesningskurs. M.: "Senter" 1997. - 280 s. .Stadnitsky G.V., Rodionov A.I. Økologi. M.: UNITY Publishing House, 1996. - 272 s. .Sadanov A.K., Svanbaeva Z.S., Økologi. Almaty: "Agrouniversity", 1999. - 197 s. .Shilov I.A. Økologi. M.: Videregående skole, 2000. - 348 s. .Reimers N.F. Økologi (Teori, lover, regler, prinsipper og hypoteser). M.: "Ungt Russland", 1994. - 260 s. .Tsvetkova L.I., Alekseev M.I., Usenov B.P. og andre Økologi. Lærebok for tekniske universiteter. M.; ASV, St. Petersburg: Khimizdat, 1999, 185 s. .Girusov E.V. Økologi og økonomi for miljøledelse. M.: Lov og lov, ENHET. 1998 - 232 s. .Vronsky V.A. Anvendt økologi: Lærebok. Fordel. Rostov ved Don: 1995. - 197 s. .Budyko M.I. Global økologi. M.: Mysl, 1977.-248 s. .Alekseenko V.A. Miljøgeokjemi. Lærebok. M.: Logos, 2000 - 410 s. .Petrov K.M. Generell økologi av S-P. Opplæringen. "Kjemi", 1997. - 218 s. .Andersen J.M. Økologi og miljøvitenskap. Biosfære, økosystem, menneske. L., 1985. 376 s. .Girenok F.I. Økologi. Sivilisasjon. Noosphere.M.: 191987. - 281 s. .På jakt etter balanse: økologi i systemet med sosialpolitiske prioriteringer M.: 1992. - 427 s. .Historie om samspill mellom samfunn og natur: Faktorer og konsept. M.: ANSSSR 1990. .Stepanovskikh A.S. Anvendt økologi: Miljøvern. Lærebok for universiteter. M.: ENHET DANA. 2003. 751 s. Lignende arbeider til - Syklus av stoffer og energistrømmer i økosystemer
Begrepene "flyt av materie" og "strøm av energi" bør være klart definert. Materiestrømmen er bevegelsen i form av kjemiske elementer og deres forbindelser fra produsenter til nedbrytere (gjennom eller uten forbrukere). Energiflyt er overføring av energi i form av kjemiske bindinger av organiske forbindelser (mat) langs næringskjeder fra et trofisk nivå til et annet (høyere).
Det skal bemerkes at, i motsetning til stoffer som hele tiden sirkulerer gjennom forskjellige blokker i økosystemet og alltid kan gå inn i syklusen igjen, kan den innkommende energien bare brukes én gang.
Som et universelt naturfenomen er enveisstrømmen av energi bestemt av termodynamikkens lover. I følge den første loven kan energi bevege seg fra en form (lysenergien) til en annen (den potensielle energien til mat), men den blir aldri skapt igjen og forsvinner ikke sporløst.
Termodynamikkens andre lov sier at det ikke kan være en enkelt prosess forbundet med transformasjonen av energi uten å miste noe av den. Av denne grunn kan det ikke være noen transformasjon med 100 % effektivitet, for eksempel av mat til stoffet som kroppen til organismen består av.
Dermed er funksjonen til alle økosystemer bestemt av en konstant strøm av energi, som er nødvendig for at alle organismer skal opprettholde sin eksistens og selvreproduksjon.
Det er også konkurranseforhold i økosystemer. I dette aspektet er loven om energimaksimering (G. Odum - E. Odum) av stor interesse: i konkurranse med andre økosystemer overlever den som best bidrar til energiforsyningen og bruker sin maksimale mengde på den mest effektive måten. (konserverer). I henhold til loven, for dette formålet: 1) skaper akkumulatorer (lagre) av høykvalitets energi (for eksempel fettreserver); 2) bruker en viss mengde akkumulert energi for å sikre tilførsel av ny energi; 3) sikrer sirkulasjonen av ulike stoffer; 4) skaper reguleringsmekanismer som støtter systemets stabilitet og dets evne til å tilpasse seg endrede forhold; 5) etablerer utveksling med andre systemer som er nødvendige for å møte behovet for spesielle energityper.
