Energiflöde i naturen. Materias och energins kretslopp i naturen
Vilket liv som helst kräver ett konstant flöde av energi och materia. Energi läggs på att utföra grundläggande livsreaktioner, materia används för att bygga organismers kroppar. Förekomsten av naturliga ekosystem åtföljs av komplexa processer av material- och energiutbyte mellan levande och livlös natur. Dessa processer är mycket viktiga och beror inte bara på sammansättningen av biotiska samhällen, utan också på deras fysiska miljö.
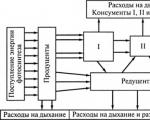
Energiflöde i samhället– detta är dess övergång från organismer på en nivå till en annan i form av kemiska bindningar av organiska föreningar (mat).
Materiens flöde (cykel) är materiens rörelse i form av kemiska grundämnen och deras föreningar från producenter till nedbrytare och sedan (genom kemiska reaktioner som sker utan medverkan av levande organismer) igen till producenter.
Materiens cirkulation och energiflödet är inte identiska begrepp, även om olika energiekvivalenter (kalorier, kilokalorier, joule) ofta används för att mäta materiens rörelse. Detta förklaras delvis av det faktum att på alla trofiska nivåer, med undantag för den första, överförs den energi som krävs för organismers liv i form av substansen i konsumerad mat. Endast växter (producenter) kan direkt använda solens strålningsenergi för sin livsaktivitet.
En strikt mätning av ett ämne som cirkulerar i ett ekosystem kan erhållas genom att ta hänsyn till cirkulationen av enskilda kemiska element, i första hand de som är huvudbyggnadsmaterialet för cytoplasman hos växt- och djurceller.
Till skillnad från ämnen som kontinuerligt cirkulerar genom olika block i ekosystemet och alltid kan komma in i kretsloppet igen, kan energi användas i kroppen endast en gång.
Enligt fysikens lagar kan energi förändras från en form (som ljusenergi) till en annan (som matens potentiella energi), men den skapas aldrig igen eller förstörs. Det kan inte finnas en enda process förknippad med omvandlingen av energi utan att förlora en del av den. I dess omvandlingar försvinner en viss mängd energi i form av värme och går därför förlorad. Av denna anledning kan det inte ske omvandlingar av till exempel livsmedelsämnen till de ämnen som utgör organismens kropp, vilket sker med hundra procent effektivitet.
Alla ekosystems existens beror på ett konstant flöde av energi, vilket är nödvändigt för att alla organismer ska kunna behålla sina vitala funktioner och självreproduktion.
Endast ungefär hälften av solflödet som faller på gröna växter absorberas av fotosyntetiska element, och endast en liten del av den absorberade energin (från 1/100 till 1/20 av delen) lagras i form av energi som behövs för aktiviteten av växtvävnader.
När man rör sig bort från primärproducenten minskar hastigheten för energiflödet (det vill säga mängden energi, uttryckt i energienheter, som överförs från en trofisk nivå till en annan) kraftigt.
Minskningen av mängden energi under övergången från en trofisk nivå till en högre bestämmer själva antalet av dessa nivåer. Det uppskattas att varje trofisk nivå bara tar emot cirka 10 % (eller något mer) av energin från den föregående nivån. Därför överstiger det totala antalet trofiska nivåer sällan 3–4.
Förhållandet mellan levande materia på olika trofiska nivåer följer i allmänhet samma regel som förhållandet mellan inkommande energi: ju högre nivå, desto lägre är den totala biomassan och antalet av dess ingående organismer.
Förhållandet mellan antalet olika grupper av organismer ger en uppfattning om samhällets stabilitet, eftersom biomassan och antalet av vissa populationer samtidigt är en indikator på livsutrymmet för organismer av denna och andra arter . Till exempel bestämmer antalet träd i en skog inte bara den totala tillgången på biomassa och energi som finns i dem, utan också mikroklimatet, såväl som antalet skyddsrum för många insekter och fåglar.
Pyramider av tal kan inverteras. Detta inträffar när bytespopulationens reproduktionshastighet är hög, och även vid låg biomassa kan en sådan population vara en tillräcklig födokälla för rovdjur som har en högre biomassa men en låg reproduktionshastighet. Till exempel kan många insekter leva och livnära sig på ett träd (en omvänd populationspyramid). Den omvända pyramiden av biomassa är karakteristisk för akvatiska ekosystem, där primärproducenterna (fytoplanktoniska alger) mycket snabbt delar sig och förökar sig i antal, och deras konsumenter (zooplanktoniska kräftdjur) är mycket större, men har en lång reproduktionscykel.
Betesmark och detritalkedjor
Energi kan flöda genom ett samhälle på många olika sätt. Den representerar alla konsumenters näringskedja(konsumentsystem) med tillägg av ytterligare två länkar: detta dött organiskt material Och nedbrytarens näringskedja(reducerande system).
Energiflöde från växter genom växtätare(de kallas bete), kallas gräsmarkens näringskedja.
Resterna av de organismer som de konsumerar och som inte används av konsumenterna fyller på dött organiskt material. Den består av avföring som innehåller en del av osmält mat, samt djurlik, vegetationsrester (löv, grenar, alger) och kallas detritus.
Flödet av energi som härrör från dött organiskt material och som passerar genom sönderdelningssystemet kallas den detritala näringskedjan.
Tillsammans med likheterna finns det djupgående skillnader i hur betesmarken och de skadliga näringskedjorna fungerar. Den består i det faktum att i i konsumentsystemet förloras avföring och döda organismer, och i det reduktiva systemet– Nej.
Förr eller senare kommer energin som finns i dött organiskt material att helt användas av nedbrytare och försvinna som värme genom andning, även om detta kräver att man passerar genom ett system av nedbrytare flera gånger. De enda undantagen är de fall då lokala abiotiska förhållanden är mycket ogynnsamma för nedbrytningsprocessen (hög luftfuktighet, permafrost). I dessa fall ackumuleras avlagringar av ofullständigt bearbetade, mycket energikrävande ämnen, som över tid och under lämpliga förhållanden omvandlas till brännbara organiska fossiler - olja, kol, torv.
Ämneskretslopp i ett ekosystem
Integriteten hos naturliga ekosystem manifesteras särskilt tydligt när man överväger de flöden av materia som cirkulerar i dem. Ämnet kan överföras i slutna kretslopp (cirkulationer), upprepade gånger cirkulerande mellan organismer och miljön.
Cirkulära rörelser (genom land, luft, vatten) av kemiska grundämnen (det vill säga ämnen) kallas biogeokemiska kretslopp eller cirkulationer.
De grundämnen och lösta salter som är nödvändiga för liv kallas konventionellt näringsämnen(ge liv) eller näringsämnen. Bland biogena element urskiljs två grupper: makrotrofa ämnen och mikrotrofa ämnen.
Makrotrofa ämnen täcka de element som utgör den kemiska basen för levande organismers vävnader. Dessa inkluderar: kol, väte, syre, kväve, fosfor, kalium, kalcium, magnesium, svavel.
Mikrotrofa ämnen inkluderar grundämnen och deras föreningar, också mycket viktiga för existensen av levande system, men i extremt små mängder. Sådana ämnen kallas ofta mikroelement. Dessa är järn, mangan, koppar, zink, bor, natrium, molybden, klor, vanadin och kobolt. Även om mikrotrofa element är väsentliga för organismer i mycket små mängder, kan deras brist allvarligt begränsa produktiviteten.
Cirkulationen av näringsämnen åtföljs vanligtvis av deras kemiska omvandlingar. Nitratkväve kan till exempel omvandlas till proteinkväve, sedan omvandlas till urea, omvandlas till ammoniak och återigen syntetiseras till nitratform under påverkan av mikroorganismer. Olika mekanismer, både biologiska och kemiska, är involverade i processerna för denitrifikation och kvävefixering.
Reserverna av näringsämnen är varierande. Processen att binda några av dem som levande biomassa minskar mängden kvar i den abiotiska miljön. Och om växter och andra organismer inte så småningom sönderfaller, skulle tillgången på näringsämnen vara uttömd och livet på jorden skulle upphöra. Av detta kan vi dra slutsatsen att aktiviteten hos heterotrofer, främst organismer som fungerar i detritala kedjor,– en avgörande faktor för att upprätthålla kretsloppet av näringsämnen och bildandet av produkter.
Låt oss titta på några numeriska data som indikerar omfattningen av ämnesöverföring, och vänder oss till den biogeokemiska kolcykeln. Den naturliga kolkällan som används av växter för syntes av organiskt material är koldioxid, som är en del av atmosfären eller är löst i vatten. Under fotosyntesen omvandlas koldioxid (koldioxid) till organiskt material som fungerar som föda för djur. Andning, jäsning och förbränning av bränslen återför koldioxid till atmosfären.
Kolreserver i atmosfären på vår planet uppskattas till 700 miljarder ton, i hydrosfären - till 50 000 miljarder ton. Enligt årliga beräkningar, som ett resultat av fotosyntes, är ökningen av växtmassa på land och i vatten 30 miljarder ton och 150 miljarder ton, respektive Kolkretsloppet fortsätter i cirka 300 år.
Ett annat exempel är fosforcykeln. De viktigaste reserverna av fosfor innehåller olika bergarter, som gradvis (som ett resultat av förstörelse och erosion) släpper ut sina fosfater till terrestra ekosystem. Fosfater konsumeras av växter och används av dem för att syntetisera organiska ämnen. När djurkroppar bryts ned av mikroorganismer återförs fosfater till jorden och används sedan igen av växter. Dessutom förs en del av fosfaterna ut av vattendrag ut i havet. Detta säkerställer utvecklingen av växtplankton och alla näringskedjor som är beroende av det. En del av fosforn som finns i havsvattnet kan återvända till land i form av guano.
ekosystemets biosfärs energicykel
Introduktion
Begrepp och struktur för ekosystem
1 Begreppet ekosystem
2 Klassificering av ekosystem
3 Zonindelning av makroekosystem
4 Ekosystemstruktur
Faktorer som säkerställer ekosystemens integritet
1 Ämnescykler
2 Energiflöde i ekosystem
3 Dynamiska processer som säkerställer integritet och hållbarhet i ekosystem
4 Biosfären som ett globalt ekosystem som säkerställer ekosystemens integritet och hållbarhet
Slutsats
Introduktion
Ett ekosystem är varje samling av organismer och oorganiska komponenter där cirkulation av ämnen kan ske. Enligt N.F. Reimers (1990), ett ekosystem är varje gemenskap av levande varelser och dess livsmiljö, förenade till en enda funktionell helhet, som uppstår på grundval av ömsesidigt beroende och orsak-och-verkan-relationer som finns mellan individuella miljökomponenter. A. Tansley (1935) föreslog följande förhållande:
Det finns mikroekosystem, mesoekosystem, makroekosystem och den globala biosfären. Stora terrestra ekosystem kallas biomer. Ekosystemen är inte utspridda i oordning, tvärtom är de grupperade i ganska regelbundna zoner, både horisontellt (på latitud) och vertikalt (i höjd).
Från ekvatorn till polerna är en viss symmetri synlig i fördelningen av biomer från olika halvklot: tropiska regnskogar, öknar, stäpper, tempererade skogar, barrskogar, taiga.
Varje ekosystem har två huvudkomponenter: organismer och faktorer från deras icke-levande miljö. Helheten av organismer (växter, djur, mikrober) kallas biota i ett ekosystem. Från den trofiska strukturens synvinkel (från den grekiska trofen - näring) kan ekosystemet delas in i två nivåer: övre, nedre.
Följande komponenter särskiljs som en del av ett ekosystem: oorganiska, organiska föreningar, luft, vatten och substratmiljöer, producenter, autotrofa organismer, konsumenter eller fagotrofer, nedbrytare och detritivorer.
Solenergi på jorden orsakar två cykler av ämnen: stora, eller geologiska, och små, biologiska (biotiska).
I ett ekosystem finns kretslopp av ämnen. De mest studerade är: kretsloppen av kol, syre, kväve, fosfor, svavel, etc.
Upprätthållandet av den vitala aktiviteten hos organismer och cirkulationen av materia i ekosystemen, det vill säga förekomsten av ekosystem, beror på det konstanta inflödet av solenergi.
I ekosystem sker ständigt förändringar i deras medlemmars tillstånd och vitala aktivitet och i förhållandet mellan befolkningar. De olika förändringar som sker i alla samhällen delas in i två huvudtyper: cykliska och progressiva.
Ett globalt ekosystem är en biosfär som kännetecknas av ekosystemens integritet och stabilitet.
1. Ekosystems koncept och struktur
1 Begreppet ekosystem
Levande organismer och deras icke-levande (abiotiska) miljö är oskiljaktigt förbundna med varandra och är i ständig interaktion. Varje enhet (biosystem) i ett ekosystem är ett ekologiskt system. Ett ekologiskt system, eller ekosystem, är den grundläggande funktionella enheten i ekologi, eftersom det inkluderar organismer och den livlösa miljön - komponenter som ömsesidigt påverkar varandras egenskaper och de nödvändiga förutsättningarna för att upprätthålla liv i den form som finns på jorden. Termen "ekosystem" föreslogs första gången 1935 av den engelske ekologen A. Tansley. För närvarande används följande definition av ekosystem i stor utsträckning. Ett ekosystem är varje samling av organismer och oorganiska komponenter där cirkulation av ämnen kan ske. Enligt N.F. Reimers (1990), ett ekosystem är varje gemenskap av levande varelser och dess livsmiljö, förenade till en enda funktionell helhet, som uppstår på grundval av ömsesidigt beroende och orsak-och-verkan-relationer som finns mellan individuella miljökomponenter. Det bör betonas att kombinationen av en specifik fysikalisk-kemisk miljö (biotop) med en gemenskap av levande organismer (biocenos) bildar ett ekosystem. A. Tansley (1935) föreslog följande förhållande:
Ekosystem = Biotop + Biocenos
Enligt definitionen av V.N. Sukachev, biogeocenosis är "en uppsättning homogena naturfenomen (atmosfär, berg, jord och hydrologiska förhållanden) över en viss utsträckning av jordens yta, som har sin egen speciella specificitet av interaktioner mellan dessa komponenter som utgör den och en viss typ av utbyte av materia och energi mellan dem själva och andra naturfenomen och representerar en internt motsägelsefull dialektisk enhet, i ständig rörelse och utveckling."
