Protok energije u prirodi. Kruženje materije i energije u prirodi
Svaki život zahtijeva stalni protok energije i materije. Energija se troši na odvijanje osnovnih životnih reakcija, materija se koristi za izgradnju tijela organizama. Postojanje prirodnih ekosustava prate složeni procesi izmjene materijala i energije između žive i nežive prirode. Ti su procesi vrlo važni i ovise ne samo o sastavu biotskih zajednica, već io njihovom fizičkom okolišu.
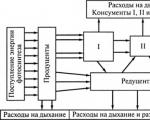
Protok energije u zajednici– ovo je njegov prijelaz iz organizama jedne razine u drugu u obliku kemijskih veza organskih spojeva (hrana).
Tijek (krug) tvari je kretanje tvari u obliku kemijskih elemenata i njihovih spojeva od proizvođača do razlagača i zatim (kroz kemijske reakcije koje se odvijaju bez sudjelovanja živih organizama) ponovno do proizvođača.
Kruženje tvari i protok energije nisu identični pojmovi, iako se za mjerenje kretanja tvari često koriste različiti energetski ekvivalenti (kalorije, kilokalorije, džuli). To se djelomično objašnjava činjenicom da se na svim trofičkim razinama, s izuzetkom prve, energija potrebna za život organizama prenosi u obliku tvari konzumirane hrane. Samo biljke (proizvođači) mogu izravno koristiti energiju zračenja Sunca za svoju životnu aktivnost.
Strogo mjerenje tvari koja kruži u ekosustavu može se dobiti uzimajući u obzir kruženje pojedinih kemijskih elemenata, prvenstveno onih koji su glavni gradivni materijal za citoplazmu biljnih i životinjskih stanica.
Za razliku od tvari koje kontinuirano cirkuliraju kroz različite blokove ekosustava i uvijek mogu ponovno ući u ciklus, energija se u tijelu može iskoristiti samo jednom.
Prema zakonima fizike, energija može prijeći iz jednog oblika (kao što je svjetlosna energija) u drugi (kao što je potencijalna energija hrane), ali se nikada više ne stvara niti uništava. Ne može postojati niti jedan proces povezan s transformacijom energije bez gubitka dijela energije. U njegovim se transformacijama određena količina energije raspršuje u obliku topline i stoga se gubi. Iz tog razloga ne može doći do transformacija, primjerice, tvari iz hrane u tvari koje čine tijelo organizma, a koje se događaju sa stopostotnom učinkovitošću.
Postojanje svih ekosustava ovisi o stalnom protoku energije koja je neophodna svim organizmima za održavanje vitalnih funkcija i samoreprodukciju.
Samo oko polovice sunčevog toka koji pada na zelene biljke apsorbiraju fotosintetski elementi, a samo mali dio apsorbirane energije (od 1/100 do 1/20 dijela) pohranjuje se u obliku energije potrebne za aktivnost biljnih tkiva.
Kako se čovjek udaljava od primarnog proizvođača, brzina protoka energije (to jest, količina energije, izražena u energetskim jedinicama, prenesena s jedne trofičke razine na drugu) naglo opada.
Pad količine energije tijekom prijelaza s jedne trofičke razine na višu određuje broj samih tih razina. Procjenjuje se da svaka trofična razina prima samo oko 10% (ili malo više) energije prethodne razine. Stoga ukupni broj trofičkih razina rijetko prelazi 3-4.
Omjer žive tvari na različitim trofičkim razinama općenito se pokorava istom pravilu kao i omjer ulazne energije: što je viša razina, to je manja ukupna biomasa i broj organizama koji je čine.
Omjer brojnosti različitih skupina organizama daje predodžbu o stabilnosti zajednice, jer su biomasa i brojnost pojedinih populacija ujedno i pokazatelj životnog prostora za organizme ove i drugih vrsta. . Na primjer, broj stabala u šumi ne određuje samo ukupnu zalihu biomase i energije sadržane u njima, već i mikroklimu, kao i broj skloništa za mnoge kukce i ptice.
Piramide brojeva mogu se obrnuti. To se događa kada je stopa reprodukcije populacije plijena visoka, pa čak i uz nisku biomasu, takva populacija može biti dovoljan izvor hrane za predatore koji imaju veću biomasu, ali nisku stopu reprodukcije. Na primjer, mnogi kukci mogu živjeti i hraniti se na jednom stablu (obrnuta populacijska piramida). Obrnuta piramida biomase karakteristična je za vodene ekosustave, gdje se primarni proizvođači (fitoplanktonske alge) vrlo brzo dijele i brojčano razmnožavaju, a njihovi konzumenti (zooplanktonski rakovi) mnogo su veći, ali imaju dug ciklus razmnožavanja.
Pašni i detritični lanci
Energija može teći kroz zajednicu na mnogo različitih načina. Predstavlja prehrambeni lanac svih potrošača(potrošački sustav) uz dodatak još dvije poveznice: ovo mrtve organske tvari I decomposer hranidbeni lanac(sustav redukcije).