Det er nødvendig å understreke en viktig omstendighet: loven om energimaksimering er også gyldig i forhold til informasjon, derfor kan den (ifølge N. F. Reimers) også betraktes som loven om energi og informasjonsmaksimering: systemet som mest bidrar til mottak, produksjon og effektiv bruk av energi og informasjon.
La oss merke oss at maksimal tilførsel av et stoff som sådan ennå ikke garanterer suksessen til systemet i en konkurrerende gruppe av andre lignende systemer.
Det ble tidligere bemerket at sterke matrelasjoner, eller en næringskjede, oppstår og etableres mellom organismene i biocenosen. Sistnevnte består av tre hovedledd: produsenter, forbrukere og nedbrytere.
Næringskjeder som begynner med fotosyntetiske organismer kalles beitekjeder (eller beitekjeder), og kjeder som begynner med dødt plantemateriale, kadaver og dyreekskrement kalles detritelle kjeder.
Plasseringen av hvert ledd i næringskjeden kalles et trofisk nivå; den er preget av forskjellige intensiteter av flyten av stoffer og energi. Det første trofiske nivået er alltid produsenter; den andre - planteetende forbrukere; den tredje - kjøttetere, lever av planteetende former; fjerde nivå - konsumere andre rovdyr, etc.
Det er forbrukere av første, andre, tredje og fjerde ordre, som okkuperer forskjellige nivåer i næringskjeden (fig. 9).
Ris. 9.
Det er åpenbart at matspesialiseringen til forbrukerne spiller en stor rolle i dette. Arter med et bredt spekter av ernæring kan inkluderes i næringskjeder på forskjellige trofiske nivåer. Kostholdet til for eksempel en person inkluderer både plantemat og kjøtt fra planteetere og rovdyr. Derfor fungerer den i forskjellige næringskjeder som forbruker av I-, II- eller III-bestillingene.
Siden energi går tapt når energi overføres fra et nivå til et annet, kan ikke kraftkjeden være lang: den består vanligvis av 4...6 ledd (tabell 1).
1. Typiske diagrammer over næringskjeder (ifølge V. M. Ivonin, 1996)
Imidlertid finnes slike kjeder i sin rene form vanligvis ikke i naturen, siden samme art kan være tilstede samtidig i forskjellige ledd. Dette skyldes det faktum at det er få monofager i naturen; oligofager og polyfager er mye mer vanlige. For eksempel er rovdyr som lever av ulike planteetere og rovdyr ledd i mange kjeder. Som et resultat, i hver biocenose dannes komplekser av næringskjeder evolusjonært, som representerer en enkelt helhet. Slik skapes kraftnettverk, som er svært komplekse.
Dermed kan vi konkludere med at næringskjeden er hovedkanalen for energioverføring i samfunnet (mellom planter - produsenter, dyr - forbrukere og mikroorganismer - nedbrytere) (Fig. 10) Allerede i diagrammet er det tydelig at ideen om næringskjeder og trofiske nivåer er snarere en abstraksjon. En lineær kjede med klart adskilte nivåer kan lages i laboratoriet. Men i naturen er det virkelig trofiske nettverk der mange populasjoner tilhører flere trofiske nivåer samtidig. Den samme organismen forbruker både dyr og planter; rovdyret kan livnære seg på forbrukere av første og andre orden; mange dyr spiser både levende og døde planter.
På grunn av kompleksiteten til trofiske relasjoner har tapet av én art ofte nesten ingen effekt på samfunnet. Andre "brukere" begynner å konsumere maten til den utdødde arten,

Ris. 10.
artene som lever av den finner nye matkilder: generelt opprettholdes balansen i samfunnet.