Förutom begreppen ekosystem A. Tensley och biogeocenosis V.N. Sukachev formulerade regeln av F. Evans (1956), som föreslog att man skulle använda termen "ekosystem" absolut "dimensionslöst" för att beteckna alla överorganismala levande system som interagerar med miljön. Men många författare gav termen "ekosystem" betydelsen av biogeocenos, d.v.s. elementärt ekosystem, och samtidigt högre i hierarkin av supra-biogeokoenotiska formationer upp till biosfärens ekosystem.
2 Klassificering av ekosystem
Ekosystemen som finns på jorden är olika. Det finns mikroekosystem (till exempel stammen på ett ruttnande träd), mesoekosystem (skog, damm, etc.), makroekosystem (kontinent, hav, etc.) och den globala biosfären.
Stora terrestra ekosystem kallas biomer. Varje biom innehåller ett antal mindre, sammanlänkade ekosystem. Det finns flera klassificeringar av ekosystem:
Vintergrön tropisk regnskog
Öken: Gräs och buske Chaparral - områden med regniga vintrar och torra somrar
Tropiska Graslenz och Savannah
Tempererad stäpp
Tempererad lövskog
Boreala barrskogar
Tundra: arktisk och alpin
Typer av sötvattensekosystem:
Ribbon (stilla vatten): sjöar, dammar, etc.
Lotic (strömmande vatten): floder, bäckar, etc.
Våtmarker: träsk och sumpiga skogar
Typer av marina ekosystem
Öppet hav (pelagiskt)
Kontinentalsockelvatten (kustvatten)
Uppväxtområden (bördiga områden med produktivt fiske)
Flodmynningar (kustvikar, sund, flodmynningar, saltmarker etc.). Terrestra biom särskiljs här av naturliga eller ursprungliga vegetationsegenskaper, och typer av akvatiska ekosystem kännetecknas av geologiska och fysiska egenskaper. De angivna huvudtyperna av ekosystem representerar miljön där den mänskliga civilisationen utvecklades och representerar de viktigaste biotiska samhällena som stödjer livet på jorden.
3 Zonindelning av makroekosystem
Studiet av den geografiska fördelningen av ekosystem kan endast genomföras på nivån av stora ekologiska enheter - makroekosystem, som betraktas på en kontinental skala. Ekosystemen är inte utspridda i oordning, tvärtom är de grupperade i ganska regelbundna zoner, både horisontellt (på latitud) och vertikalt (i höjd). Detta bekräftas av den periodiska lagen om geografisk zonering av A.A. Grigorieva - M.I. Budyko: med förändringen av fysisk-geografiska zoner på jorden upprepas liknande landskapszoner och några av deras allmänna egenskaper med jämna mellanrum. Detta diskuterades också när man övervägde livets mark-luft-miljö. Den periodicitet som fastställts i lag manifesteras i det faktum att värdena på torrhetsindexet varierar i olika zoner från 0 till 4-5, tre gånger mellan polerna och ekvatorn är de nära enhet. Dessa värden motsvarar den högsta biologiska produktiviteten i landskap.
Den periodiska upprepningen av egenskaper i en serie system på en hierarkisk nivå är förmodligen en allmän lag för universum, formulerad som periodicitetslagen i strukturen av systemiska aggregat, eller en systemperiodisk lag - specifika naturliga system på en nivå ( undernivå) av organisationen utgör en periodisk eller upprepande serie av morfologiskt likartade strukturer inom övre och nedre systemrum-tid-gränser, bortom vilka existensen av system på en given nivå blir omöjlig. De går in i ett instabilt tillstånd eller förvandlas till en annan systemstruktur, inklusive en annan organisationsnivå.
Temperatur och nederbörd bestämmer platsen för de viktigaste jordlevande biomen på jordens yta. Mönstret av temperatur och nederbörd i ett visst område under en ganska lång tidsperiod är vad vi kallar klimat. Klimatet i olika delar av världen är olika. Den årliga nederbörden varierar från 0 till 2500 mm eller mer. Regimer av temperatur och nederbörd kombineras på mycket olika sätt.
De specifika klimatförhållandena bestämmer i sin tur utvecklingen av en viss biom.
Från ekvatorn till polerna är en viss symmetri synlig i fördelningen av biomer från olika hemisfärer:
Tropiska regnskogar (norra Sydamerika, Centralamerika, västra och centrala delar av Ekvatorialafrika, Sydostasien, kustområden i nordvästra Australien, öar i Indiska och Stilla havet). Klimatet är utan årstidsbyte (närhet till ekvatorn), den genomsnittliga årstemperaturen är över 17°C (vanligtvis 28°C), den genomsnittliga årliga nederbörden överstiger 2400 mm.
Vegetation: skog dominerar. Vissa typer av träd är upp till 60 m höga. På deras stammar och grenar finns epifytiska växter, vars rötter inte når jorden, och vedartade vinrankor som slår rot i jorden och klättrar upp i träden till sina toppar. Allt detta bildar en tjock baldakin.
Fauna: artsammansättningen är rikare än i alla andra biomer tillsammans. Särskilt många är amfibier, reptiler och fåglar (grodor, ödlor, ormar, papegojor), apor och andra små däggdjur, exotiska insekter med ljusa färger och färgglada fiskar i reservoarer.
Funktioner: jordar är ofta tunna och fattiga, de flesta av näringsämnena finns i ytbiomassan hos rotade vegetationer.
2. Savannas (subekvatorialafrika, Sydamerika, en betydande del av södra Indien). Klimatet är torrt och varmt större delen av året. Kraftigt regn under den våta årstiden. Temperatur: genomsnittlig års-hög. Nederbörden är 750-1650 mm/år, främst under regnperioden. Vegetation - blågräs (spannmåls)växter med sällsynta lövträd. Fauna: stora växtätande däggdjur, såsom antiloper, zebror, giraffer, noshörningar, rovdjur - lejon, leoparder, geparder.
Öknar (vissa områden i Afrika, till exempel Sahara; Mellanöstern och Centralasien, Great Basin och sydvästra USA och norra Mexiko, etc.). Klimatet är mycket torrt. Temperatur - varma dagar och kalla nätter. Nederbörden är mindre än 250 mm/år. Vegetation: glesa buskar, ofta taggiga, ibland kaktusar och låga gräs, som snabbt täcker marken med en blommande matta efter sällsynta regn. Växter har omfattande ytrotsystem som fångar upp fukt från sällsynt nederbörd, samt kranrötter som penetrerar marken till grundvattennivån (30 m och djupare). Fauna: olika gnagare (kängururåtta, etc.), paddor, ödlor, ormar och andra reptiler, ugglor, örnar, gamar, småfåglar och insekter i stora mängder.
4. Stäpper (mitten av Nordamerika, Ryssland, vissa områden i Afrika och Australien, sydost om Sydamerika). Klimatet är säsongsbetonat. Temperaturer - sommartemperaturerna varierar från måttligt varmt till varmt, vintertemperaturer under 0°C. Nederbörd - 750-2000 mm/år. Vegetation: domineras av blågräs (spannmål) upp till 2 m och högre i höjden i vissa prärier i Nordamerika eller upp till 50 cm, till exempel i de ryska stäpperna, med isolerade träd och buskar i våta områden. Fauna: stora växtätande däggdjur - bison, pronghorn antilop (Nordamerika), vilda hästar (Eurasien), känguruer (Australien), giraffer, zebror, vit noshörning, antiloper (Afrika); Predatorer inkluderar prärievargar, lejon, leoparder, geparder, hyenor, en mängd olika fåglar och små grävande däggdjur som kaniner, markekorrar och jordvarkar.
5. Tempererade skogar (Västra Europa, Östasien, östra USA). Klimat - säsongsbetonat med vintertemperaturer under 0 0C. Nederbörd - 750-2000 mm/år. Vegetation: domineras av skogar av lövträd upp till 35-45 m höga (ek, hickory, lönn), buskig undervegetation, mossor, lavar. Fauna: däggdjur (vitsvanshjortar, piggsvin, tvättbjörn, opossum, ekorre, kanin, smuss), fåglar (sångare, hackspettar, koltrast, ugglor, falkar), ormar, grodor, salamander, fiskar (öring, abborre, havskatt, etc.) . ), riklig markmikrofauna (Fig. 12.3). Biotan är anpassad till det säsongsbetonade klimatet: dvala, migration, dvala under vintermånaderna. 6. Barrskogar, taiga (norra regionerna i Nordamerika, Europa och Asien). Klimatet är långa och kalla vintrar, med en del nederbörd som faller i form av snö. Vegetation: vintergröna barrskogar dominerar, mestadels gran, tall och gran. Fauna: stora växtätande klövdjur (mulhjortar, renar), små växtätande däggdjur (hare, ekorre, gnagare), varg, lo, räv, svartbjörn, grizzlybjörn, järv, mink och andra rovdjur, många blodsugande insekter på kort sommar tid. Klimatet är mycket kallt med polardagar och polarnätter. Den årliga medeltemperaturen är under -5°C. Under några veckors kort sommar tinar marken inte mer än en meter djupt. Nederbörden är mindre än 250 mm/år. Vegetation: domineras av långsamt växande lavar, mossor, gräs och starr och dvärgbuskar. Fauna: stora växtätande klövdjur (renar, myskoxe), små grävande däggdjur (året om t.ex. lämlar), rovdjur som får en kamouflagevit färg på vintern (fjällräv, lo, hermelin, snöuggla). Under den korta sommaren häckar ett stort antal flyttfåglar i tundran, bland dem finns det särskilt många sjöfåglar, som livnär sig på de insekter och sötvattensryggradslösa djur som finns i riklig mängd här. Den vertikala höjdzoneringen av markekosystem, särskilt på platser med uttalad lättnad, är också mycket tydlig. Fuktighet är den viktigaste faktorn som bestämmer typen av biom. Med tillräckligt stora mängder nederbörd utvecklas skogsvegetation. Temperaturen avgör vilken typ av skog det är. Situationen är exakt densamma i stäpp- och ökenbiomen. Förändringar i vegetationstyper i kalla områden sker med lägre årliga nederbördsmängder, eftersom mindre vatten förloras vid avdunstning vid låga temperaturer. Temperaturen blir en viktig faktor endast under mycket kalla förhållanden med permafrost. Ekosystemens sammansättning beror alltså till stor del på deras funktionella "syfte" och vice versa. Enligt N.F. Reimers (1994), detta återspeglas i principen om ekologisk komplementaritet (komplementaritet): ingen funktionell del av ekosystemet (ekologisk komponent, element etc.) kan existera utan andra funktionellt komplementära delar. Nära det och utvidga det är principen om ekologisk kongruens (korrespondens):. funktionellt kompletterar varandra utvecklar ekosystemets levande komponenter lämpliga anpassningar för detta, samordnade med förutsättningarna i den abiotiska miljön, som till stor del omvandlas av samma organismer (bioklimat etc.), d.v.s. det finns en dubbel serie av överensstämmelse - mellan organismer och deras livsmiljö - extern och skapad av cenosen. 4 Ekosystemstruktur Från den trofiska strukturens synvinkel (från den grekiska trofen - näring) kan ekosystemet delas in i två nivåer: Den övre - autotrofa (självmatande) nivån, eller "gröna bältet", inklusive växter eller deras delar som innehåller klorofyll, där fixering av räkneenergi och användning av enkla oorganiska föreningar dominerar. Den nedre är det heterotrofa (matade av andra) skiktet, eller det "bruna bältet" av jordar och sediment, ruttnande material, rötter etc., där användning, omvandling och nedbrytning av komplexa föreningar dominerar. Ur biologisk synvinkel särskiljs följande komponenter som en del av ett ekosystem: I ett ekosystem är mat- och energikopplingar mellan kategorier alltid entydiga och går i riktning mot: autotrofer - heterotrofer eller i en mer komplett form; autotrofer -> konsumenter -> nedbrytare (förstörare). Den primära energikällan för ekosystemen är solen. Energiflödet (enligt T.A. Akimova, V.V. Khaskin, 1994) som solen skickar till planeten jorden överstiger 20 miljoner EJ per år. Endast en fjärdedel av detta flöde närmar sig atmosfärens gräns. Av detta reflekteras cirka 70 %, absorberas av atmosfären och sänds ut i form av långvågig infraröd strålning. Solstrålningen som faller på jordens yta är 1,54 miljoner EJ per år. Denna enorma mängd energi är 5000 gånger mänsklighetens hela energi i slutet av 1900-talet och 5,5 gånger energin av alla tillgängliga fossila bränsleresurser av organiskt ursprung, ackumulerade under minst 100 miljoner år. Det mesta av solenergin som når planetens yta omvandlas direkt till värme, värmande vatten eller jord, vilket i sin tur värmer luften. Denna värme fungerar som drivkraften bakom vattnets kretslopp, luftströmmar och havsströmmar som bestämmer vädret, och släpps gradvis ut i yttre rymden, där den går förlorad. För att bestämma ekosystemens plats i detta naturliga energiflöde är det viktigt att inse att hur omfattande och komplexa de än är så använder de bara en liten del av det. Detta innebär en av de grundläggande principerna för ekosystemens funktion: de existerar på grund av icke-förorenande och nästan evig solenergi, vars mängd är relativt konstant och riklig. Låt oss ge mer i detalj var och en av de listade egenskaperna hos solenergi: 2. Faktorer som säkerställer ekosystemens integritet 1 Ämnescykler Båda cyklerna är ömsesidigt förbundna och representerar så att säga en enda process. Det uppskattas att allt syre som finns i atmosfären cirkulerar genom organismer (kombineras under andning och frigörs under fotosyntesen) under 2000 år, koldioxiden i atmosfären cirkulerar i motsatt riktning på 300 år, och allt vatten på jorden är sönderdelas och återskapas genom fotosyntes och andning på 2 000 000 år. Samspelet mellan abiotiska faktorer och levande organismer i ekosystemet åtföljs av en kontinuerlig cirkulation av materia mellan biotopen och biocenosen i form av alternerande organiska och mineraliska föreningar. Utbytet av kemiska element mellan levande organismer och den oorganiska miljön, vars olika stadier sker inom ett ekosystem, kallas biogeokemisk cirkulation, eller biogeokemisk cykel. Vattnets kretslopp. Den viktigaste cykeln på jorden när det gäller överförda massor och energiförbrukning är den planetariska hydrologiska cykeln - vattnets kretslopp. Varje sekund är 16,5 miljoner m3 vatten inblandade i det och mer än 40 miljarder MW solenergi spenderas på detta, (enligt T.A. Akimova V.V. Haskin, (1994)). Men denna cykel är inte bara överföringen av vattenmassor. Dessa är fasomvandlingar, bildning av lösningar och suspensioner, utfällning, kristallisation, fotosyntesprocesser samt olika kemiska reaktioner. Livet uppstod och fortsätter i denna miljö. Vatten är det grundläggande elementet som är nödvändigt för liv. Kvantitativt är det den vanligaste oorganiska komponenten i levande materia. Hos människor utgör vatten 63% av kroppsvikten, i svamp - 80%, i växter - 80-90% och hos vissa maneter - 98% Vatten, som vi kommer att se lite senare, deltar i det biologiska kretsloppet och fungerar som en källa till väte och