Protok energije od biljaka kroz biljojede(nazivaju se ispaša), nazvan hranidbeni lanac travnjaka.
Ostaci organizama koje konzumiraju, a koje potrošači ne koriste, nadoknađuju mrtvu organsku tvar. Sastoji se od izmeta koji sadrži dio neprobavljene hrane, kao i životinjskih leševa, ostataka vegetacije (lišće, grane, alge) i naziva se detritus.
Tok energije koji potječe iz mrtve organske tvari i prolazi kroz sustav koji se raspada naziva se detritivnim hranidbenim lancem.
Uz sličnosti, postoje duboke razlike u funkcioniranju hranidbenih lanaca pašnjaka i detritusa. Sastoji se u tome što u u potrošačkom sustavu gube se izmet i mrtvi organizmi, a u redukcijskom– Ne.
Prije ili kasnije, energiju sadržanu u mrtvoj organskoj tvari razlagači će u potpunosti iskoristiti i raspršiti je kao toplinu kroz disanje, čak i ako to zahtijeva prolazak kroz sustav razlagača nekoliko puta. Jedina iznimka su oni slučajevi kada su lokalni abiotski uvjeti vrlo nepovoljni za proces razgradnje (visoka vlažnost, permafrost). U tim slučajevima nakupljaju se naslage nepotpuno prerađenih, energetski visoko intenzivnih tvari, koje se tijekom vremena i pod odgovarajućim uvjetima pretvaraju u zapaljive organske fosile - naftu, ugljen, treset.
Kruženje tvari u ekosustavu
Cjelovitost prirodnih ekosustava posebno se jasno očituje kada se uzmu u obzir tokovi tvari koji u njima kruže. Tvar se može prenositi u zatvorenim ciklusima (kruženjima), opetovano kružeći između organizama i okoliša.
Kružna kretanja (zemljom, zrakom, vodom) kemijskih elemenata (odnosno tvari) nazivaju se biogeokemijskim ciklusima ili kruženjima.
Elementi i otopljene soli potrebne za život konvencionalno se nazivaju hranjivim tvarima(davanje života) ili hranjivim tvarima. Među biogenim elementima razlikuju se dvije skupine: makrotrofne tvari i mikrotrofne tvari.
Makrotrofne tvari obuhvaćaju elemente koji čine kemijsku osnovu tkiva živih organizama. Tu spadaju: ugljik, vodik, kisik, dušik, fosfor, kalij, kalcij, magnezij, sumpor.
Mikrotrofne tvari uključuju elemente i njihove spojeve, također vrlo važne za postojanje živih sustava, ali u iznimno malim količinama. Takve se tvari često nazivaju mikroelemenata. To su željezo, mangan, bakar, cink, bor, natrij, molibden, klor, vanadij i kobalt. Iako su mikrotrofni elementi neophodni organizmima u vrlo malim količinama, njihov nedostatak može ozbiljno ograničiti produktivnost.
Kruženje hranjivih tvari obično je popraćeno njihovim kemijskim pretvorbama. Nitratni dušik, na primjer, može se pretvoriti u proteinski dušik, zatim pretvoriti u ureu, pretvoriti u amonijak i ponovno sintetizirati u nitratni oblik pod utjecajem mikroorganizama. U procese denitrifikacije i fiksacije dušika uključeni su različiti mehanizmi, kako biološki tako i kemijski.
Rezerve hranjivih tvari su promjenjive. Proces izdvajanja nekih od njih kao žive biomase smanjuje količinu koja ostaje u abiotskom okolišu. A da se biljke i drugi organizmi na kraju ne razgrade, zalihe hranjivih tvari bile bi iscrpljene i život na Zemlji bi prestao. Iz ovoga možemo zaključiti da aktivnost heterotrofa, prvenstveno organizama koji funkcioniraju u detritalnim lancima,– odlučujući čimbenik u održavanju ciklusa hranjivih tvari i formiranju proizvoda.
Pogledajmo neke numeričke podatke koji pokazuju razmjere prijenosa tvari, okrećući se biogeokemijskom ciklusu ugljika. Prirodni izvor ugljika koji biljke koriste za sintezu organske tvari je ugljikov dioksid koji je dio atmosfere ili je otopljen u vodi. Tijekom fotosinteze ugljikov dioksid (ugljični dioksid) se pretvara u organsku tvar koja služi kao hrana životinjama. Disanje, fermentacija i izgaranje goriva vraćaju ugljični dioksid u atmosferu.
Rezerve ugljika u atmosferi našeg planeta procjenjuju se na 700 milijardi tona, u hidrosferi - na 50 000 milijardi tona Prema godišnjim izračunima, kao rezultat fotosinteze, povećanje biljne mase na kopnu iu vodi iznosi 30 milijardi tona i 150 milijardi tona, odnosno Ciklus ugljika traje oko 300 godina.