Energien som absorberes av produsenter flyter gjennom næringskjeder og forbrukes gradvis. På slutten av en næringskjede er energimengden alltid mindre enn i begynnelsen. Under fotosyntesen binder planter i gjennomsnitt bare rundt 1 % av solenergien som treffer dem. Et dyr som har spist en plante fordøyer ikke en del av maten og skiller den ut i form av ekskrementer. Vanligvis fordøyes 20...60% av plantematen; Den absorberte energien brukes til å opprettholde dyrets liv. Funksjonen til celler og organer er ledsaget av frigjøring av varme, det vil si at en betydelig andel av matenergien raskt forsvinner i miljøet. En relativt liten del av maten brukes til å bygge nytt vev og lage fettreserver. Deretter mottar et rovdyr som har spist en planteeter og representerer det tredje trofiske nivået bare den energien fra energien akkumulert av planten som holdes tilbake i byttets kropp (andre nivå) i form av en økning i biomasse.
Det er kjent at på hvert trinn under overføringen av materie og energi i næringskjeden, går omtrent 90 % av energien tapt og bare omtrent en tiendedel av den går til neste forbruker, dvs. overføringen av energi i matforbindelsene av organismer adlyder «ti prosent-regelen» (Lindemanns prinsipp ). For eksempel er mengden energi som når tertiære rovdyr (femte trofiske nivå) bare omtrent 10 -4 av energien som absorberes av produsenter. Dette forklarer det begrensede antallet (5...6) ledd (nivåer) i næringskjeden, uavhengig av kompleksiteten i artssammensetningen til biocenosen.

Ris. elleve.
Med tanke på strømmen av energi i økosystemer, er det også lett å forstå hvorfor biomassen avtar når det trofiske nivået øker. Her manifesteres det tredje grunnleggende prinsippet om økosystemenes funksjon: jo større biomasse en befolkning har, desto lavere skal det trofiske nivået være, eller på annen måte: på slutten av lange næringskjeder kan det ikke være en stor biomasse.
De tre grunnleggende prinsippene for økosystemfunksjoner som er oppført ovenfor - syklusen av næringsstoffer, strømmen av solenergi og nedgangen i biomasse med økende trofisk nivå - kan presenteres i form av et generelt diagram (fig. 11). Hvis vi ordner organismer i henhold til deres ernæringsmessige forhold, og indikerer for hver av dem "inngang" og "utgang" av energi og næringsstoffer, blir det åpenbart at næringsstoffer kontinuerlig resirkuleres i økosystemet, og strømmen av energi passerer gjennom det.
Å opprettholde den vitale aktiviteten til organismer og sirkulasjonen av materie i økosystemene, det vil si eksistensen av økosystemer, avhenger av den konstante strømmen av energi som er nødvendig for alle organismer for deres vitale funksjoner og selvreproduksjon.
I motsetning til stoffer som kontinuerlig sirkulerer gjennom forskjellige blokker av økosystemet, som alltid kan gjenbrukes og gå inn i syklusen, kan energi bare brukes én gang, det vil si at det er en lineær strøm av energi gjennom økosystemet.
Enveistilstrømningen av energi som et universelt naturfenomen oppstår som et resultat av termodynamikkens lover. Første lov sier at energi kan omdannes fra en form (som lys) til en annen (som den potensielle energien til mat), men kan ikke skapes eller ødelegges. Andre lov sier at det ikke kan være en enkelt prosess knyttet til transformasjonen av energi uten å miste noe av den. En viss mengde energi i slike transformasjoner spres til utilgjengelig termisk energi og går derfor tapt. Derfor kan det ikke være transformasjoner av for eksempel matstoffer til stoffet som utgjør kroppen til organismen, som skjer med 100 prosent effektivitet.
Dermed er levende organismer energiomdannere. Og hver gang energi omdannes, går en del av den tapt i form av varme. Til syvende og sist blir all energien som kommer inn i den biotiske syklusen til et økosystem bort som varme. Levende organismer bruker faktisk ikke varme som energikilde for å utføre arbeid – de bruker lys og kjemisk energi.