syre, och utgör endast en liten del av dess totala volym. I flytande, fast och ånga tillstånd finns vatten i alla tre huvudkomponenterna i biosfären: atmosfären, hydrosfären och litosfären. Alla vatten förenas av det gemensamma konceptet "hydrosfär". Komponenterna i hydrosfären är sammankopplade genom konstant utbyte och interaktion. Vatten, som kontinuerligt rör sig från ett tillstånd till ett annat, gör små och stora kretslopp. Avdunstningen av vatten från havsytan, kondensationen av vattenånga i atmosfären och nederbörden på havsytan bildar en liten cykel. När vattenånga förs med luftströmmar till land blir cykeln mycket mer komplex. I det här fallet avdunstar en del av nederbörden och går tillbaka till atmosfären, den andra matar floder och reservoarer, men återvänder i slutändan till havet genom flod och underjordisk avrinning, och fullbordar därmed den stora cykeln. Biotisk (biologisk) cykel. Det biotiska (biologiska) kretsloppet avser cirkulationen av ämnen mellan jord, växter, djur och mikroorganismer. Det biotiska (biologiska) kretsloppet är flödet av kemiska grundämnen från marken, vattnet och atmosfären till levande organismer, omvandlingen av inkommande grundämnen till nya komplexa föreningar och deras återkomst tillbaka i livets process med det årliga fallet av en del av det organiska materia eller med helt döda organismer som ingår i ekosystemsammansättningen. Nu ska vi presentera det biotiska kretsloppet i en cyklisk form. Den centrala biotiska cykeln (enligt T.A. Akimova, V.V., Khaskhin) bestod av primitiva encelliga producenter (P) och nedbrytare-förstörare (D). Mikroorganismer kan snabbt föröka sig och anpassa sig till olika förhållanden, till exempel använda alla typer av substrat - kolkällor - i sin kost. Högre organismer har inte sådana förmågor. I kompletta ekosystem kan de existera i form av en struktur på grunden av mikroorganismer. Först utvecklas flercelliga växter (P) - högre producenter. Tillsammans med encelliga organismer skapar de organiskt material genom fotosyntesen, med hjälp av energin från solstrålningen. Därefter är primära konsumenter anslutna - växtätande djur (T), och sedan köttätande konsumenter. Vi undersökte markens biotiska kretslopp. Detta gäller fullt ut den biotiska cykeln i akvatiska ekosystem, till exempel havet. Alla organismer upptar en viss plats i det biotiska kretsloppet och utför sina funktioner för att omvandla grenarna av energiflödet som de tar emot och överföra biomassa. Ett system av encelliga nedbrytare (förstörare) förenar alla, avpersonifierar deras substanser och sluter den allmänna cirkeln. De återvänder till biosfärens abiotiska miljö med alla de element som behövs för nya och nya revolutioner. De viktigaste egenskaperna hos den biotiska cykeln bör betonas. Fotosyntes är en kraftfull naturlig process som årligen involverar enorma massor av biosfärmaterial i kretsloppet och bestämmer dess höga syrepotential. På grund av koldioxid och vatten syntetiseras organiskt material och fritt syre frigörs. De direkta produkterna av fotosyntes är olika organiska föreningar, och i allmänhet är fotosyntesprocessen ganska komplex. Förutom fotosyntes med deltagande av syre, den så kallade oxygeniska fotosyntesen, bör vi också fokusera på syrefri fotosyntes, eller kemosyntes. Kemosyntetiska organismer inkluderar nitrifierare, karboxidobakterier, svavelbakterier, tioniska järnbakterier och vätebakterier. De är uppkallade efter deras oxidationssubstrat, som kan vara NH3, NO2, CO, H2S, S, Fe2+, H2. Vissa arter är obligatoriska kemolitoautotrofer, andra är fakultativa. De senare inkluderar karboxidobakterier och vätebakterier. Kemosyntes är karakteristisk för hydrotermiska öppningar i djuphavet. Fotosyntes sker, med få undantag, på hela jordens yta, skapar en enorm geokemisk effekt och kan uttryckas som mängden av hela massan av kol som årligen är involverad i konstruktionen av organisk - levande materia i hela biosfären. Den allmänna kretsloppet av materia i samband med konstruktionen av organiskt material genom fotosyntes involverar också kemiska element som N, P, S, såväl som metaller - K, Ca, Mg, Na, Al. När en organism dör sker den omvända processen - nedbrytning av organiskt material genom oxidation, sönderfall, etc. med bildning av slutliga nedbrytningsprodukter. I jordens biosfär leder denna process till att mängden biomassa av levande materia tenderar att vara något konstant. Ekosfärens biomassa (2 10|2t) är sju storleksordningar mindre än massan av jordskorpan (2 .10|9t). Jordens växter producerar årligen organiskt material som motsvarar 1.6.10"% eller 8% av biomassan i ekosfären. Destruktorer, som utgör mindre än 1% av den totala biomassan av planetens organismer, bearbetar en massa organiskt material som är 10 gånger större än deras egen biomassa. I genomsnitt är biomassans förnyelseperiod 12,5 år. Låt oss anta att massan av levande materia och biosfärens produktivitet var densamma från kambrium till nutid (530 miljoner år), då den totala mängden organiskt material som passerade genom den globala biotiska cykeln och användes av livet på planeten kommer att vara 2,10" 2-5,ZL08/12,5=8,5L0|9t, vilket är 4 gånger massan av jordskorpan. Beträffande dessa beräkningar N.S. Pechurkin (1988) skrev: "Vi kan säga att atomerna som utgör våra kroppar fanns i gamla bakterier och i dinosaurier och i mammutar." Lagen om biogen migration av atomer V.I. Vernadsky säger: "Migrationen av kemiska element på jordens yta och i biosfären som helhet sker antingen med direkt deltagande av levande materia (biogen migration), eller så sker den i en miljö vars geokemiska egenskaper (O2, CO2, H2, etc.) är bestämd levande materia, både den som för närvarande lever i biosfären och den som har verkat på jorden genom geologisk historia." IN OCH. Vernadsky 1928-1930 i sina djupa generaliseringar om processer i biosfären gav han en uppfattning om de fem huvudsakliga biogeokemiska funktionerna hos levande materia. Den första funktionen är gas. Den andra funktionen är koncentration. Den tredje funktionen är redox. Den fjärde funktionen är biokemisk. Den femte funktionen är mänsklighetens biogeokemiska aktivitet, som täcker en ständigt ökande mängd materia i jordskorpan för industrins, transporternas och jordbrukets behov. Det biologiska kretsloppet varierar i olika naturliga zoner och klassificeras enligt en uppsättning indikatorer: växtbiomassa, strö, strö, mängden grundämnen fixerade i biomassan, etc. Den totala biomassan är högst i skogszonen, och andelen underjordiska organ i skogarna är lägst. Detta bekräftas av det biologiska cykelintensitetsindexet - förhållandet mellan ströets massa och den del av ströet som bildar den. Kolets kretslopp. Av alla biogeokemiska kretslopp är kolets kretslopp utan tvekan den mest intensiva. Kol cirkulerar i hög hastighet mellan olika oorganiska medel och genom näringsvävar inom samhällen av levande organismer. CO och CO2 spelar en viss roll i kolets kretslopp, ofta i jordens biosfär representeras kol av den mest rörliga formen av CO2. Källan till primär koldioxid i biosfären är vulkanisk aktivitet associerad med sekulär avgasning av manteln och de nedre horisonterna av jordskorpan. Migration av CO2 i biosfären sker på två sätt. Det första sättet är att absorbera det under fotosyntesen med bildning av glukos och andra organiska ämnen från vilka alla växtvävnader är uppbyggda. De transporteras sedan genom näringskedjor och bildar vävnader hos alla andra levande varelser i ekosystemet. När växter och djur dör på ytan sker oxidation av organiska ämnen med bildandet av CO2. Kolatomer återförs också till atmosfären när organiskt material förbränns. Ett viktigt och intressant inslag i kolets kretslopp är att i avlägsna geologiska epoker, för hundratals miljoner år sedan, användes en betydande del av det organiska materialet som skapades i fotosyntesprocesserna inte av vare sig konsumenter eller nedbrytare, utan ackumulerades i litosfären. i form av fossila bränslen: olja, kol, oljeskiffer, torv, etc. Dessa fossila bränslen bryts i enorma mängder för att möta energibehovet i vårt industrisamhälle. Genom att bränna det fullbordar vi på sätt och vis kolets kretslopp. På det andra sättet utförs kolmigrering genom att skapa ett karbonatsystem i olika reservoarer, där CO2 omvandlas till H2CO3, HCO, CO3. Med hjälp av kalcium (eller magnesium) löst i vatten fälls karbonater (CaCO3) ut genom biogena och abiogena vägar. Tjocka lager av kalksten bildas. Enligt A.B. Ronov, förhållandet mellan begravt kol i fotosyntetiska produkter och kol i karbonatstenar är 1:4. Tillsammans med det stora kolkretsloppet finns det också ett antal små kolkretslopp på landytan och i havet. I allmänhet, utan antropogen intervention, bibehålls kolhalten i biogeokemiska reservoarer: biosfären (biomassa + jord och detritus), sedimentära bergarter, atmosfär och hydrosfär med en hög grad av konstanthet (enligt T.A. Akimova, V.V. Haskin (1994) ). Det ständiga utbytet av kol, å ena sidan, mellan biosfären, och å andra sidan, mellan atmosfären och hydrosfären, bestäms av den levande materiens gasfunktion - processerna fotosyntes, andning och förstörelse, och uppgår till ca. 6-1010 ton/år Det finns ett flöde av kol till atmosfären och hydrosfären och under vulkanisk aktivitet i genomsnitt 4,5 106 t/år. Den totala massan av kol i fossila bränslen (olja, gas, kol etc.) uppskattas till 3,2*1015 ton, vilket motsvarar en genomsnittlig ackumuleringshastighet på 7 miljoner ton/år. Denna mängd är obetydlig jämfört med massan av cirkulerande kol och, som det var, föll ur cykeln och gick förlorad i den. Följaktligen är graden av öppenhet (imperfektion) av cykeln 10"4, eller 0,01%, och följaktligen är graden av stängning 99,99%. Detta betyder å ena sidan att varje kolatom deltog i cykeln. tiotusentals gånger innan de faller ut ur kretsloppet, hamnade i djupet, Å andra sidan anpassas flödena av syntes och sönderfall av organiska ämnen i biosfären till varandra med mycket hög precision. 0,2 % av det mobila kolförrådet är i konstant cirkulation. Biomassa kol förnyas om 12 år, i atmosfären - om 8 år. Syrecykel. Syre (O2) spelar en viktig roll i livet för de flesta levande organismer på vår planet. I kvantitativa termer är detta huvudkomponenten i levande materia. Till exempel, om vi tar hänsyn till vattnet som finns i vävnader, innehåller människokroppen 62,8% syre och 19,4% kol. I allmänhet är detta element i biosfären, jämfört med kol och väte, det viktigaste bland enkla ämnen. Inom biosfären sker ett snabbt utbyte av syre med levande organismer eller deras kvarlevor efter döden. Växter producerar som regel fritt syre, och djur konsumerar det genom andning. Eftersom det är det mest utbredda och mobila elementet på jorden, begränsar syre inte ekosfärens existens och funktioner, även om tillgången på syre för vattenlevande organismer kan vara tillfälligt begränsad. Syrecykeln i biosfären är extremt komplex, eftersom ett stort antal organiska och oorganiska ämnen reagerar med den. Som ett resultat uppstår många epicykler som uppstår mellan litosfären och atmosfären eller mellan hydrosfären och dessa två miljöer. Syrecykeln liknar på vissa sätt den omvända koldioxidcykeln. Den enes rörelse sker i motsatt riktning mot den andras rörelse. Förbrukningen av atmosfäriskt syre och dess ersättning av primärproducenter sker relativt snabbt. Det tar alltså 2000 år att helt förnya allt atmosfäriskt syre. Nuförtiden sker ingen syreansamling i atmosfären, och dess innehåll (20,946%) förblir konstant. I de övre lagren av atmosfären, när ultraviolett strålning verkar på syre, bildas ozon - O3 -. Cirka 5% av solenergin som når jorden går åt till bildning av ozon - cirka 8,6 * 1015 W. Reaktionerna är lätt reversibla. När ozon sönderfaller frigörs denna energi, vilket håller höga temperaturer i den övre atmosfären. Den genomsnittliga ozonkoncentrationen i atmosfären är cirka 106 vol. %; den maximala O3-koncentrationen - upp till 4-10 "* vol.% - uppnås på höjder av 20-25 km (T.A. Akimova, V.V. Haskin, 1998). Ozon fungerar som ett slags UV-filter: det blockerar en betydande del av hårda ultravioletta strålar. Förmodligen var bildningen av ozonskiktet en av förutsättningarna för att liv skulle komma ur havet och kolonisera land. Det mesta av syret som producerades under geologiska epoker fanns inte kvar i atmosfären utan fixerades av litosfären i form av karbonater, sulfater, järnoxider etc. Denna massa är 590 * 1014 ton mot 39 * 1014 ton syre, som cirkulerar i biosfären i form av gas eller sulfater lösta i kontinentala och oceaniska vatten. Kvävets kretslopp. Kväve är ett väsentligt biogent element, eftersom det är en del av proteiner och nukleinsyror. Kvävecykeln är en av de mest komplexa, eftersom den inkluderar både gas- och mineralfaser, och samtidigt de mest idealiska kretsloppen. Kvävets kretslopp är nära relaterat till kolets kretslopp. Som regel följer kväve kol, tillsammans med vilket det deltar i bildandet av alla proteinämnen. Atmosfärisk luft, som innehåller 78 % kväve, är en outtömlig reservoar. Men majoriteten av levande organismer kan inte direkt använda detta kväve. För att kväve ska tas upp av växter måste det vara en del av ammonium (NH*) eller nitratjoner (NO3). Kvävgas släpps kontinuerligt ut i atmosfären som ett resultat av arbetet med att denitrifiera bakterier, och fixerande bakterier, tillsammans med blågröna alger (cyanofyter), absorberar det hela tiden och omvandlar det till nitrater. Kvävets kretslopp är tydligt synligt på destruktörsnivå. Proteiner och andra former av organiskt kväve som finns i växter och djur exponeras efter deras död för heterotrofa bakterier, aktinomyceter, svampar (bioreducerande mikroorganismer), som producerar den energi de behöver genom att reducera detta organiska kväve och omvandla det till ammoniak. I jordar sker nitrifikationsprocessen, bestående av en kedja av reaktioner, där, med deltagande av mikroorganismer, oxidation av ammoniumjon (NH4+) till nitrit (NO~) eller nitrit till nitrat (N0~) sker. Reduktionen av nitriter och nitrater till de gasformiga föreningarna molekylärt kväve (N2) eller