Drugi primjer je ciklus fosfora. Glavne rezerve fosfora sadrže razne stijene, koje postupno (kao rezultat razaranja i erozije) otpuštaju svoje fosfate u kopnene ekosustave. Fosfate konzumiraju biljke i koriste ih za sintezu organskih tvari. Kada mikroorganizmi razgrade lešine životinja, fosfati se vraćaju u tlo, a zatim ih ponovno koriste biljke. Osim toga, dio fosfata vodotoci odnesu u more. Time se osigurava razvoj fitoplanktona i svih hranidbenih lanaca koji o njemu ovise. Dio fosfora sadržanog u morskoj vodi može se vratiti na kopno u obliku guana.
ekosustav biosphere energetski ciklus
Uvod
Pojam i struktura ekosustava
1 Pojam ekosustava
2 Klasifikacija ekosustava
3 Zoniranje makroekosustava
4 Struktura ekosustava
Čimbenici koji osiguravaju cjelovitost ekosustava
1 Ciklusi tvari
2 Protok energije u ekosustavima
3 Dinamički procesi koji osiguravaju cjelovitost i održivost ekosustava
4 Biosfera kao globalni ekosustav koji osigurava cjelovitost i održivost ekosustava
Zaključak
Uvod
Ekosustav je svaka zbirka organizama i anorganskih komponenti u kojima se može dogoditi kruženje tvari. Prema N.F. Reimers (1990), ekosustav je svaka zajednica živih bića i njihova staništa, ujedinjena u jedinstvenu funkcionalnu cjelinu, nastala na temelju međuovisnosti i uzročno-posljedičnih veza koje postoje između pojedinih komponenti okoliša. A. Tansley (1935) predložio je sljedeći odnos:
Postoje mikroekosustavi, mezoekosustavi, makroekosustavi i globalna biosfera. Veliki kopneni ekosustavi nazivaju se biomi. Ekosustavi nisu raštrkani u neredu, naprotiv, grupirani su u prilično pravilne zone, vodoravno (po širini) i okomito (po visini).
Od ekvatora do polova vidljiva je određena simetrija u rasporedu bioma različitih hemisfera: tropske kišne šume, pustinje, stepe, umjerene šume, crnogorične šume, tajga.
Svaki ekosustav ima dvije glavne komponente: organizme i čimbenike iz njihove nežive okoline. Ukupnost organizama (biljke, životinje, mikrobi) naziva se biota ekosustava. S gledišta trofičke strukture (od grčkog trophe - prehrana), ekosustav se može podijeliti u dva sloja: gornji, donji.
Kao dio ekosustava razlikuju se sljedeće komponente: anorganski, organski spojevi, okoliš zraka, vode i supstrata, proizvođači, autotrofni organizmi, potrošači ili fagotrofi, razlagači i detritivori.
Sunčeva energija na Zemlji uzrokuje dva ciklusa tvari: veliki, ili geološki, i mali, biološki (biotički).
U ekosustavu postoje ciklusi tvari. Najviše su proučavani: ciklusi ugljika, kisika, dušika, fosfora, sumpora itd.
Održavanje životne aktivnosti organizama i kruženje tvari u ekosustavima, odnosno postojanje ekosustava, ovisi o stalnom dotoku sunčeve energije.
U ekosustavima se stalno događaju promjene u stanju i životnoj aktivnosti njihovih članova te u omjeru populacija. Različite promjene koje se događaju u bilo kojoj zajednici mogu se podijeliti u dvije glavne vrste: cikličke i progresivne.
Globalni ekosustav je biosfera koju karakterizira cjelovitost i stabilnost ekosustava.
1. Pojam i struktura ekosustava
1 Pojam ekosustava
Živi organizmi i njihov neživi (abiotski) okoliš međusobno su neraskidivo povezani iu stalnoj su interakciji. Svaka jedinica (biosustav) ekosustava je ekološki sustav. Ekološki sustav ili ekosustav osnovna je funkcionalna cjelina u ekologiji, budući da uključuje organizme i neživi okoliš – komponente koje međusobno utječu na svojstva i nužne uvjete za održanje života u obliku kakav postoji na Zemlji. Pojam "ekosustav" prvi je predložio 1935. godine engleski ekolog A. Tansley. Trenutno se široko koristi sljedeća definicija ekosustava. Ekosustav je svaka zbirka organizama i anorganskih komponenti u kojima se može dogoditi kruženje tvari. Prema N.F. Reimers (1990), ekosustav je svaka zajednica živih bića i njihova staništa, ujedinjena u jedinstvenu funkcionalnu cjelinu, nastala na temelju međuovisnosti i uzročno-posljedičnih veza koje postoje između pojedinih komponenti okoliša. Treba naglasiti da spoj specifičnog fizikalno-kemijskog okoliša (biotopa) sa zajednicom živih organizama (biocenoza) čini ekosustav. A. Tansley (1935) predložio je sljedeći odnos:
Ekosustav = Biotop + Biocenoza
Prema definiciji V.N. Sukačev, biogeocenoza je „skup homogenih prirodnih pojava (atmosfere, stijena, tla i hidroloških uvjeta) na određenom dijelu zemljine površine, koji ima svoju posebnu specifičnost međudjelovanja ovih komponenti koje ga čine i određenu vrstu razmjenu materije i energije između sebe i drugih prirodnih pojava i predstavljaju unutarnje proturječno dijalektičko jedinstvo, u stalnom kretanju i razvoju."