Økologiske pyramider. Innenfor hvert økosystem har næringsnettene en veldefinert struktur, som er preget av arten og antallet organismer representert på hvert nivå i de ulike næringskjedene. For å studere forholdet mellom organismer i et økosystem og for å skildre dem grafisk, bruker de vanligvis økologiske pyramider i stedet for næringsnettdiagrammer. Økologiske pyramider uttrykker den trofiske strukturen til et økosystem i geometrisk form. De er konstruert i form av rektangler med samme bredde, men lengden på rektanglene må være proporsjonal med verdien av objektet som måles. Herfra kan du få pyramider av tall, biomasse og energi.
Økologiske pyramider gjenspeiler de grunnleggende egenskapene til enhver biocenose når de viser dens trofiske struktur:
Høyden deres er proporsjonal med lengden på den aktuelle næringskjeden, dvs. antall trofiske nivåer den inneholder;
Formen deres gjenspeiler mer eller mindre effektiviteten til energitransformasjoner under overgangen fra et nivå til et annet.
Pyramider av tall. De representerer den enkleste tilnærmingen til studiet av den trofiske strukturen til et økosystem. I dette tilfellet telles først antallet organismer i et gitt territorium, gruppert etter trofiske nivåer og presentert i form av et rektangel, hvis lengde (eller areal) er proporsjonal med antall organismer som lever i et gitt område ( eller i et gitt volum, hvis det er et akvatisk økosystem). Det er etablert en grunnleggende regel om at i ethvert miljø er det flere planter enn dyr, flere planteetere enn rovdyr, flere insekter enn fugler, etc.
Befolkningspyramider gjenspeiler tettheten av organismer på hvert trofisk nivå. Det er stort mangfold i byggingen av ulike befolkningspyramider. Ofte er de opp ned.
For eksempel er det i en skog betydelig færre trær (primærprodusenter) enn insekter (planteetere).
Biomassepyramide. Gjenspeiler matforholdet i økosystemet mer fullstendig, siden det tar hensyn til den totale massen av organismer (biomasse) hvert trofisk nivå. Rektanglene i biomassepyramidene representerer massen av organismer på hvert trofiske nivå per enhetsareal eller volum. Formen på biomassepyramiden er ofte lik formen på befolkningspyramiden. En nedgang i biomasse på hvert påfølgende trofisk nivå er karakteristisk.
Pyramider av biomasse, så vel som tall, kan ikke bare være rette, men også inverterte. Inverterte pyramider av biomasse er karakteristiske for akvatiske økosystemer, der primærprodusenter, som planteplanktoniske alger, deler seg veldig raskt, og forbrukerne deres - zooplanktoniske krepsdyr - er mye større, men har en lang reproduksjonssyklus. Spesielt gjelder dette ferskvannsmiljøer, der primærproduktiviteten leveres av mikroskopiske organismer hvis metabolske hastigheter økes, dvs. biomassen er lav, produktiviteten er høy.
Pyramide av energi. Den mest grunnleggende måten å vise forbindelser mellom organismer på forskjellige trofiske nivåer er gjennom energipyramider. De representerer eog produktiviteten til næringskjeder og er konstruert ved å telle mengden energi (kcal) akkumulert per overflateenhet per tidsenhet og brukt av organismer på hvert trofisk nivå. Dermed er det relativt enkelt å bestemme mengden energi som er lagret i biomasse, men det er vanskeligere å anslå den totale mengden energi som absorberes på hvert trofisk nivå. Etter å ha konstruert en graf (fig. 12.28), kan vi slå fast at destruktorer, hvis betydning synes liten i biomassepyramiden, og omvendt i befolkningspyramiden; motta en betydelig del av energien som passerer gjennom økosystemet. Dessuten forblir bare en del av all denne energien i organismer på hvert trofiske nivå i økosystemet og lagres i biomasse; resten brukes til å tilfredsstille de metabolske behovene til levende vesener: opprettholdelse av eksistens, vekst, reproduksjon. Dyr bruker også en betydelig mengde energi på muskelarbeid.