dikväveoxid (N20) är kärnan i denitrifikationsprocessen. Bildandet av oorganiskt nitrat i små mängder sker ständigt i atmosfären genom att atmosfäriskt kväve binds med syre under elektriska urladdningar under åskväder och sedan faller ner med regn på markytan. En annan källa till atmosfäriskt kväve är vulkaner, som kompenserar för förlusten av kväve som utesluts från kretsloppet under sedimentering eller avsättning till havens botten. Generellt sett överstiger den genomsnittliga tillförseln av nitratkväve av abiotiskt ursprung under avsättning från atmosfären till marken inte 10 kg (år/ha), fria bakterier producerar 25 kg (år/ha), medan symbiosen av Rhizobium med baljväxter producerar i genomsnitt 200 kg (år/ha). Den övervägande delen av det fixerade kvävet bearbetas genom denitrifiering av bakterier till N2 och återförs till atmosfären. Endast cirka 10 % av ammonifierat och nitrifierat kväve absorberas från marken av högre växter och hamnar till förfogande för flercelliga representanter för biocenoser Fosforcykeln. Fosforcykeln i biosfären är förknippad med metaboliska processer hos växter och djur. Detta viktiga och nödvändiga element av protoplasma, som finns i landväxter och alger 0,01-0,1%, djur från 0,1% till flera procent, cirkulerar och förvandlas gradvis från organiska föreningar till fosfater, som återigen kan användas av växter. Fosfor, till skillnad från andra biofila grundämnen, bildar dock inte en gasform under migration. Reservoaren av fosfor är inte atmosfären, som kväve, utan den mineraliska delen av litosfären. De huvudsakliga källorna till oorganisk fosfor är magmatiska bergarter (apatiter) eller sedimentära bergarter (fosforiter). Från bergarter är oorganisk fosfor involverad i cirkulationen genom urlakning och upplösning i kontinentala vatten. Genom att komma in i terrestra ekosystem och jord, absorberas fosfor av växter från en vattenlösning i form av oorganisk fosfatjon och ingår i olika organiska föreningar, där det uppträder i form av organiskt fosfat. Fosfor rör sig genom näringskedjorna från växter till andra organismer i ekosystemet. Fosfor transporteras in i akvatiska ekosystem av strömmande vatten. Floder berikar ständigt haven med fosfater. Där fosfor blir en del av växtplankton. Vissa fosforföreningar migrerar på grunda djup och konsumeras av organismer, medan den andra delen går förlorad på större djup. Döda rester av organismer leder till ackumulering av fosfor på olika djup. Svavelcykel. Det finns många gasformiga svavelföreningar, såsom svavelväte H2S och svaveldioxid SO2. Den övervägande delen av kretsloppet för detta grundämne är dock sedimentär till sin natur och förekommer i mark och vatten.Tillgängligheten av oorganiskt svavel i ekosystemet underlättas av den goda lösligheten av många sulfater i vatten. Växter, absorberar sulfater, reducerar dem och producerar svavelhaltiga aminosyror (metionin, cystein, cystin) som spelar en viktig roll i utvecklingen av den tertiära strukturen av proteiner under bildandet av disulfidbryggor mellan olika zoner i polypeptidkedjan. Många grundläggande funktioner i den biogeokemiska cykeln är tydligt synliga: S -> S -> SO2 - färglösa, gröna och lila svavelbakterier; - "H2S (anaerob reduktion av sulfat) - Desulfovibno; H2S - "SO2" (aerob oxidation av sulfid) - thiobacillus; organiskt S i SO och H2S. - aeroba respektive anaeroba heterotrofa mikroorganismer. Primärproduktion säkerställer införlivandet av sulfat i organiskt material, och utsöndring av djur fungerar som ett sätt att återföra sulfat till kretsloppet. I allmänhet kräver ekosystemet mindre svavel jämfört med kväve och fosfor. Därför är svavel mer sällan en begränsande faktor för växter och djur. Samtidigt är svavelcykeln en nyckel i den övergripande processen för produktion och nedbrytning av biomassa. Till exempel, när järnsulfider bildas i sediment, överförs fosfor från en olöslig form till en löslig form och blir tillgänglig för organismer. Detta är en bekräftelse på hur en cykel regleras av en annan. 2 Energiflöde i ekosystem Till skillnad från ämnen som kontinuerligt cirkulerar genom olika block av ekosystemet, som alltid kan återanvändas och komma in i kretsloppet, kan energi användas en gång, d.v.s. det finns ett linjärt flöde av energi genom ekosystemet. Envägsinflödet av energi som ett universellt naturfenomen uppstår som ett resultat av termodynamikens lagar. Den första lagen säger att energi kan omvandlas från en form (som ljus) till en annan (som matens potentiella energi), men kan inte skapas eller förstöras. Den andra lagen säger att det inte kan finnas en enda process förknippad med omvandlingen av energi utan att förlora en del av den. En viss mängd energi i sådana transformationer försvinner till otillgänglig termisk energi och går därför förlorad. Därför: det kan inte förekomma omvandlingar, till exempel, av livsmedelsämnen till det ämne som utgör organismens kropp, som sker med 100 % effektivitet. Således är levande organismer energiomvandlare. Och varje gång energi omvandlas går en del av den förlorad i form av värme. I slutändan försvinner all energi som kommer in i den biotiska cykeln i ett ekosystem som värme. Levande organismer använder faktiskt inte värme som energikälla för att utföra arbete – de använder ljus och kemisk energi. Livsmedelskedjor och nätverk, trofiska nivåer. Inom ett ekosystem skapas energihaltiga ämnen av autotrofa organismer och fungerar som föda för heterotrofer. Matkopplingar är mekanismer för att överföra energi från en organism till en annan. Ett typiskt exempel: ett djur äter växter. Detta djur kan i sin tur ätas av ett annat djur. På så sätt kan energi överföras genom ett antal organismer - varje efterföljande livnär sig på den föregående, som förser den med råvaror och energi. Denna sekvens av energiöverföring kallas en livsmedelskedja (trofisk) eller näringskedja. Placeringen av varje länk i näringskedjan är en trofisk nivå. Den första trofiska nivån, som nämnts tidigare, upptas av autotrofer, eller så kallade primära producenter. Organismer på den andra trofiska nivån kallas primära konsumenter, den tredje - sekundära konsumenter, etc. Det finns generellt tre typer av näringskedjor: Köttätarnas näringskedja börjar med växter och går från små organismer till allt större organismer. På land består näringskedjor av tre till fyra länkar. En av de enklaste näringskedjorna ser ut så här: växt -> hare -> varg producent -" växtätare -> -> köttätare Följande näringskedjor är också utbredda: Växtmaterial (t.ex. nektar) - "fluga -" -" spindel -> shrew -> uggla. Rosenbusksav -> bladlöss -> -> nyckelpiga -> -> spindel - "insektätande fågel -> rovfågel. I akvatiska och i synnerhet marina ekosystem tenderar rovdjurens näringskedjor att vara längre än i terrestra. Den tredje typen av näringskedjor, som börjar med döda växtrester, kadaver och djurexkrementer, kallas detritala (saprofytiska) näringskedjor eller detritala nedbrytningskedjor. Lövskogar spelar en viktig roll i de skadliga näringskedjorna i terrestra ekosystem, där det mesta av lövverket inte konsumeras av växtätare och är en del av skräpet av nedfallna löv. Bladen krossas av många detritivorer - svampar, bakterier, insekter (till exempel collembola), etc., och intas sedan av daggmaskar, som jämnt fördelar humus i jordens ytskikt och bildar den så kallade mull. På denna nivå utvecklar svampar mycelium. De sönderfallande mikroorganismerna som fullbordar kedjan producerar den slutliga mineraliseringen av dött organiskt material. I allmänhet kan typiska detritala näringskedjor i våra skogar representeras enligt följande: Lövströ -> daggmask -> koltrast - sparvhök; Dött djur - "larver av kadaverflugor -" gräsgroda -> vanlig gräsorm. I de diskuterade näringskedjediagrammen representeras varje organism som livnär sig på andra organismer av en typ. De faktiska födokopplingarna i ett ekosystem är mycket mer komplexa, eftersom ett djur kan livnära sig på organismer av olika typer från samma näringskedja eller från olika näringskedjor, till exempel rovdjur på de övre trofiska nivåerna. Djur livnär sig ofta på både växter och andra djur. De kallas allätare. Matnät i ekosystem är mycket komplexa, och vi kan dra slutsatsen att energin som kommer in i dem tar lång tid att migrera från en organism till en annan. Ekologiska pyramider. Inom varje ekosystem har näringsnäten en väldefinierad struktur, som kännetecknas av arten och antalet organismer representerade på varje nivå i de olika näringskedjorna. För att studera sambanden mellan organismer i ett ekosystem och för att avbilda dem grafiskt använder de sig vanligtvis av ekologiska pyramider snarare än diagram i näringsväven. Ekologiska pyramider uttrycker den trofiska strukturen i ett ekosystem i geometrisk form. De är konstruerade i form av rektanglar med samma bredd, men längden på rektanglarna måste vara proportionell mot värdet på objektet som mäts. Härifrån kan du hämta pyramider av siffror, biomassa och energi. Ekologiska pyramider återspeglar de grundläggande egenskaperna hos varje biocenos när de visar sin trofiska struktur. Pyramider av siffror. De representerar den enklaste approximationen till studiet av den trofiska strukturen i ett ekosystem. Biomassa pyramid. Återspeglar mer fullständigt näringsförhållandena i ekosystemet, eftersom det tar hänsyn till den totala massan av organismer (biomassa) för varje trofisk nivå. Pyramid av energi. Det mest grundläggande sättet att visa kopplingar mellan organismer på olika trofiska nivåer är genom energipyramider. De representerar energiomvandlingseffektiviteten och produktiviteten i näringskedjorna och är konstruerade genom att räkna mängden energi (kcal) som ackumuleras per ytenhet per tidsenhet och som används av organismer på varje trofisk nivå. Solenergin som växten tar emot används endast delvis i fotosyntesprocessen. Energin som är fixerad i kolhydrater representerar ekosystemets bruttoproduktion.Kolhydrater används för att bygga protoplasma och växttillväxt. En del av deras energi går åt till att andas. Andra ordningens konsumenter (rovdjur) förstör inte hela biomassan från sina offer. Dessutom, av den mängd som de förstör, används bara en del för att skapa biomassa av deras egen trofiska nivå. Resten går främst till andningsenergi och utsöndras med exkrementer och exkrementer. År 1942 formulerade R. Lindeman för första gången lagen om energipyramiden, som i läroböcker ofta kallas "lagen om 10%". Enligt denna lag rör sig i genomsnitt inte mer än 10% av energin från en trofisk nivå i den ekologiska pyramiden till en annan nivå. Endast 10-20% av den initiala energin överförs till efterföljande heterotrofer. Med hjälp av energipyramidens lag är det lätt att beräkna att mängden energi som når tertiära köttätare (trofisk nivå V) är cirka 0,0001 av den energi som absorberas av producenter. Det följer att överföringen av energi från en nivå till en annan sker med mycket låg effektivitet. Detta förklarar det begränsade antalet länkar i näringskedjan, oavsett en viss biocenos. E. Odum (1959) i en extremt förenklad näringskedja - alfalfa -> kalv - "bedömde barnet omvandlingen av energi, illustrerade omfattningen av dess förluster. Låt oss säga, resonerade han, det finns alfalfa grödor på ett område med 4 hektar. Kalvar äter på detta fält (det antas att de bara äter alfalfa), och en 12-årig pojke äter uteslutande kalvkött. Resultaten av beräkningar, presenterade i form av tre pyramider: antal, biomassa och energi, indikerar att alfalfa bara använder 0,24 % av all solenergi som faller på fältet, kalven absorberar 8 % av denna produkt och endast 0,7 % av kalvens biomassa säkerställer barnets utveckling under året. E. Odum visade alltså att endast en miljondel av den infallande solenergin omvandlas till köttätande biomassa, vilket i detta fall bidrar till en ökning av barnets vikt, och resten går förlorad och försvinner i en degraderad form i miljön. Ovanstående exempel illustrerar tydligt ekosystemens mycket låga ekologiska effektivitet och den låga effektiviteten av omvandling i näringskedjor. Vi kan ange följande: om 1000 kcal (dag m 2) inspelad av producenter, sedan 10 kcal (dag m 2) går in i växtätares biomassa och endast 1 kcal (dag m 2) - in i köttätares biomassa. Eftersom en viss mängd av ett ämne kan användas av varje biocenos upprepade gånger, och en del energi en gång, är det mer ändamålsenligt att säga att en kaskadöverföring av energi sker i ekosystemet. Konsumenter fungerar som en hanterande och stabiliserande länk i ekosystemet. Konsumenter genererar ett spektrum av mångfald i cenosen, vilket förhindrar dominanternas monopol. Regeln om konsumenternas kontrollerande värde kan med rätta anses vara ganska grundläggande. Enligt cybernetiska synsätt bör kontrollsystemet vara mer komplext till sin struktur än det kontrollerade, då blir orsaken till mångfalden av typer av instrument tydlig. Konsumenternas kontrollerande betydelse har också en energisk grund. Energiflödet genom en eller annan trofisk nivå kan inte absolut bestämmas av tillgången på mat i den underliggande trofiska nivån. Som bekant finns det alltid en tillräcklig "reserv" kvar, eftersom en fullständig förstörelse av livsmedel skulle leda till konsumenternas död. Dessa allmänna mönster observeras inom ramen för befolkningsprocesser, samhällen, nivåer av den ekologiska pyramiden och biocenoser som helhet. 