Uz koncepte ekosustava A. Tensley i biogeocenoze V.N. Sukačev je formulirao pravilo F. Evansa (1956.), koji je predložio korištenje pojma "ekosustav" apsolutno "bezdimenzionalno" za označavanje bilo kojeg supraorganizmskog živog sustava u interakciji s okolišem. Međutim, mnogi autori su pojmu "ekosustav" dali značenje biogeocenoze, tj. elementarni ekosustav, a ujedno i viši u hijerarhiji suprabiogeocenotskih tvorevina do ekosustava biosfere.
2 Klasifikacija ekosustava
Ekosustavi koji postoje na Zemlji su raznoliki. Postoje mikroekosustavi (na primjer, deblo trulog drveta), mezoekosustavi (šuma, ribnjak itd.), makroekosustavi (kontinent, ocean itd.) i globalna biosfera.
Veliki kopneni ekosustavi nazivaju se biomi. Svaki biom sadrži niz manjih, međusobno povezanih ekosustava. Postoji nekoliko klasifikacija ekosustava:
Zimzelena tropska kišna šuma
Pustinja: Trava i grmlje Chaparral - područja s kišnim zimama i suhim ljetima
Tropski Graslenz i Savannah
Umjerena stepa
Listopadna šuma umjerenog pojasa
Borealne crnogorične šume
Tundra: arktička i alpska
Tipovi slatkovodnih ekosustava:
Ribbon (mirne vode): jezera, bare itd.
Lotic (tekuće vode): rijeke, potoci itd.
Močvare: močvare i močvarne šume
Vrste morskih ekosustava
Otvoreni ocean (pelagikal)
Vode kontinentalnog pojasa (obalne vode)
Područja uzdizanja (plodna područja s produktivnim ribolovom)
Estuariji (obalni zaljevi, tjesnaci, riječna ušća, slane močvare itd.). Kopneni biomi ovdje se razlikuju po prirodnim ili izvornim značajkama vegetacije, a tipovi vodenih ekosustava razlikuju se po geološkim i fizičkim značajkama. Navedeni glavni tipovi ekosustava predstavljaju okoliš u kojem se razvila ljudska civilizacija i predstavljaju glavne biotske zajednice koje podržavaju život na Zemlji.
3 Zoniranje makroekosustava
Proučavanje geografske distribucije ekosustava može se poduzeti samo na razini velikih ekoloških cjelina – makroekosustava, koji se promatraju na kontinentalnoj razini. Ekosustavi nisu raštrkani u neredu, naprotiv, grupirani su u prilično pravilne zone, vodoravno (po širini) i okomito (po visini). To potvrđuje periodični zakon geografske zonalnosti A.A. Grigorieva - M.I. Budyko: s promjenom fizičko-geografskih zona Zemlje povremeno se ponavljaju slične krajobrazne zone i neka njihova opća svojstva. O tome se također raspravljalo kada se razmatralo tlo-zračno okruženje života. Zakonom utvrđena periodičnost očituje se u činjenici da vrijednosti indeksa suhoće variraju u različitim zonama od 0 do 4-5, tri puta između polova i ekvatora blizu su jedinice. Ove vrijednosti odgovaraju najvećoj biološkoj produktivnosti krajolika.
Periodično ponavljanje svojstava u nizu sustava jedne hijerarhijske razine vjerojatno je opći zakon svemira, formuliran kao zakon periodičnosti u strukturi sustavnih agregata, ili sustavno-periodički zakon - specifični prirodni sustavi jedne razine ( podrazina) organizacije čine periodični ili ponavljajući niz morfološki sličnih struktura unutar gornjih i donjih prostorno-vremenskih granica sustava, izvan kojih postojanje sustava na danoj razini postaje nemoguće. Oni prelaze u nestabilno stanje ili se pretvaraju u drugu strukturu sustava, uključujući drugu razinu organizacije.
Temperatura i padaline određuju položaj glavnih kopnenih bioma na zemljinoj površini. Obrazac temperature i padalina u određenom području tijekom prilično dugog vremenskog razdoblja je ono što nazivamo klimom. Klima u različitim dijelovima svijeta je različita. Godišnja količina padalina varira od 0 do 2500 mm ili više. Režimi temperature i padalina kombiniraju se na vrlo različite načine.
Specifičnosti klimatskih uvjeta pak određuju razvoj pojedinog bioma.
Od ekvatora do polova vidljiva je određena simetrija u rasporedu bioma različitih hemisfera:
Tropske kišne šume (sjever Južne Amerike, Srednja Amerika, zapadni i središnji dijelovi ekvatorijalne Afrike, jugoistočna Azija, obalna područja sjeverozapadne Australije, otoci Indijskog i Tihog oceana). Klima je bez promjene godišnjih doba (blizina ekvatora), prosječna godišnja temperatura je iznad 17°C (obično 28°C), prosječna godišnja količina padalina prelazi 2400 mm.