R. Lindeman ble først formulert i 1942 lov om energipyramide, som i lærebøker ofte kalles "loven på 10%". I henhold til denne loven, fra en trofisk nivå av den økologiske pyramiden I gjennomsnitt går ikke mer enn 10 % av energien til et annet nivå.
Forbrukere fungerer som et administrerende og stabiliserende ledd i økosystemet. Forbrukere genererer et spekter av mangfold i folketellingen, og forhindrer dominerende monopol. Regel for kontroll verdi av forbrukere kan med rette betraktes som ganske grunnleggende. I følge kybernetiske synspunkter bør kontrollsystemet være mer komplekst i struktur enn det kontrollerte, da blir årsaken til mangfoldet av forbrukertyper tydelig. Forbrukernes kontrollerende betydning har også et energisk grunnlag. Strømmen av energi gjennom et eller annet trofisk nivå kan ikke absolutt bestemmes av tilgjengeligheten av mat i det underliggende trofiske nivået. Som kjent er det alltid en tilstrekkelig "reserve" igjen, siden fullstendig ødeleggelse av mat vil føre til forbrukernes død. Disse generelle mønstrene observeres innenfor rammen av befolkningsprosesser, samfunn, nivåer av den økologiske pyramiden og biocenoser som helhet.
For å bruke forhåndsvisninger av presentasjoner, opprett en Google-konto og logg på den: https://accounts.google.com
Lysbildetekster:
Syklus av materie og energi i naturen
Stoffers syklus er de gjentatte prosessene med transformasjon og bevegelse av materie i naturen, som er mer eller mindre sykliske. Alle stoffer på planeten vår er i ferd med å sirkulere. Det er to hovedsykluser i naturen: Stor (geologisk) Liten (biogeokjemisk)
Den store syklusen av stoffer Den store syklusen varer i millioner av år og er forårsaket av samspillet mellom solenergi og jordens dype energi. Assosiert med geologiske prosesser, dannelse og ødeleggelse av bergarter og påfølgende bevegelse av ødeleggelsesprodukter.
Liten syklus av stoffer Den lille syklusen (biogeokjemisk) oppstår i biosfæren, på nivået av biocenose. Dens essens er dannelsen av levende materiale fra uorganiske forbindelser under prosessen med fotosyntese og transformasjonen av organisk materiale under nedbrytning tilbake til uorganiske forbindelser. Biogeokjemiske sykluser – Vernadsky V.I.
Vannsyklus Tr avrenning inf Vannfordampning Dampkondensasjon Nedbør avrenning Transpirasjonsinfiltrasjon
Transpirasjon er prosessen med vann som beveger seg gjennom en plante og dets fordampning gjennom plantens ytre organer, som blader, stilker og blomster. Vann er essensielt for plantelivet, men bare en liten del av vannet som tilføres gjennom røttene brukes direkte til vekst og metabolisme.
Vannets kretsløp
Vannets kretsløp Det meste av vannet er konsentrert i havene. Vann som fordamper fra overflaten deres forsyner naturlige og kunstige landøkosystemer. Jo nærmere et område er havet, jo mer nedbør er det. Landet returnerer stadig vann til havet: noe av fuktigheten fordamper, mest aktivt i skoger, noe samles opp av elver: de mottar regn og smeltevann. Utveksling av fuktighet mellom hav og land krever svært høye energikostnader: omtrent 30 % av solenergien som når jorden brukes på dette.
Menneskelig påvirkning på vannets kretsløp Vannets kretsløp i biosfæren før sivilisasjonsutviklingen var i likevekt, d.v.s. havet mottok like mye vann fra elvene som det forbrukte gjennom fordampning. Med utviklingen av sivilisasjonen begynte denne syklusen å bli forstyrret. Spesielt fordamper skoger mindre og mindre vann fordi... deres areal er avtagende, og jordoverflaten, tvert imot, øker, fordi Arealet med vannet jordbruksland øker. lander. Elvene i de sørlige regionene ble grunne. Vann fordamper verre fra overflaten av havet, fordi... en betydelig del av den er dekket med en oljefilm. Alt dette forverrer vanntilførselen til biosfæren.