3 Dynamiska processer som säkerställer integritet och hållbarhet i ekosystem Cykliska förändringar i samhällen återspeglar den dagliga, säsongsbetonade och långsiktiga periodiciteten av yttre förhållanden och manifestationen av endogena rytmer hos organismer. Ekosystemens dagliga dynamik är främst förknippad med naturfenomens rytm och är strikt periodisk till sin natur. Vi har redan ansett att det i varje biocenos finns grupper av organismer vars livsaktivitet sker vid olika tidpunkter på dygnet. Vissa är aktiva på dagen, andra på natten. Därför inträffar periodiska förändringar i sammansättningen och förhållandet mellan enskilda typer av biocenos i ett visst ekosystem, eftersom enskilda organismer stängs av från det under en viss tid. Den dagliga dynamiken i biocenosen tillhandahålls av både djur och växter. Som bekant förändras intensiteten och naturen hos fysiologiska processer i växter under dagen - fotosyntes sker inte på natten, ofta blommor i växter öppnar endast på natten och pollineras av nattdjur, andra är anpassade till pollinering under dagen. Daglig dynamik i biocenoser är som regel mer uttalad, ju större skillnaden är i temperatur, luftfuktighet och andra miljöfaktorer mellan dag och natt. Mer betydande avvikelser i biocenoser observeras med säsongsbetonad dynamik. Detta beror på organismernas biologiska cykler, som beror på naturfenomens säsongsmässiga cyklicitet. Säsongsbytet har således en betydande inverkan på djurs och växters livsaktivitet (dvala, vintersömn, diapaus och migration hos djur; perioder av blomning, fruktsättning, aktiv tillväxt, lövfall och vintervila hos växter). Biocenosens skiktade struktur är ofta föremål för säsongsvariationer. Enskilda lager av växter kan helt försvinna under de lämpliga årstiderna på året, till exempel ett örtartat lager som består av ettåriga växter. De biologiska årstidernas längd varierar på olika breddgrader. I detta avseende är säsongsdynamiken för biocenoser i de arktiska, tempererade och tropiska zonerna olika. Tydligast uttrycks det i tempererade klimatekosystem och nordliga breddgrader. Långtidsvariation är normal i livet för alla biocenoser. Således fluktuerar mängden nederbörd som faller i Barabinsks skogssteppe kraftigt från år till år, ett antal torra år växlar med långvariga perioder med riklig nederbörd. Detta har en betydande inverkan på växter och djur. I det här fallet sker utvecklingen av ekologiska nischer - funktionell avgränsning i den framväxande uppsättningen eller dess tillägg med liten mångfald. Långvariga förändringar i sammansättningen av biocenoser upprepas också i samband med periodiska förändringar i atmosfärens allmänna cirkulation, vilket i sin tur orsakas av en ökning eller minskning av solaktiviteten. I processen med daglig och säsongsbetonad dynamik kränks integriteten hos biocenoser vanligtvis inte. Biocenosen upplever endast periodiska fluktuationer i kvalitativa och kvantitativa egenskaper. Progressiva förändringar i ekosystemet leder i slutändan till att en biocenos ersätts med en annan, med en annan uppsättning dominerande arter. Orsakerna till sådana förändringar kan vara faktorer utanför biocenosen som under lång tid verkar i en riktning, till exempel ökad förorening av vattenförekomster, ökad uttorkning av sumpmarker till följd av återvinning, ökat bete m.m. Dessa förändringar från en biocenos till en annan kallas exogenetiska. I det fall när det ökande inflytandet av en faktor leder till en gradvis förenkling av biocenosens struktur, utarmning av deras sammansättning och en minskning av produktiviteten, kallas sådana förskjutningar digressiva eller digressiva. Endogenetiska förändringar uppstår som ett resultat av processer som sker inom själva biocenosen. Den successiva ersättningen av en biocenos med en annan kallas ekologisk succession (från lat. - succession - sekvens, förändring). Succession är en process av självutveckling av ekosystem. Succession baseras på ofullständigheten i den biologiska cykeln i en given biocenos. Det är känt att levande organismer, som ett resultat av sin vitala aktivitet, förändrar miljön runt dem, tar bort vissa ämnen från den och mättar den med metaboliska produkter. När populationer existerar under en relativt lång tid ändrar de sin miljö i en ogynnsam riktning och som ett resultat blir de undanträngda av populationer av andra arter, för vilka de resulterande miljöomvandlingarna visar sig vara ekologiskt fördelaktiga. I en biocenos sker alltså en förändring av dominerande arter. Regeln (principen) om ekologisk dubbelarbete är tydligt synlig här. Den långsiktiga existensen av en biocenos är möjlig endast om förändringar i miljön orsakade av vissa levande organismers aktivitet är gynnsamma för andra med motsatta krav. Baserat på konkurrerande interaktioner mellan arter under succession, bildas gradvis mer stabila kombinationer som motsvarar specifika abiotiska miljöförhållanden. Ett exempel på succession som leder till att ett samhälle ersätts med ett annat är igenväxt av en liten sjö följt av uppkomsten av ett träsk och sedan en skog i dess ställe. Först bildas en flytande matta längs kanterna av sjön - en flytande matta av säd, mossor och andra växter. Sjön fylls ständigt med döda växtrester - torv. Ett träsk bildas, gradvis bevuxet med skog. En successiv serie gemenskaper som gradvis och naturligt ersätter varandra i följd kallas en successionsserie. Successioner i naturen är extremt varierande i omfattning. De kan observeras i banker med kulturer, som är planktonsamhällen - olika typer av flytande alger och deras konsumenter - hjuldjur, flagellater i vattenpölar och dammar, träsk, ängar, skogar, övergivna åkermarker, vittrade stenar etc. Det finns en hierarki. i organiseringen av ekosystem Det visar sig också i successionsprocesser - större omvandlingar av biocenoser består av mindre. I stabila ekosystem med ett reglerat kretslopp av ämnen sker också ständigt lokala successiva förändringar, vilket stöder samhällenas komplexa interna struktur. Typer av successiva förändringar. Det finns två huvudtyper av successiva förändringar: 1 - med deltagande av autotrofa och heterotrofa populationer; 2 - med deltagande av endast heterotrofer. Succession av den andra typen sker endast under förhållanden där en preliminär tillförsel eller konstant tillförsel av organiska föreningar skapas, på grund av vilket samhället existerar: i högar eller högar av gödsel, i sönderfallande växtmaterial, i reservoarer förorenade med organiska ämnen, etc. Successionsprocessen. Enligt F. Clements (1916) består successionsprocessen av följande steg: Succession med byte av vegetation kan vara primär och sekundär. Primär succession är processen för utveckling och förändring av ekosystem i tidigare obebodda områden, som börjar med deras kolonisering. Ett klassiskt exempel är den ständiga nedsmutsningen av kala stenar med den eventuella utvecklingen av skogar på dem. Således, i de primära följderna som förekommer på klipporna i Uralbergen, urskiljs följande stadier: Sekundär succession är återställandet av ett ekosystem som en gång funnits i ett givet område. Det börjar när de etablerade relationerna mellan organismer i en etablerad biocenos störs till följd av ett vulkanutbrott, brand, avverkning, plöjning etc. Förskjutningar som leder till återställandet av den tidigare biocenosen kallas demutation inom geobotanik. Ett exempel är dynamiken i artmångfalden på ön Krakatoa efter den fullständiga förstörelsen av inhemsk flora och fauna av en vulkan. Ett annat exempel är den sekundära följden av den sibiriska mörka barrskogen (grancedar taiga) efter en förödande skogsbrand. I mer brända områden uppstår pionjärmossor från sporer som blåser av vinden: 3-5 år efter en brand är den vanligaste "eldmossan" Funaria hygrometrica, Geratodon purpureus etc. Bland högre växter kommer eldgräs (Chamaenerion angustifolium) mycket snabbt koloniserar brända områden ), som redan efter 2-3 månader blommar rikligt i elden, samt markrör (Calamagrostis epigeios) och andra arter. Ytterligare successionsfaser observeras: vassängen ger vika för buskar, sedan björk- eller aspskog, tallblandskog, tallskog, tall-cederskog och slutligen, efter 250 år, sker återställandet av ceder-granskogen . Sekundära successioner sker som regel snabbare och lättare än primära, eftersom markprofilen, frön, primordier och en del av den tidigare populationen och tidigare kopplingar i den störda livsmiljön bevaras. Demutation är inte en upprepning av något stadium av primär succession. Klimax ekosystem. Succession slutar med ett skede då alla arter i ekosystemet, medan de reproducerar, bibehåller ett relativt konstant antal och ingen ytterligare förändring i dess sammansättning sker. Detta jämviktstillstånd kallas klimax, och ekosystemet kallas klimax. Under olika abiotiska förhållanden bildas olika klimax-ekosystem. I ett varmt och fuktigt klimat blir det en tropisk regnskog, i ett torrt och varmt klimat blir det en öken. Jordens huvudsakliga biomer är ekosystemens klimax i sina respektive geografiska områden. Förändringar i ekosystemet under succession. Produktivitet och biomassa. Som redan nämnts är succession en naturlig, riktad process och de förändringar som sker i ett eller annat skede är karakteristiska för alla samhällen och är inte beroende av dess artsammansättning eller geografiska läge. Det finns fyra huvudtyper av successiva förändringar: Det bör också noteras att bytet av successionsfaserna sker i enlighet med vissa regler. Varje fas förbereder miljön för uppkomsten av nästa. Lagen om sekvensen av utvecklingsfasernas gång fungerar här; utvecklingsfaserna för ett naturligt system kan följa endast i en evolutionärt fixerad (historiskt, miljömässigt bestämd) ordning, vanligtvis från relativt enkel till komplex, som regel, utan förlust av mellanstadier, men kanske med sin mycket snabba passage eller evolutionärt fixerade frånvaro. När ett ekosystem närmar sig ett tillstånd av klimakteriet, i det, som i alla jämviktssystem, saktar alla utvecklingsprocesser ner. Denna situation återspeglas i lagen om successiv avmattning: processer som sker i mogna jämviktsekosystem som är i ett stabilt tillstånd tenderar som regel att sakta ner. I det här fallet ändras den restaurerande typen av succession till deras sekulära kurs, d.v.s. självutveckling sker inom gränserna för klimakteriet eller nodal utveckling. Den empiriska lagen om successiv avmattning är en konsekvens av regeln från G. Odum och R. Pinkerton, eller regeln om maximal energi för att upprätthålla ett moget system: succession fortsätter i riktning mot en fundamental förändring i energiflödet mot att öka dess kvantitet, som syftar till att upprätthålla systemet. Regeln för G. Odum och R. Pinkerton bygger i sin tur på regeln om maximal energi i biologiska system, formulerad av A. Lotka. Denna fråga utvecklades senare väl av R. Margalef, Y. Odum och är känd som bevis på principen om "nollmaximum", eller minimering av tillväxt i ett moget ekosystem: ett ekosystem i successiv utveckling tenderar att bilda den största biomassan med minsta biologiska produktivitet. Lindeman (1942) bevisade experimentellt att succession åtföljs av en ökning av produktiviteten upp till klimaxgemenskapen, där energiomvandlingen sker mest effektivt. Data från studier av följden av ek- och ekskogar visar att deras produktivitet faktiskt ökar i senare skeden. Men under övergången till en klimaxgemenskap sker vanligtvis en minskning av den totala produktiviteten. Produktiviteten i gammal skog är alltså lägre än i ungskog, som i sin tur kan vara mindre produktiv än de mer artrika örtartade skikten som föregår dem. Liknande minskningar av produktiviteten har observerats i vissa vattensystem. Det finns flera anledningar till detta. En av dem är att ansamling av näringsämnen i växande skogsbiomassa kan leda till att deras cykling minskar. Nedgången i den totala produktiviteten kan helt enkelt vara resultatet av minskad vitalitet hos individer när deras medelålder i samhället ökar. När successionen fortskrider ackumuleras en ökande andel av tillgängliga näringsämnen i samhällets biomassa, och följaktligen minskar deras innehåll i ekosystemets abiotiska komponent (i mark eller vatten). Mängden av detritus som produceras ökar också. De huvudsakliga primära konsumenterna är inte växtätare, utan skadliga organismer. Motsvarande förändringar sker även i trofiska nätverk. Detritus blir den huvudsakliga källan till näringsämnen. Under succession ökar slutenheten i biogeokemiska cykler av ämnen. Ungefär 10 år från det att restaureringen av vegetationstäcket börjar, minskar cyklernas öppenhet från 100 till 10%, och sedan minskar den ännu mer och når ett minimum i klimaxfasen. Regeln om att öka stängningen av den biogeokemiska cykeln av ämnen under succession, kan det sägas med all säkerhet, kränks av den antropogena omvandlingen av vegetation och naturliga ekosystem i allmänhet. Detta leder utan tvekan till en lång rad anomalier i biosfären och dess indelningar. En minskning av artmångfalden under klimakteriet betyder inte dess låga ekologiska betydelse. Mångfalden av arter formar succession, dess riktning och säkerställer att det verkliga rummet fylls med liv. Ett otillräckligt antal arter som utgör komplexet kunde inte bilda en följdserie, och gradvis, med förstörelsen av klimax-ekosystem, skulle fullständig ökenspridning av planeten inträffa. Värdet av mångfald är funktionellt både statiskt och dynamiskt. Det bör noteras att där mångfalden av arter inte räcker till för att bilda biosfären, som fungerar som grunden för det normala naturliga förloppet av successionsprocessen, och miljön i sig är kraftigt störd, når successionen inte klimaxfasen, men slutar med en nodalgemenskap - ett paraklimax, en långsiktig eller kortsiktig härledd gemenskap. Ju djupare störningen av miljön i ett visst utrymme är, desto tidigare faser slutar successionen. När en eller en grupp av arter går förlorade som ett resultat av deras förstörelse (antropogent försvinnande av livsmiljöer, mer sällan utrotning), är uppnåendet av klimakteriet inte en fullständig återställande av den naturliga miljön. I själva verket är detta ett nytt ekosystem, eftersom nya kopplingar har uppstått i det, många gamla har gått förlorade och en annan "fransning" av arter har utvecklats. Ekosystemet kan inte återgå till sitt gamla tillstånd, eftersom en förlorad art inte kan återställas. När någon abiotisk eller biotisk faktor förändras, till exempel med ihållande avkylning, eller introduktion av en ny art, kommer en art som är dåligt anpassad till nya förhållanden att möta en av tre vägar. Lagen om evolutionär-ekologisk irreversibilitet säger att ett ekosystem som har förlorat en del av sina element eller har ersatts av ett annat till följd av en obalans av ekologiska komponenter inte kan återgå till sitt ursprungliga tillstånd under succession om det under förändringarna är evolutionärt (mikroevolutionärt). ) förändringar har inträffat i ekologiska element (bevarade eller tillfälligt förlorade). I fallet när vissa arter går förlorade i mellanliggande faser av succession kan denna förlust funktionellt kompenseras, men inte helt. När mångfalden minskar bortom en kritisk nivå förvrängs successionsförloppet, och i själva verket kan en klimax identisk med det förflutna inte uppnås. För att bedöma arten av återställda ekosystem är lagen om evolutionär-ekologisk irreversibilitet viktig. Med förlusten av grundämnen är det i själva verket helt nya ekologiska naturformationer med nybildade mönster och samband. Överföringen i det förflutna av en art som hoppade ur ekosystemet under sin återklimatisering betyder alltså inte dess mekaniska återkomst. Detta är faktiskt införandet av en ny art i ett förnyat ekosystem Lagen om evolutionär-ekologisk irreversibilitet betonar evolutionens riktning inte bara på biosystemsnivå, utan även på alla andra hierarkiska nivåer i biotan. 