Vegetacija: dominiraju šume. Neke vrste drveća visoke su i do 60 m. Na njihovim deblima i granama nalaze se epifitske biljke, čije korijenje ne dopire do tla, te drvenaste loze koje se ukorijenjuju u tlu i penju se na stabla do vrhova. Sve to čini gustu krošnju.
Fauna: sastav vrsta je bogatiji nego u svim ostalim biomima zajedno. Osobito su brojni vodozemci, gmazovi i ptice (žabe, gušteri, zmije, papige), majmuni i drugi mali sisavci, egzotični kukci jarkih boja, au akumulacijama ribe jarkih boja.
Značajke: tla su često tanka i siromašna, većina hranjivih tvari sadržana je u površinskoj biomasi ukorijenjene vegetacije.
2. Savane (subekvatorijalna Afrika, Južna Amerika, značajan dio južne Indije). Klima je suha i vruća veći dio godine. Obilne padaline tijekom kišne sezone. Temperatura: srednja godišnja-vis. Padalina iznosi 750-1650 mm/god, uglavnom tijekom kišne sezone. Vegetacija - biljke plave trave (žitarice) s rijetkim listopadnim drvećem. Fauna: veliki sisavci biljojedi, kao što su antilope, zebre, žirafe, nosoroze, predatori - lavovi, leopardi, gepardi.
Pustinje (neka područja Afrike, na primjer Sahara; Bliski istok i središnja Azija, Veliki bazen i jugozapad SAD-a i sjeverni Meksiko itd.). Klima je vrlo suha. Temperatura - topli dani i hladne noći. Padalina je manje od 250 mm/god. Vegetacija: rijetki grmovi, često trnoviti, ponekad kaktusi i niske trave, brzo prekrivaju tlo cvjetnim tepihom nakon rijetkih kiša. Biljke imaju opsežan površinski korijenski sustav koji hvata vlagu iz rijetkih oborina, kao i korijenje koje prodire u tlo do razine podzemne vode (30 m i dublje). Fauna: razni glodavci (klokan i dr.), krastače, gušteri, zmije i drugi gmazovi, sove, orlovi, lešinari, male ptice i kukci u velikim količinama.
4. Stepe (središte Sjeverne Amerike, Rusija, pojedina područja Afrike i Australije, jugoistok Južne Amerike). Klima je sezonska. Temperature - ljetne temperature kreću se od umjereno toplih do vrućih, zimske ispod 0°C. Padalina - 750-2000 mm/god. Vegetacija: dominira modra trava (žitarice) visine do 2 m i više u nekim prerijama Sjeverne Amerike ili do 50 cm, na primjer, u ruskim stepama, s izoliranim drvećem i grmljem u vlažnim područjima. Fauna: veliki sisavci biljojedi - bizon, antilopa viloroga (Sjeverna Amerika), divlji konji (Euroazija), klokani (Australija), žirafe, zebre, bijeli nosorozi, antilope (Afrika); Grabežljivci uključuju kojote, lavove, leoparde, geparde, hijene, razne ptice i male sisavce koji kopaju jame poput zečeva, vjeverica i mrtvaca.
5. Umjerene šume (zapadna Europa, istočna Azija, istočni SAD). Klima - sezonska sa zimskim temperaturama ispod 0 0C. Padalina - 750-2000 mm/god. Vegetacija: prevladavaju šume širokolisnog listopadnog drveća do 35-45 m visine (hrast, hikorija, javor), grmolika šikara, mahovine, lišajevi. Fauna: sisavci (bjelorepi jelen, dikobraz, rakun, oposum, vjeverica, zec, rovke), ptice (cvrčice, djetlići, kosovi, sove, sokolovi), zmije, žabe, daždevnjaci, ribe (pastrve, grgeči, somovi i dr. .), bogata mikrofauna tla (Sl. 12.3). Biota je prilagođena sezonskoj klimi: zimski san, seoba, mirovanje u zimskim mjesecima. 6. Crnogorične šume, tajga (sjeverna područja Sjeverne Amerike, Europe i Azije). Klima je duga i hladna zima, s nešto padalina koje padaju u obliku snijega. Vegetacija: prevladavaju vazdazelene crnogorične šume, pretežno smreke, bora i jele. Životinjski svijet: veliki biljojedi papkari (mazga, sob), mali biljojedi sisavci (zec, vjeverica, glodavci), vuk, ris, lisica, crni medvjed, grizli, vukodlak, kunac i drugi grabežljivci, brojni krvopije u kratkom ljetu vrijeme. Klima je vrlo hladna s polarnim danima i polarnim noćima. Prosječna godišnja temperatura je ispod - 5°C. U nekoliko tjedana kratkog ljeta tlo se otopi ne više od jednog metra duboko. Padalina je manje od 250 mm/god. Vegetacija: dominiraju spororastući lišajevi, mahovine, trave i šaševi te patuljasto grmlje. Fauna: veliki biljojedi papkari (sobovi, mošusni volovi), mali sisavci koji se ukopaju (tijekom cijele godine, na primjer, leminzi), grabežljivci koji zimi poprimaju maskirnu bijelu boju (arktička lisica, ris, hermelin, snježna sova). U kratkom ljetu u tundri se gnijezdi veliki broj ptica selica, među kojima je