Tørke blir stadig hyppigere, og lommer med miljøkatastrofer dukker opp. For eksempel har en katastrofal tørke vart i mer enn 35 år i Afrika, i Sahel-sonen - en semi-ørkenregion som skiller Sahara fra de nordlige landene på kontinentet. Ferskvann som går tilbake til havet og andre vannmasser fra land er ofte forurenset. Vannet i mange russiske elver har blitt praktisk talt uegnet til å drikke. Andelen ferskvann tilgjengelig for levende organismer er ganske liten, så det må brukes sparsomt og ikke forurenses! Hver fjerde person på planeten mangler rent drikkevann. I mange områder av verden er det ikke nok vann til industriell produksjon og vanning.
Ulike komponenter i hydrosfæren deltar i vannets kretsløp på forskjellige måter og med forskjellige hastigheter. For fullstendig fornyelse av vann i isbreer tar det 8000 år, grunnvann - 5000 år, hav - 3000 år, jord - 1 år. Atmosfærisk damp og elvevann fornyes fullstendig på 10–12 dager. Vannets kretsløp i naturen tar omtrent 1 million år.
Oksygensyklus Oksygen er en del av de vanligste elementene i biosfæren. Oksygeninnholdet i atmosfæren er nesten 21 %. Oksygen er en del av vannmolekyler og levende organismer (proteiner, fett, karbohydrater, nukleinsyrer). Oksygen produseres av produsenter (grønne planter). Ozon spiller en viktig rolle i oksygensyklusen. Ozonlaget ligger i en høyde på 20-30 km over havet. Oksygeninnholdet i atmosfæren påvirkes av 2 hovedprosesser: 1) fotosyntese 2) nedbrytning av organisk materiale, hvor det forbrukes.
Oksygensyklusen er en langsom prosess. Det tar omtrent 2000 år før alt oksygenet i atmosfæren er fullstendig fornyet. Til sammenligning: fullstendig fornyelse av karbondioksid i atmosfæren skjer på omtrent 3 år. Oksygen brukes til respirasjon av de fleste levende organismer. Oksygen brukes ved forbrenning av drivstoff i forbrenningsmotorer, i ovner til termiske kraftverk, i fly- og rakettmotorer, etc. Ytterligere antropogent forbruk kan forstyrre balansen i oksygensyklusen. Så langt kompenserer biosfæren for menneskelig inngripen: tap kompenseres av grønne planter. Med ytterligere reduksjon av skogareal og forbrenning av stadig mer drivstoff vil oksygeninnholdet i atmosfæren begynne å avta.
DET ER VIKTIG!!! Når oksygeninnholdet i luften synker til 16%, forverres en persons helse (spesielt hjertet lider), til 7%, en person mister bevisstheten, og til 3% oppstår døden.
Karbon syklus
Karbonsyklus Karbon er grunnlaget for organiske forbindelser; det er en del av alle levende organismer i form av proteiner, fett og karbohydrater. Karbon kommer inn i atmosfæren i form av karbondioksid. I atmosfæren, hvor hoveddelen av karbondioksid er konsentrert, skjer det hele tiden en utveksling: planter absorberer karbondioksid under fotosyntesen, og alle organismer frigjør det under respirasjon. Opptil 50 % av karbon i form av CO 2 blir returnert til atmosfæren av nedbrytere - jordmikroorganismer. Karbon forlater syklusen som kalsiumkarbonat.