4 Biosfären som ett globalt ekosystem som säkerställer ekosystemens integritet och hållbarhet Biosfären är ett globalt ekosystem. Som noterats tidigare är biosfären uppdelad i geobiosfär, hydrobiosfär och aerobiosfär (Fig. 2.4). Geobiosfären har indelningar i enlighet med de huvudsakliga miljöbildande faktorerna: terrabiosfären och lithobiosfären - inom geobiosfären, marinobiosfären (oceanobiosfären) och akvabiosfären - inom hydrobiosfären. Dessa formationer kallas undersfärer. Den ledande miljöbildande faktorn i deras bildning är den fysiska fasen av livsmiljön: luft-vatten i aerobiosfären, vatten - sötvatten och saltvatten i hydrobiosfären, fast luft i terrabiosfären och fast vatten i litobiosfären. I sin tur faller de alla i lager: aerobiosfären - in i tropobiosfären och altobiosfären; hydrobiosfär - till fotosfär, disfotosfär och afotosfär. De strukturbildande faktorerna här, förutom den fysiska miljön, energi (ljus och värme), speciella förutsättningar för bildning och utveckling av liv - de evolutionära riktningarna för penetration av biota på land, i dess djup, in i utrymmena ovanför jorden, havets avgrund, är utan tvekan annorlunda. Tillsammans med apobiosfären, parabiosfären och andra sub- och suprabiosfäriska lager utgör de den så kallade "livets lagerkaka" och geosfären (ekosfären) för dess existens inom megabiosfärens gränser. I systemisk mening är de listade formationerna stora funktionella delar av praktiskt taget universella eller subplanetära dimensioner. Forskare tror att det i biosfären finns minst 8-9 nivåer av relativt oberoende cykler av ämnen inom sammankopplingen av 7 huvudsakliga material-energi-ekologiska komponenter och den 8:e - informativ. Globala, regionala och lokala kretslopp av ämnen är inte slutna och "korsar sig" delvis inom ekosystemhierarkin. Denna material-energi och delvis informativa "koppling" säkerställer integriteten hos ekologiska supersystem upp till biosfären som helhet. Ekosystemens integritet och hållbarhet. Biosfären bildas i större utsträckning inte av yttre faktorer, utan av inre mönster. Den viktigaste egenskapen hos biosfären är interaktionen mellan levande och icke-levande saker, vilket återspeglas i lagen om biogen migration av atomer av V. I. Vernadsky. Lagen om biogen migration av atomer gör det möjligt för mänskligheten att medvetet kontrollera biogeokemiska processer både på jorden som helhet och i dess regioner. Mängden levande materia i biosfären är som bekant inte föremål för märkbara förändringar. Detta mönster formulerades i form av lagen om beständighet för mängden levande materia av V.I. Vernadsky: mängden levande materia i biosfären för en given geologisk period är konstant. I praktiken är denna lag en kvantitativ konsekvens av lagen om intern dynamisk jämvikt för det globala ekosystemet - biosfären. Eftersom levande materia, i enlighet med lagen om biogen migration av atomer, är en energiförmedlare mellan solen och jorden, måste dess kvantitet antingen vara konstant, eller så måste dess energiegenskaper förändras. Lagen om fysisk och kemisk enhet av levande materia (all levande materia på jorden är fysisk och kemiskt förenad och utesluter betydande förändringar i den sistnämnda egenskapen. Kvantitativ stabilitet är därför oundviklig för den levande materien på planeten. Den är helt kännetecknande för antalet arter. Levande materia, som en ackumulator av solenergi, måste samtidigt reagera på både yttre (kosmiska) påverkan och inre förändringar. En minskning eller ökning av mängden levande materia på en plats i biosfären bör leda till en process precis motsatsen på en annan plats, eftersom de frigjorda näringsämnena kan assimileras av resten av den levande materien eller deras brist kommer att observeras. Här måste vi ta hänsyn till processens hastighet, som vid antropogen förändring är mycket lägre än direkt störning av naturen av människan. Förutom beständigheten och beständigheten för mängden levande materia, vilket återspeglas i lagen om fysisk och kemisk enhet av levande materia, finns i den levande naturen ett konstant bevarande av den informationsmässiga och somatiska strukturen, trots att den förändras något med evolutionens gång. Denna egenskap noterades av Yu. Goldsmith (1981) och kallades lagen om bevarande av biosfärens struktur - informativ och somatisk, eller ekodynamikens första lag. För att bevara biosfärens struktur strävar levande varelser efter att uppnå ett tillstånd av mognad eller ekologisk balans. Lagen om önskan om klimakteriet - den andra lagen för ekodynamiken av Yu. Goldsmith, gäller biosfären och andra nivåer av ekologiska system, även om det finns detaljer - biosfären är ett mer slutet system än dess underavdelningar. Enheten av den levande materien i biosfären och homologin i strukturen av dess subsystem leder till det faktum att de levande elementen från olika geologiska åldrar och ursprungliga geografiska ursprung som uppstod på den är intrikat sammanflätade evolutionärt. Sammanvävningen av element från olika rums-temporala tillkomster på alla ekologiska nivåer i biosfären återspeglar regeln eller principen om heterogenes av levande materia. Detta tillägg är inte kaotiskt, utan är föremål för principerna om ekologisk komplementaritet, ekologisk överensstämmelse (kongruens) och andra lagar. Inom ramen för Yu. Goldsmiths ekodynamik är detta dess tredje lag - principen om ekologisk ordning, eller ekologisk mutualism, som indikerar en global egenskap på grund av helhetens inflytande på dess delar, det omvända inflytandet av differentierade delar på utveckling av helheten etc. vilket sammantaget leder till bevarandestabiliteten för biosfären som helhet. Ömsesidig hjälp inom ramen för den ekologiska ordningen, eller systemisk ömsesidighet, bekräftas av lagen om ordning och reda för att fylla utrymme och rums-temporal säkerhet: fyllningen av utrymmet inom ett naturligt system, på grund av interaktionen mellan dess delsystem, ordnas i ett sådant sätt som gör att systemets homeostatiska egenskaper kan realiseras med minimala motsättningar mellan delarna inom det. Av denna lag följer att den långsiktiga förekomsten av olyckor som är "onödiga" för naturen, inklusive mänskliga skapelser främmande för den, är omöjlig. Reglerna för den mutualistiska systemordningen i biosfären innefattar också principen om systemkomplementaritet, som säger att delsystemen i ett naturligt system i sin utveckling utgör en förutsättning för framgångsrik utveckling och självreglering av andra delsystem som ingår i samma system. Den fjärde ekodynamikens lag av Yu. Goldsmith inkluderar lagen om självkontroll och självreglering av levande varelser: levande system och system under kontrollerande inflytande av levande varelser är kapabla till självkontroll och självreglering i processen för deras anpassning till förändringar i omgivningen. I biosfären sker självkontroll och självreglering under kaskad- och kedjeprocesser av allmän interaktion - under kampen för existensen av naturligt urval (i den vidaste bemärkelsen av detta koncept), anpassning av system och delsystem, bred samevolution , etc. Dessutom leder alla dessa processer till positiva resultat "ur naturens synvinkel" - bevarande och utveckling av biosfärens ekosystem och den som helhet. Den sammanbindande länken mellan generaliseringar av strukturell och evolutionär karaktär är regeln för automatiskt underhåll av den globala livsmiljön: levande materia, under självreglering och interaktion med abiotiska faktorer, bibehåller autodynamiskt en livsmiljö som är lämplig för dess utveckling. Denna process begränsas av förändringar i den kosmiska och globala ekosfärskalan och förekommer i alla ekosystem och biosystem på planeten, som en kaskad av självreglering som når en global skala. Regeln om automatiskt underhåll av den globala livsmiljön följer av de biogeokemiska principerna i V.I. Vernadsky, reglerna för att bevara arters livsmiljöer, relativ intern konsistens och fungerar som en konstant för närvaron av konservativa mekanismer i biosfären och bekräftar samtidigt regeln om systemdynamisk komplementaritet. Den kosmiska inverkan på biosfären bevisas av lagen om brytning av kosmiska effekter: kosmiska faktorer, som påverkar biosfären och särskilt dess underavdelningar, är föremål för förändring av planetens ekosfär och därför, i termer av styrka och tid , manifestationer kan försvagas och förskjutas eller till och med helt förlora sin effekt. Generaliseringen här är viktig på grund av det faktum att det ofta finns ett flöde av synkrona effekter av solaktivitet och andra kosmiska faktorer på jordens ekosystem och de organismer som lever i den. Det bör noteras att många processer på jorden och i dess biosfär, även om de är föremål för inflytande från rymden, antas cykler av solaktivitet med intervaller på 1850, 600, 400, 178, 169, 88, 83, 33, 22, 16 , 11, 5 (11 ,1), 6,5 och 4,3 år, biosfären själv och dess indelningar behöver inte nödvändigtvis reagera med samma cyklicitet i alla fall. Biosfärsystemets kosmiska influenser kan blockeras helt eller delvis. Slutsats Ekosystem är den grundläggande funktionella enheten i ekologi, eftersom de inkluderar organismer och den livlösa miljön - komponenter som ömsesidigt påverkar varandras egenskaper och de nödvändiga förutsättningarna för att upprätthålla liv i den form som finns på jorden. Kombinationen av en specifik fysikalisk-kemisk miljö (biotop) med en gemenskap av levande organismer (biocenos) bildar ett ekosystem. Det finns mikroekosystem, mesoekosystem och det globala ekosystemet - biosfären. Ekosystemen är inte utspridda i oordning, tvärtom är de grupperade i ganska regelbundna zoner, både horisontellt (på latitud) och vertikalt (i höjd). Den primära energikällan för ekosystemen är solen. Solstrålningen som faller på jordens yta är 1,54 miljoner EJ per år. Det mesta av solenergin som når planetens yta omvandlas direkt till värme, värmande vatten eller jord, vilket i sin tur värmer luften. Ekosystem existerar på grund av icke-förorenande och nästan evig solenergi, vars mängd är relativt konstant och riklig. Solenergi på jorden orsakar två cykler av ämnen: stora, eller geologiska, och små, biologiska (biotiska). Båda cyklerna är ömsesidigt förbundna och representerar så att säga en enda process. Förekomsten av biogeokemiska cykler, eller biogeokemiska cykler, skapar möjlighet till självreglering (homeostas) av systemet, vilket ger ekosystemet stabilitet: en fantastisk konstanthet av den procentuella halten av olika grundämnen. Det finns ganska många kretslopp av ämnen. Den mest betydelsefulla cykeln på jorden i termer av överförda massor och energiförbrukning är den planetära hydrologiska cykeln eller vattencykeln. Det biotiska (biologiska) kretsloppet avser cirkulationen av ämnen mellan jord, växter, djur och mikroorganismer. Av alla biogeokemiska kretslopp är kolets kretslopp utan tvekan den mest intensiva. Det mesta av syret som producerades under geologiska epoker fanns inte kvar i atmosfären utan fixerades av litosfären i form av karbonater, sulfater, järnoxider etc. Denna massa är 590x1014 ton mot 39x1014 ton syre, som cirkulerar i biosfären i form av gas eller sulfater lösta i kontinentala och oceaniska vatten. Kväve är ett väsentligt biogent element, eftersom det är en del av proteiner och nukleinsyror. Kvävecykeln är en av de mest komplexa, eftersom den inkluderar både gas- och mineralfaser, och samtidigt de mest idealiska kretsloppen. Fosforcykeln i biosfären är förknippad med metaboliska processer hos växter och djur. Detta viktiga och nödvändiga element av protoplasma, som finns i landväxter och alger 0,01-0,1%, djur från 0,1% till flera procent, cirkulerar och förvandlas gradvis från organiska föreningar till fosfater, som återigen kan användas av växter. Det finns många gasformiga svavelföreningar, såsom svavelväte H2S och svaveldioxid SO2. Att upprätthålla den vitala aktiviteten hos organismer och cirkulationen av materia i ekosystemen, d.v.s. ekosystemens existens, beror på det konstanta energiflödet som är nödvändigt för alla organismer för deras vitala funktioner och självreproduktion. Till skillnad från ämnen som kontinuerligt cirkulerar genom olika block i ekosystemet, som alltid kan återanvändas och komma in i kretsloppet, kan energi användas en gång, det vill säga det finns ett linjärt flöde av energi genom ekosystemet. Inom ett ekosystem skapas energihaltiga ämnen av autotrofa organismer och fungerar som föda för heterotrofer. Matkopplingar är mekanismer för att överföra energi från en organism till en annan. Inom varje ekosystem har näringsnäten en väldefinierad struktur, som kännetecknas av arten och antalet organismer representerade på varje nivå i de olika näringskedjorna. Bildandet av ekosystem är en dynamisk process. I ekosystem sker ständigt förändringar i deras medlemmars tillstånd och vitala aktivitet och i förhållandet mellan befolkningar. De olika förändringar som sker i alla samhällen delas in i två huvudtyper: cykliska och progressiva. Succession är en naturlig, riktad process och de förändringar