posebno mnogo ptica močvarica, koje se hrane kukcima i slatkovodnim beskralješnjacima kojih ovdje ima u izobilju. Vrlo je jasna i vertikalna visinska zonalnost kopnenih ekosustava, posebno na mjestima s izraženim reljefom. Vlažnost je glavni faktor koji određuje vrstu bioma. Uz dovoljno velike količine padalina razvija se šumska vegetacija. Temperatura određuje vrstu šume. Potpuno je ista situacija u stepskim i pustinjskim biomima. Promjene u tipovima vegetacije u hladnim krajevima nastaju s nižim godišnjim količinama oborine, budući da se pri niskim temperaturama manje vode gubi isparavanjem. Temperatura postaje glavni faktor samo u vrlo hladnim uvjetima s permafrostom. Dakle, sastav ekosustava uvelike ovisi o njihovoj funkcionalnoj "namjeni" i obrnuto. Prema N.F. Reimers (1994), to se ogleda u principu ekološke komplementarnosti (komplementarnosti): niti jedan funkcionalni dio ekosustava (ekološka komponenta, element itd.) ne može postojati bez drugih funkcionalno komplementarnih dijelova. Njemu je blisko i proširuje ga načelo ekološke podudarnosti (korespondencije):. funkcionalno se nadopunjujući, žive komponente ekosustava razvijaju za to odgovarajuće prilagodbe, usklađene s uvjetima abiotičkog okoliša, koji je u velikoj mjeri transformiran istim organizmima (bioklima i dr.), tj. postoji dvostruki niz korespondencije – između organizama i njihovog staništa – vanjskog i stvorenog cenozom. 4 Struktura ekosustava S gledišta trofičke strukture (od grčkog trophe - prehrana), ekosustav se može podijeliti u dvije razine: Gornji - autotrofni (samohraneći) sloj ili "zeleni pojas", uključujući biljke ili njihove dijelove koji sadrže klorofil, gdje prevladava fiksacija energije brojanja i korištenje jednostavnih anorganskih spojeva. Donji je heterotrofni (koji se hrani od drugih) sloj, ili "smeđi pojas" tla i sedimenata, raspadajuće tvari, korijenja itd., u kojem prevladava korištenje, transformacija i razgradnja složenih spojeva. S biološke točke gledišta, sljedeće komponente razlikuju se kao dio ekosustava: U ekosustavu su prehrambene i energetske veze među kategorijama uvijek nedvosmislene i idu u smjeru: autotrofi - heterotrofi ili u potpunijem obliku; autotrofi -> potrošači -> razlagači (destruktori). Primarni izvor energije za ekosustave je Sunce. Protok energije (prema T.A. Akimova, V.V. Khaskin, 1994.) koji sunce šalje planeti Zemlji prelazi 20 milijuna EJ godišnje. Samo četvrtina ovog toka se približava granici atmosfere. Od toga se oko 70% reflektira, apsorbira atmosfera i emitira u obliku dugovalnog infracrvenog zračenja. Sunčevo zračenje koje pada na Zemljinu površinu iznosi 1,54 milijuna EJ godišnje. Ova ogromna količina energije je 5000 puta veća od ukupne energije čovječanstva na kraju 20. stoljeća i 5,5 puta veća od energije svih dostupnih izvora fosilnih goriva organskog podrijetla, akumuliranih tijekom najmanje 100 milijuna godina. Većina sunčeve energije koja dospije na površinu planeta pretvara se izravno u toplinu, zagrijavajući vodu ili tlo, što zauzvrat zagrijava zrak. Ta toplina služi kao pokretačka snaga ciklusa vode, zračnih struja i oceanskih struja koje određuju vrijeme te se postupno oslobađa u svemir, gdje se gubi. Da bismo odredili mjesto ekosustava u ovom prirodnom tijeku energije, važno je shvatiti da, bez obzira na to koliko su opsežni i složeni, oni koriste samo mali dio nje. To implicira jedno od temeljnih načela funkcioniranja ekosustava: oni postoje zahvaljujući nezagađujućoj i gotovo vječnoj sunčevoj energiji, čija je količina relativno stalna i izdašna. Navedimo detaljnije svaku od navedenih karakteristika sunčeve energije: 2. Čimbenici koji osiguravaju cjelovitost ekosustava 1 Ciklusi tvari Oba ciklusa su međusobno povezana i predstavljaju takoreći jedan proces. Procjenjuje se da sav kisik sadržan u atmosferi kruži kroz organizme (spaja se tijekom disanja i oslobađa tijekom fotosinteze) u 2000 godina, ugljični dioksid u atmosferi kruži u suprotnom smjeru u 300 godina, a sva voda na Zemlji je razgradio i ponovno stvorio fotosintezom i disanjem u 2 000 000 godina. Međudjelovanje abiotskih čimbenika i živih organizama ekosustava prati kontinuirano kruženje tvari između biotopa i biocenoze u obliku izmjeničnih organskih