Menneskelig påvirkning på karbonkretsløpet Teknogen menneskelig aktivitet forstyrrer den naturlige balansen i karbonkretsløpet: 1) under forbrenning av organisk brensel slippes det ut ca. 6 milliarder tonn CO 2 til atmosfæren årlig: a) Elektrisitetsproduksjon ved termiske kraftverk b. ) Bileksosgasser 2) ødeleggelse av skog. I løpet av de siste 100 årene har nivået av karbondioksid i atmosfæren vokst jevnt og raskt. Karbondioksid + metan + vanndamp + ozon + nitrogenoksider = klimagass. Resultatet er drivhuseffekten – global oppvarming, som kan føre til store naturkatastrofer.
Nitrogensyklus I fri form er nitrogen en komponent av luft - 78%. Nitrogen er et av de viktigste elementene for organismers liv. Nitrogen er en del av alle proteiner. Nitrogenmolekylet er veldig sterkt, og det er grunnen til at de fleste organismer ikke klarer å absorbere atmosfærisk nitrogen. Nitrogen absorberes av levende organismer kun i form av forbindelser med hydrogen og oksygen. Fiksering av nitrogen til kjemiske forbindelser skjer som følge av vulkansk aktivitet og tordenvær, men mest som følge av aktiviteten til mikroorganismer - nitrogenfiksere (nitrogenfikserende bakterier og blågrønnalger).
Nitrogen kommer inn i røttene til planter i form av nitrater, som brukes til syntese av organisk materiale (proteiner). Dyr forbruker nitrogen gjennom plante- eller animalsk mat. Nitrogen føres tilbake til atmosfæren gjennom nedbrytning av dødt organisk materiale. Jordbakterier bryter ned proteiner til uorganiske stoffer - gasser - ammoniakk, nitrogenoksider, som kommer inn i atmosfæren. Nitrogen som kommer inn i vannmassene passerer også gjennom næringskjeden plante-dyr-mikroorganismer og går tilbake til atmosfæren.
Menneskelig påvirkning på nitrogensyklusen Teknogen menneskelig aktivitet forstyrrer den naturlige balansen i nitrogensyklusen. Når du pløyer land, reduseres aktiviteten til mikroorganismer - nitrogenfiksere nesten 5 ganger, derfor reduseres nitrogeninnholdet i jorda, noe som fører til en reduksjon i jordens fruktbarhet. Derfor introduserer folk overflødig nitrat i jorden, som er inkludert i mineralgjødsel. En stor mengde nitrogenoksider kommer inn i atmosfæren under forbrenning og prosessering av gass, olje og kull og faller i form av sur nedbør. Å gjenopprette den naturlige nitrogensyklusen er mulig ved å redusere produksjonen av nitrogengjødsel, redusere industrielle utslipp av nitrogenoksider til atmosfæren, etc.
Fosfor syklus
I motsetning til vann-, karbon-, nitrogen- og oksygensyklusene, som er lukkede, er fosforsyklusen åpen, fordi Fosfor danner ikke flyktige forbindelser som slippes ut i atmosfæren. Fosfor finnes i bergarter, hvorfra det kommer inn i økosystemene under naturlig ødeleggelse av bergarter eller når fosforgjødsel tilføres åkre. Planter absorberer uorganiske fosforforbindelser, og dyr som lever av disse plantene akkumulerer fosfor i vevet. Etter nedbrytningen av de døde kroppene til dyr og planter er ikke alt fosfor involvert i syklusen. Noe av det vaskes ut av jorden til vannmasser (elver, innsjøer, hav) og legger seg til bunnen. Fosfor kommer tilbake til land i små mengder med fisk fanget av mennesker.
Menneskelig påvirkning på fosforsyklusen Overføringen av fosfor fra land til havet har økt markant under menneskelig påvirkning. Når skog ødelegges og jordsmonn pløyes, øker volumet av overflatevannstrømmen, og i tillegg kommer fosforgjødsel tilført elver og innsjøer fra åkre. Siden fosforreservene på land er begrensede, og det er vanskelig å komme tilbake fra havet, kan det i fremtiden bli mangel på fosfor i jordbruket, noe som vil føre til nedgang i avlingene (først og fremst kornavlinger).