som sker i ett eller annat skede är karakteristiska för vilket samhälle som helst och är inte beroende av dess artsammansättning eller geografiska läge. Det globala ekosystemet är biosfären. Som noterats tidigare är biosfären uppdelad i geobiosfär, hydrobiosfär och aerobiosfär. Biosfären bildas i större utsträckning inte av yttre faktorer, utan av inre mönster. Biosfärens viktigaste egenskap är interaktionen mellan levande och icke-levande saker, vilket återspeglas i lagen om biogen migration av atomer av V.I. Vernadsky. Ekosystemen som finns på jorden är olika. Det finns mikroekosystem (till exempel stammen på ett ruttnande träd), mesoekosystem (skog, damm, etc.), makroekosystem (kontinent, hav, etc.) och den globala biosfären. Stora terrestra ekosystem kallas biomer. Varje biom innehåller ett antal mindre, sammanlänkade ekosystem. Det finns flera klassificeringar av ekosystem: vintergrön tropisk regnskog; öken: gräsbevuxen och buskig; tropiskt gräs och savann; tempererad stäpp; tempererad lövskog: boreala barrskogar. Tundra: arktisk och alpin. Typer av sötvattensekosystem: band (stilla vatten): sjöar, dammar, etc.; lotic (strömmande vatten): floder, bäckar, etc.; våtmarker: träsk och sumpiga skogar; Typer av marina ekosystem: öppet hav (pelagiskt); vatten på kontinentalsockeln (kustvatten); uppväxtområden (bördiga områden med produktivt fiske); flodmynningar (kustvikar, sund, flodmynningar, saltmarker etc.). Varje ekosystem har två huvudkomponenter: organismer och faktorer från deras icke-levande miljö. Helheten av organismer (växter, djur, mikrober) kallas biota i ett ekosystem. Sätten att interagera mellan olika kategorier av organismer är dess biotiska struktur. Solenergi på jorden orsakar två kretslopp av ämnen: en stor, eller geologisk, tydligast manifesterad i vattnets kretslopp och atmosfärscirkulation, och en liten, biologisk (biotisk), som utvecklas på basis av en stor och består av en kontinuerlig, cyklisk, men ojämn i tid och rum, och åtföljd av mer eller mindre betydande förluster i den naturliga omfördelningen av materia, energi och information inom ekologiska system på olika organisationsnivåer. Båda cyklerna är ömsesidigt förbundna och representerar så att säga en enda process. Samspelet mellan abiotiska faktorer och levande organismer i ekosystemet åtföljs av en kontinuerlig cirkulation av materia mellan biotopen och biocenosen i form av alternerande organiska och mineraliska föreningar. Utbytet av kemiska element mellan levande organismer och den oorganiska miljön, vars olika stadier sker inom ett ekosystem, kallas det biogeokemiska kretsloppet, eller biogeokemiskt kretslopp. Förekomsten av sådana cykler skapar möjlighet till självreglering (homeostas) av systemet, vilket ger ekosystemet stabilitet: en fantastisk konstanthet av procentandelen av olika element. Principen om ekosystemens funktion gäller här: anskaffning av resurser och omhändertagande av avfall sker inom ramen för alla elements kretslopp. Lista över begagnad litteratur 1.Bigon M.I. Ekologi, individer, befolkningar och samhällen. M.: 1989. - 290 sid. 2.Beysenova A.S., Shildebaev Zh.B., Sautbaeva G.Z. Ekologi. Almaty: "Gylym, 2001. - 201 sid. 3.Akimova T.A., Khaskin V.V. Ekologi. M.: UNITY Publishing House, 1998. - 233 sid. .Gorelov A.A. Ekologi. Föreläsningskurs. M.: "Center" 1997. - 280 sid. .Stadnitsky G.V., Rodionov A.I. Ekologi. M.: UNITY Publishing House, 1996. - 272 sid. .Sadanov A.K., Svanbaeva Z.S., Ekologi. Almaty: "Agrouniversity", 1999. - 197 s. .Shilov I.A. Ekologi. M.: Högre skola, 2000. - 348 sid. .Reimers N.F. Ekologi (Teori, lagar, regler, principer och hypoteser). M.: "Unga Ryssland", 1994. - 260 s. .Tsvetkova L.I., Alekseev M.I., Usenov B.P. och andra Ekologi. Lärobok för tekniska universitet. M.; ASV, St Petersburg: Khimizdat, 1999, 185 sid. .Girusov E.V. Ekologi och ekonomi för miljöledning. M.: Lag och lag, ENHET. 1998 - 232 sid. .Vronsky V.A. Tillämpad ekologi: Lärobok. Dra nytta av. Rostov-on-Don: 1995. - 197 sid. .Budyko M.I. Global ekologi. M.: Mysl, 1977.-248 sid. .Alekseenko V.A. Miljögeokemi. Lärobok. M.: Logos, 2000 - 410 sid. .Petrov K.M. Allmän ekologi för S-P. Handledning. "Kemi", 1997. - 218 sid. .Andersen J.M. Ekologi och miljövetenskap. Biosfär, ekosystem, människa. L., 1985. 376 sid. .Girenok F.I. Ekologi. Civilisation. Noosphere.M.: 191987. - 281 sid. .På jakt efter balans: ekologi i systemet för socialpolitiska prioriteringar M.: 1992. - 427 s. .Historia om samverkan mellan samhälle och natur: Faktorer och koncept. M.: ANSSSR 1990. .Stepanovskikh A.S. Tillämpad ekologi: Miljöskydd. Lärobok för universitet. M.: ENHET DANA. 2003. 751 sid. Liknande verk till - Ämneskretslopp och energiflöden i ekosystem
Termerna "materiaflöde" och "energiflöde" bör vara tydligt definierade. Materiens flöde är rörelsen i form av kemiska grundämnen och deras föreningar från producenter till nedbrytare (genom eller utan konsumenter). Energiflöde är överföring av energi i form av kemiska bindningar av organiska föreningar (mat) längs näringskedjor från en trofisk nivå till en annan (högre).
Det bör noteras att, till skillnad från ämnen som ständigt cirkulerar genom olika block i ekosystemet och alltid kan gå in i kretsloppet igen, kan den inkommande energin bara användas en gång.
Som ett universellt naturfenomen bestäms envägsflödet av energi av termodynamikens lagar. Enligt den första lagen kan energi flytta från en form (ljusets energi) till en annan (matens potentiella energi), men den skapas aldrig igen och försvinner inte spårlöst.
Termodynamikens andra lag säger att det inte kan finnas en enda process förknippad med omvandlingen av energi utan att förlora en del av den. Av denna anledning kan det inte ske någon omvandling med 100 % effektivitet, till exempel av mat till den substans som organismens kropp består av.
Således bestäms alla ekosystems funktion av ett konstant flöde av energi, vilket är nödvändigt för att alla organismer ska kunna behålla sin existens och självreproduktion.
Det finns också konkurrensförhållanden i ekosystemen. I denna aspekt är lagen om energimaximering (G. Odum - E. Odum) av stort intresse: i konkurrens med andra ekosystem överlever det som bäst bidrar till energiförsörjningen och använder sin maximala mängd på det mest effektiva sättet (sylt). Enligt lagen, för detta ändamål: 1) skapar ackumulatorer (lagringar) av högkvalitativ energi (till exempel fettreserver); 2) spenderar en viss mängd ackumulerad energi för att säkerställa tillgången på ny energi; 3) säkerställer cirkulationen av olika ämnen; 4) skapar regleringsmekanismer som stödjer systemets stabilitet och dess förmåga att anpassa sig till förändrade förhållanden; 5) etablerar utbyten med andra system som är nödvändiga för att möta behovet av speciella energislag.
Det är nödvändigt att betona en viktig omständighet: lagen om energimaximering är också giltig i förhållande till information, därför kan den (enligt N. F. Reimers) också betraktas som lagen om energi och informationsmaximering: det system som mest bidrar till att mottagning, produktion och effektiv användning av energi och information.
Låt oss notera att det maximala utbudet av ett ämne som sådant ännu inte garanterar systemets framgång i en konkurrensutsatt grupp av andra liknande system.
Det har tidigare noterats att starka matrelationer, eller en näringskedja, uppstår och etableras mellan biocenosens organismer. Den senare består av tre huvudlänkar: producenter, konsumenter och nedbrytare.
Näringskedjor som börjar med fotosyntetiska organismer kallas betes- (eller betes-)kedjor, och kedjor som börjar med döda växtmaterial, kadaver och djurexkrementer kallas detritalkedjor.
Placeringen av varje länk i näringskedjan kallas en trofisk nivå; det kännetecknas av olika intensiteter i flödet av ämnen och energi. Den första trofiska nivån är alltid producenter; den andra - växtätande konsumenter; den tredje - köttätare, som lever av växtätande former; fjärde nivån - konsumera andra köttätare, etc.
Det finns konsumenter av den första, andra, tredje och fjärde beställningen, som upptar olika nivåer i livsmedelskedjan (fig. 9).
Ris. 9.
Det är uppenbart att konsumenternas livsmedelsspecialisering spelar en stor roll i detta. Arter med ett brett näringsspektrum kan ingå i näringskedjor på olika trofiska nivåer. En persons kost inkluderar till exempel både vegetabilisk mat och kött från växtätare och köttätare. Därför agerar den i olika livsmedelskedjor som konsument av I-, II- eller III-beställningarna.
Eftersom energi går förlorad när energi överförs från en nivå till en annan kan kraftkedjan inte vara lång: den består vanligtvis av 4...6 länkar (tabell 1).
1. Typiska diagram över näringskedjor (enligt V. M. Ivonin, 1996)
Men sådana kedjor i sin rena form finns vanligtvis inte i naturen, eftersom samma art kan vara närvarande samtidigt i olika länkar. Detta beror på det faktum att det finns få monofager i naturen; oligofager och polyfager är mycket vanligare. Till exempel är rovdjur som livnär sig på olika växtätare och köttätare länkar i många kedjor. Som ett resultat av varje biocenos bildas komplex av näringskedjor evolutionärt, som representerar en enda helhet. Det är så kraftnät skapas, som är mycket komplexa.
Därmed kan vi dra slutsatsen att näringskedjan är den huvudsakliga kanalen för energiöverföring i samhället (mellan växter - producenter, djur - konsumenter och mikroorganismer - nedbrytare) (Fig. 10) Redan i diagrammet är det tydligt att idén om näringskedjor och trofiska nivåer är snarare en abstraktion. En linjär kedja med tydligt separerade nivåer kan skapas i laboratoriet. Men i naturen finns det verkligen trofiska nätverk där många populationer tillhör flera trofiska nivåer samtidigt. Samma organism konsumerar både djur och växter, rovdjuret kan livnära sig på konsumenter av första och andra ordningen, många djur äter både levande och döda växter.
På grund av komplexiteten i trofiska förhållanden har förlusten av en art ofta nästan ingen effekt på samhället. Andra "användare" börjar konsumera maten från den utdöda arten,

Ris. 10.
arterna som livnär sig på den hittar nya källor till mat: i allmänhet upprätthålls balansen i samhället.
Energin som tas upp av producenter flödar genom näringskedjorna och förbrukas gradvis. I slutet av en näringskedja är mängden energi alltid mindre än i början. Under fotosyntesen binder växter i genomsnitt bara cirka 1 % av solenergin som träffar dem. Ett djur som har ätit en växt smälter inte en del av maten och utsöndrar den i form av exkrementer. Vanligtvis smälts 20...60% av växtfödan; Den absorberade energin används för att upprätthålla djurets liv. Funktionen hos celler och organ åtföljs av frigöring av värme, det vill säga, därigenom försvinner en betydande del av matenergin snart i miljön. En relativt liten del av maten används för att bygga ny vävnad och skapa fettreserver. Därefter får ett rovdjur som har ätit en växtätare och representerar den tredje trofiska nivån endast den energin från den energi som ackumuleras av växten som hålls kvar i kroppen av sitt bytesdjur (andra nivån) i form av en ökning av biomassan.
Det är känt att i varje steg under överföringen av materia och energi i näringskedjan går cirka 90 % av energin förlorad och endast cirka en tiondel av den går till nästa konsument, dvs. överföringen av energi i livsmedelsförbindelserna av organismer lyder "tioprocentsregeln" (Lindemanns princip ). Till exempel är mängden energi som når tertiära köttätare (femte trofisk nivå) bara cirka 10 -4 av den energi som absorberas av producenter. Detta förklarar det begränsade antalet (5...6) länkar (nivåer) i näringskedjan, oavsett komplexiteten i artsammansättningen av biocenosen.

Ris. elva.
Med tanke på energiflödet i ekosystemen är det också lätt att förstå varför biomassan minskar när den trofiska nivån ökar. Här manifesteras den tredje grundläggande principen för ekosystemens funktion: ju större biomassa en befolkning har, desto lägre bör den trofiska nivån vara, eller på annat sätt: i slutet av långa näringskedjor kan det inte finnas en stor biomassa.
De tre grundläggande principerna för ekosystems funktion som listas ovan - näringsämnenas kretslopp, solenergiflödet och minskningen av biomassa med ökande trofisk nivå - kan presenteras i form av ett generellt diagram (fig. 11). Om vi ordnar organismer efter deras näringsförhållanden, och för var och en av dem indikerar "input" och "output" av energi och näringsämnen, blir det uppenbart att näringsämnen kontinuerligt återvinns inom ekosystemet, och energiflödet passerar genom det.
Att upprätthålla den vitala aktiviteten hos organismer och cirkulationen av materia i ekosystemen, d.v.s. ekosystemens existens, beror på det konstanta energiflödet som är nödvändigt för alla organismer för deras vitala funktioner och självreproduktion.
Till skillnad från ämnen som kontinuerligt cirkulerar genom olika block i ekosystemet, som alltid kan återanvändas och komma in i kretsloppet, kan energi endast användas en gång, det vill säga det finns ett linjärt flöde av energi genom ekosystemet.
Envägsinflödet av energi som ett universellt naturfenomen uppstår som ett resultat av termodynamikens lagar. Första lagen säger att energi kan omvandlas från en form (som ljus) till en annan (som matens potentiella energi), men kan inte skapas eller förstöras. Andra lagen anger att det inte kan finnas en enda process förknippad med omvandlingen av energi utan att förlora en del av den. En viss mängd energi i sådana transformationer försvinner till otillgänglig termisk energi och går därför förlorad. Därför kan det inte ske omvandlingar av till exempel livsmedelsämnen till det ämne som utgör organismens kropp, vilket sker med 100 procent effektivitet.
Således är levande organismer energiomvandlare. Och varje gång energi omvandlas går en del av den förlorad i form av värme. I slutändan försvinner all energi som kommer in i den biotiska cykeln i ett ekosystem som värme. Levande organismer använder faktiskt inte värme som energikälla för att utföra arbete – de använder ljus och kemisk energi.