i mineralnih spojeva. Razmjena kemijskih elemenata između živih organizama i anorganskog okoliša, čiji se različiti stupnjevi odvijaju unutar ekosustava, naziva se biogeokemijska cirkulacija ili biogeokemijski ciklus. Kruženje vode. Najznačajniji ciklus na Zemlji po prenesenim masama i potrošnji energije je planetarni hidrološki ciklus - ciklus vode. Svake sekunde u njega je uključeno 16,5 milijuna m3 vode i na to se troši više od 40 milijardi MW sunčeve energije, (prema T.A. Akimova V.V. Haskin, (1994.)). Ali ovaj ciklus nije samo prijenos vodenih masa. To su fazne transformacije, stvaranje otopina i suspenzija, taloženje, kristalizacija, procesi fotosinteze, kao i razne kemijske reakcije. Život je nastao i traje u ovoj sredini. Voda je osnovni element neophodan za život. Kvantitativno je najčešća anorganska komponenta žive tvari. Kod ljudi voda čini 63% tjelesne težine, kod gljiva - 80%, kod biljaka - 80-90%, a kod nekih meduza - 98% Voda, kao što ćemo vidjeti malo kasnije, sudjeluje u biološkom ciklusu i služi kao izvor vodika i kisika, čineći samo mali dio njezinog ukupnog volumena. U tekućem, čvrstom i parovitom stanju voda je prisutna u sve tri glavne komponente biosfere: atmosferi, hidrosferi i litosferi. Sve vode su ujedinjene zajedničkim konceptom "hidrosfere". Komponente hidrosfere međusobno su povezane stalnom razmjenom i međudjelovanjem. Voda, neprestano prelazeći iz jednog stanja u drugo, čini male i velike cikluse. Isparavanje vode s površine oceana, kondenzacija vodene pare u atmosferi i oborina na površini oceana čine mali ciklus. Kada vodenu paru zračne struje nose na kopno, ciklus postaje mnogo složeniji. U tom slučaju, dio oborina ispari i vrati se u atmosferu, drugi hrani rijeke i rezervoare, ali se na kraju vraća u ocean kroz riječno i podzemno otjecanje, čime se završava veliki ciklus. Biotički (biološki) ciklus. Biotički (biološki) ciklus odnosi se na kruženje tvari između tla, biljaka, životinja i mikroorganizama. Biotički (biološki) ciklus je protok kemijskih elemenata iz tla, vode i atmosfere u žive organizme, transformacija pristiglih elemenata u nove složene spojeve i njihov povratak natrag u proces života uz godišnji pad dijela organskih tvari. tvari ili s potpuno mrtvim organizmima uključenim u sastav ekosustava. Sada ćemo prikazati biotički ciklus u cikličkom obliku. Središnji biotički ciklus (prema T.A. Akimova, V.V., Khaskhin) sastojao se od primitivnih jednostaničnih proizvođača (P) i dekompozitora-destruktora (D). Mikroorganizmi se mogu brzo razmnožavati i prilagođavati različitim uvjetima, na primjer, koristiti sve vrste supstrata - izvora ugljika - u svojoj prehrani. Viši organizmi nemaju takve sposobnosti. U cjelovitim ekosustavima mogu postojati u obliku strukture na temelju mikroorganizama. Prvo se razvijaju višestanične biljke (P) – viši proizvođači. Zajedno s jednostaničnim organizmima stvaraju organsku tvar procesom fotosinteze, koristeći energiju sunčevog zračenja. Naknadno se spajaju primarni konzumenti - biljojedi (T), a potom konzumenti mesojedi. Ispitali smo biotički ciklus kopna. To se u potpunosti odnosi na biotički ciklus vodenih ekosustava, na primjer, oceana. Svi organizmi zauzimaju određeno mjesto u biotičkom ciklusu i obavljaju svoje funkcije transformacije ogranaka protoka energije koje primaju i prijenosa biomase. Sustav jednostaničnih razlagača (destruktora) ujedinjuje sve, depersonalizira njihove tvari i zatvara opći krug. Oni u abiotski okoliš biosfere vraćaju sve elemente potrebne za nove i nove revolucije. Treba istaknuti najvažnije značajke biotskog ciklusa. Fotosinteza je snažan prirodni proces koji svake godine uključuje goleme mase biosferske tvari u ciklus i određuje njezin visoki kisikov potencijal. Zbog ugljičnog dioksida i vode dolazi do sintetizacije organske tvari i oslobađanja slobodnog kisika. Izravni produkti fotosinteze su različiti organski spojevi, a općenito je proces fotosinteze prilično složen. Osim fotosinteze uz sudjelovanje kisika, tzv. kisikove fotosinteze, treba se usmjeriti i na fotosintezu bez kisika, odnosno kemosintezu. Kemosintetski organizmi uključuju nitrifikatore, karboksidobakterije, sumporne bakterije, tionske željezne bakterije i vodikove bakterije. Ime