Ekologiska pyramider. Inom varje ekosystem har näringsnäten en väldefinierad struktur, som kännetecknas av arten och antalet organismer representerade på varje nivå i de olika näringskedjorna. För att studera sambanden mellan organismer i ett ekosystem och för att avbilda dem grafiskt använder de sig vanligtvis av ekologiska pyramider snarare än diagram i näringsväven. Ekologiska pyramider uttrycker den trofiska strukturen i ett ekosystem i geometrisk form. De är konstruerade i form av rektanglar med samma bredd, men längden på rektanglarna måste vara proportionell mot värdet på objektet som mäts. Härifrån kan du få pyramider av siffror, biomassa och energi.
Ekologiska pyramider återspeglar de grundläggande egenskaperna hos varje biocenos när de visar dess trofiska struktur:
Deras höjd är proportionell mot längden av näringskedjan i fråga, d.v.s. antalet trofiska nivåer den innehåller;
Deras form återspeglar mer eller mindre effektiviteten av energiomvandlingar under övergången från en nivå till en annan.
Pyramider av siffror. De representerar den enklaste approximationen till studiet av den trofiska strukturen i ett ekosystem. I detta fall räknas först antalet organismer i ett givet territorium, grupperas efter trofiska nivåer och presenteras i form av en rektangel, vars längd (eller area) är proportionell mot antalet organismer som lever i ett givet område ( eller i en given volym, om det är ett akvatiskt ekosystem). En grundregel har fastställts att det i alla miljöer finns fler växter än djur, fler växtätare än köttätare, fler insekter än fåglar, etc.
Befolkningspyramider återspeglar tätheten av organismer på varje trofisk nivå. Det finns stor mångfald i konstruktionen av olika befolkningspyramider. Ofta är de upp och ner.
Till exempel finns det i en skog betydligt färre träd (primärproducenter) än insekter (växtätare).
Biomassa pyramid.Återspeglar mer fullständigt födoförhållandena i ekosystemet, eftersom det tar hänsyn till den totala massan av organismer (biomassa) varje trofisk nivå. Rektanglarna i biomassapyramiderna representerar massan av organismer på varje trofisk nivå per ytenhet eller volym. Formen på biomassapyramiden liknar ofta formen på befolkningspyramiden. En minskning av biomassa vid varje trofisk nivå är karakteristisk.
Pyramider av biomassa, såväl som siffror, kan inte bara vara raka utan också inverterade. Inverterade pyramider av biomassa är karakteristiska för akvatiska ekosystem, där primärproducenter, såsom växtplanktonalger, delar sig mycket snabbt, och deras konsumenter - zooplanktoniska kräftdjur - är mycket större men har en lång reproduktionscykel. I synnerhet gäller detta sötvattensmiljöer, där primär produktivitet tillhandahålls av mikroskopiska organismer vars ämnesomsättning ökar, dvs biomassan är låg, produktiviteten är hög.
Pyramid av energi. Det mest grundläggande sättet att visa kopplingar mellan organismer på olika trofiska nivåer är genom energipyramider. De representerar energiomvandlingseffektiviteten och produktiviteten i näringskedjorna och är konstruerade genom att räkna mängden energi (kcal) som ackumuleras per ytenhet per tidsenhet och som används av organismer på varje trofisk nivå. Det är alltså relativt enkelt att bestämma mängden energi som lagras i biomassa, men det är svårare att uppskatta den totala mängden energi som absorberas på varje trofisk nivå. Efter att ha konstruerat en graf (fig. 12.28) kan vi konstatera att förstörare, vars betydelse verkar liten i biomassapyramiden och vice versa i befolkningspyramiden; ta emot en betydande del av energin som passerar genom ekosystemet. Dessutom finns bara en del av all denna energi kvar i organismer på varje trofisk nivå i ekosystemet och lagras i biomassa; resten används för att tillfredsställa de metaboliska behoven hos levande varelser: upprätthålla existens, tillväxt, reproduktion. Djur förbrukar också en betydande mängd energi för muskelarbete.
R. Lindeman formulerades första gången 1942 energipyramidlag, som i läroböcker ofta kallas "lagen om 10%". Enligt denna lag från en trofisk nivå av den ekologiska pyramiden I genomsnitt går inte mer än 10% av energin till en annan nivå.
Konsumenter fungerar som en hanterande och stabiliserande länk i ekosystemet. Konsumenter genererar ett spektrum av mångfald i cenosen, vilket förhindrar dominanternas monopol. Regel för kontrollvärde för konsumenter kan med rätta anses vara ganska grundläggande. Enligt cybernetiska synsätt bör kontrollsystemet vara mer komplext till sin struktur än det kontrollerade, då blir orsaken till mångfalden av konsumenttyper tydlig. Konsumenternas kontrollerande betydelse har också en energisk grund. Energiflödet genom en eller annan trofisk nivå kan inte absolut bestämmas av tillgången på mat i den underliggande trofiska nivån. Som bekant finns det alltid en tillräcklig "reserv" kvar, eftersom en fullständig förstörelse av livsmedel skulle leda till konsumenternas död. Dessa allmänna mönster observeras inom ramen för befolkningsprocesser, samhällen, nivåer av den ekologiska pyramiden och biocenoser som helhet.
För att använda presentationsförhandsvisningar, skapa ett Google-konto och logga in på det: https://accounts.google.com
Bildtexter:
Materias och energins kretslopp i naturen
Ämnescykeln är de återkommande processerna för omvandling och rörelse av materia i naturen, som är mer eller mindre cykliska. Alla ämnen på vår planet är i cirkulationsprocessen. Det finns två huvudcykler i naturen: Stor (geologisk) Liten (biogeokemisk)
Den stora cykeln av substanser Den stora cykeln varar i miljontals år och orsakas av samspelet mellan solenergi och jordens djupa energi. Förknippas med geologiska processer, bildning och förstörelse av stenar och efterföljande förflyttning av förstörelseprodukter.
Litet kretslopp av ämnen Det lilla kretsloppet (biogeokemiskt) sker inom biosfären, på biocenosnivå. Dess väsen är bildandet av levande materia från oorganiska föreningar under fotosyntesprocessen och omvandlingen av organiskt material under nedbrytning tillbaka till oorganiska föreningar. Biogeokemiska cykler – Vernadsky V.I.
Vattencykel Tr avrinning inf Vattenavdunstning Ångkondensation Nederbörd avrinning Transpiration infiltration
Transpiration är processen för vatten som rör sig genom en växt och dess avdunstning genom växtens yttre organ, såsom löv, stjälkar och blommor. Vatten är nödvändigt för växtlivet, men endast en liten del av vattnet som tillförs genom rötterna används direkt för tillväxt och ämnesomsättning.
Vattnets kretslopp
Vattnets kretslopp Det mesta av vattnet är koncentrerat i haven. Vatten som avdunstar från deras yta försörjer naturliga och konstgjorda markekosystem. Ju närmare havet ett område är, desto mer nederbörd finns det. Landet returnerar ständigt vatten till havet: en del av fukten avdunstar, mest aktivt i skogar, en del samlas upp av floder: de får regn och smältvatten. Utbytet av fukt mellan hav och land kräver mycket höga energikostnader: cirka 30 % av solenergin som når jorden går åt till detta.
Människans inflytande på vattnets kretslopp Vattnets kretslopp i biosfären före civilisationens utveckling var i jämvikt, d.v.s. havet fick lika mycket vatten från floderna som det konsumerade genom avdunstning. Med utvecklingen av civilisationen började denna cykel att störas. Framför allt avdunstar skogarna mindre och mindre vatten eftersom... deras yta minskar, och jordytan, tvärtom, ökar, eftersom Arealen med bevattnad jordbruksmark ökar. landar. Floderna i de södra regionerna blev grunda. Vatten avdunstar värre från havets yta, eftersom... en betydande del av den är täckt med en film av olja. Allt detta försämrar vattentillförseln till biosfären.
Torkan blir allt vanligare och fickor med miljökatastrofer växer fram. Till exempel har en katastrofal torka varat i mer än 35 år i Afrika, i Sahel-zonen - en halvökenregion som skiljer Sahara från de nordliga länderna på kontinenten. Färskvatten som återgår till havet och andra vattenförekomster från land är ofta förorenat. Vattnet i många ryska floder har blivit praktiskt taget olämpligt att dricka. Andelen sötvatten tillgängligt för levande organismer är ganska liten, så det måste användas sparsamt och inte förorenas! Var fjärde person på planeten saknar rent dricksvatten. I många delar av världen finns det inte tillräckligt med vatten för industriell produktion och bevattning.
Olika komponenter i hydrosfären deltar i vattnets kretslopp på olika sätt och med olika hastigheter. För fullständig förnyelse av vatten i glaciärer tar det 8000 år, grundvatten - 5000 år, hav - 3000 år, mark - 1 år. Atmosfärisk ånga och flodvatten förnyas helt på 10–12 dagar. Vattnets kretslopp i naturen tar cirka 1 miljon år.
Syrekretslopp Syre är en del av de vanligaste elementen i biosfären. Syrehalten i atmosfären är nästan 21 %. Syre är en del av vattenmolekyler och levande organismer (proteiner, fetter, kolhydrater, nukleinsyror). Syre produceras av producenter (gröna växter). Ozon spelar en viktig roll i syrecykeln. Ozonskiktet ligger på en höjd av 20-30 km över havet. Syrehalten i atmosfären påverkas av 2 huvudprocesser: 1) fotosyntes 2) nedbrytning av organiskt material, under vilken det förbrukas.
Syrecykeln är en långsam process. Det tar cirka 2000 år innan allt syre i atmosfären är helt förnyat. Som jämförelse: fullständig förnyelse av koldioxid i atmosfären sker på cirka 3 år. Syre används för andning av de flesta levande organismer. Syre används vid förbränning av bränsle i förbränningsmotorer, i ugnar i termiska kraftverk, i flygplan och raketmotorer, etc. Ytterligare antropogen konsumtion kan rubba balansen i syrecykeln. Än så länge kompenserar biosfären för mänskligt ingripande: förluster kompenseras av gröna växter. Med ytterligare minskning av skogsarealen och förbränning av allt mer bränsle kommer syrehalten i atmosfären att börja minska.
DET ÄR VIKTIGT!!! När syrehalten i luften minskar till 16%, förvärras en persons hälsa (hjärtat lider särskilt), till 7%, en person förlorar medvetandet och till 3% inträffar döden.
Kolets kretslopp
Kolets kretslopp Kol är grunden för organiska föreningar, det är en del av alla levande organismer i form av proteiner, fetter och kolhydrater. Kol kommer in i atmosfären i form av koldioxid. I atmosfären, där huvuddelen av koldioxiden är koncentrerad, sker ständigt ett utbyte: växter absorberar koldioxid under fotosyntesen, och alla organismer släpper ut den under andning. Upp till 50 % av kolet i form av CO 2 återförs till atmosfären av nedbrytare - markmikroorganismer. Kol lämnar kretsloppet som kalciumkarbonat.
Människans påverkan på kolets kretslopp Teknogen mänsklig aktivitet stör den naturliga balansen i kolets kretslopp: 1) vid förbränning av organiskt bränsle släpps cirka 6 miljarder ton CO 2 ut i atmosfären årligen: a) Elproduktion vid värmekraftverk b. ) Bilavgaser 2) förstörelse av skog. Under de senaste 100 åren har halten koldioxid i atmosfären ökat stadigt och snabbt. Koldioxid + metan + vattenånga + ozon + kväveoxider = växthusgas. Resultatet är växthuseffekten – global uppvärmning, som kan leda till storskaliga naturkatastrofer.
Kvävekretslopp I fri form är kväve en komponent i luft - 78%. Kväve är ett av de viktigaste elementen för organismers liv. Kväve är en del av alla proteiner. Kvävemolekylen är mycket stark, varför de flesta organismer inte kan ta upp atmosfäriskt kväve. Kväve tas upp av levande organismer endast i form av föreningar med väte och syre. Fixering av kväve i kemiska föreningar sker som ett resultat av vulkanisk aktivitet och åskväder, men mest som ett resultat av aktiviteten hos mikroorganismer - kvävefixare (kvävefixerande bakterier och blågröna alger).
Kväve kommer in i växternas rötter i form av nitrater, som används för syntes av organiskt material (proteiner). Djur konsumerar kväve genom växt- eller djurfoder. Kväve återförs till atmosfären genom nedbrytning av dött organiskt material. Jordbakterier sönderdelar proteiner till oorganiska ämnen - gaser - ammoniak, kväveoxider, som kommer in i atmosfären. Kväve som kommer in i vattenkroppar passerar också genom näringskedjan växt-djur-mikroorganismer och återgår till atmosfären.
Människans påverkan på kvävekretsloppet Teknogen mänsklig aktivitet stör den naturliga balansen i kvävekretsloppet. Vid plöjning av mark minskar aktiviteten hos mikroorganismer - kvävefixerare nästan 5 gånger, därför minskar kvävehalten i jorden, vilket leder till en minskning av jordens bördighet. Därför introducerar människor överskott av nitrater i jorden, som ingår i mineralgödsel. En stor mängd kväveoxider kommer in i atmosfären under förbränning och bearbetning av gas, olja och kol och faller i form av surt regn. Att återställa det naturliga kvävekretsloppet är möjligt genom att minska produktionen av kvävegödselmedel, minska industriella utsläpp av kväveoxider till atmosfären, etc.
Fosforcykel
Till skillnad från vatten-, kol-, kväve- och syrekretsloppen, som är slutna, är fosforcykeln öppen, eftersom Fosfor bildar inte flyktiga föreningar som släpps ut i atmosfären. Fosfor finns i bergarter, varifrån det kommer in i ekosystemen under den naturliga förstörelsen av bergarter eller när fosforgödselmedel appliceras på åkrar. Växter absorberar oorganiska fosforföreningar, och djur som livnär sig på dessa växter samlar fosfor i sina vävnader. Efter nedbrytningen av djurens och växternas döda kroppar är inte all fosfor involverad i kretsloppet. En del av det sköljs ur jorden till vattenmassor (floder, sjöar, hav) och lägger sig till botten. Fosfor återvänder till land i små mängder med fisk som fångas av människor.
Människans påverkan på fosforns kretslopp Överföringen av fosfor från land till havet har ökat markant under mänsklig påverkan. När skogar förstörs och jordar plöjs ökar volymen av ytvattenflödet, och dessutom kommer fosforgödsel som appliceras på floder och sjöar från åkrar. Eftersom fosforreserverna på land är begränsade, och dess återföring från havet är svår, kan det i framtiden bli brist på fosfor i jordbruket, vilket kommer att orsaka minskade skördar (främst spannmålsgrödor).