su dobili po svojim oksidacijskim supstratima, koji mogu biti NH3, NO2, CO, H2S, S, Fe2+, H2. Neke vrste su obvezni kemolitoautotrofi, druge su fakultativne. Potonji uključuju karboksidobakterije i vodikove bakterije. Kemosinteza je karakteristična za hidrotermalne izvore dubokih mora. Fotosinteza se odvija, osim nekoliko izuzetaka, na cijeloj površini Zemlje, stvara ogroman geokemijski učinak i može se izraziti količinom cjelokupne mase ugljika koji godišnje učestvuje u izgradnji organske – žive tvari cijele biosfere. Opći ciklus tvari povezan s izgradnjom organske tvari fotosintezom uključuje i kemijske elemente poput N, P, S, kao i metale - K, Ca, Mg, Na, Al. Kada organizam ugine, dolazi do obrnutog procesa - razgradnje organske tvari oksidacijom, raspadanjem itd. uz stvaranje konačnih proizvoda razgradnje. U biosferi Zemlje ovaj proces dovodi do činjenice da količina biomase žive tvari ima tendenciju da bude donekle konstantna. Biomasa ekosfere (2 10|2t) je sedam redova veličine manja od mase zemljine kore (2 .10|9t). Biljke Zemlje godišnje proizvedu organsku tvar jednaku 1,6-10"% ili 8% biomase ekosfere. Destruktori, koji čine manje od 1% ukupne biomase organizama planete, prerađuju masu organske tvari koja iznosi 10 puta veća od vlastite biomase.U prosjeku je razdoblje obnavljanja biomase 12,5 godina.Pretpostavimo da su masa žive tvari i produktivnost biosfere bile iste od kambrija do danas (530 milijuna godina), tada ukupna količina organske tvari koja je prošla kroz globalni biotički ciklus i koju je iskoristio život na planeti bit će 2,10 " 2-5,ZL08/12,5=8,5L0|9t, što je 4 puta više od mase zemljine kore. Što se tiče ovih izračuna, N.S. Pečurkin (1988.) je napisao: “Možemo reći da su atomi koji čine naša tijela bili u drevnim bakterijama, i u dinosaurima, i u mamutima.” Zakon biogene migracije atoma V.I. Vernadsky navodi: „Migracija kemijskih elemenata na zemljinoj površini i u biosferi kao cjelini događa se ili uz izravno sudjelovanje žive tvari (biogena migracija), ili se događa u okolišu čije su geokemijske značajke (O2, CO2, H2, itd.) su određena živa tvar, kako ona koja trenutno nastanjuje biosferu, tako i ona koja je djelovala na Zemlju kroz geološku povijest." U I. Vernadskog 1928.-1930 u svojim dubokim generalizacijama o procesima u biosferi dao je ideju o pet glavnih biogeokemijskih funkcija žive tvari. Prva funkcija je plin. Druga funkcija je koncentracija. Treća funkcija je redoks. Četvrta funkcija je biokemijska. Peta funkcija je biogeokemijska aktivnost čovječanstva, koja obuhvaća sve veću količinu tvari u zemljinoj kori za potrebe industrije, prometa i poljoprivrede. Biološki ciklus varira u različitim prirodnim zonama i klasificira se prema nizu pokazatelja: biljna biomasa, stelja, stelja, količina elemenata fiksiranih u biomasi itd. Ukupna biomasa najveća je u šumskom pojasu, a najmanji je udio podzemnih organa u šumama. To potvrđuje i indeks intenziteta biološkog ciklusa – omjer mase legla i dijela legla koji ga čini. Ciklus ugljika. Od svih biogeokemijskih ciklusa, ciklus ugljika je bez sumnje najintenzivniji. Ugljik cirkulira velikim brzinama između različitih anorganskih sredstava i kroz hranidbene mreže unutar zajednica živih organizama. CO i CO2 igraju određenu ulogu u ciklusu ugljika.Često je u Zemljinoj biosferi ugljik predstavljen najpokretljivijim oblikom CO2. Izvor primarnog ugljičnog dioksida u biosferi je vulkanska aktivnost povezana sa sekularnim otplinjavanjem plašta i nižih horizonata zemljine kore. Migracija CO2 u biosferi odvija se na dva načina. Prvi način je apsorbiranje tijekom fotosinteze uz stvaranje glukoze i drugih organskih tvari od kojih su izgrađena sva biljna tkiva. Nakon toga se transportiraju kroz prehrambene lance i tvore tkiva svih drugih živih bića u ekosustavu. Odumiranjem biljaka i životinja na površini dolazi do oksidacije organskih tvari uz stvaranje CO2. Atomi ugljika se također vraćaju u atmosferu kada organska tvar izgara. Važna i zanimljiva značajka kruženja ugljika je da u dalekim geološkim epohama, prije više stotina milijuna godina, značajan dio organske tvari nastale u procesima fotosinteze nije iskorišten ni od konzumenata ni od razlagača, već se akumulirao u litosferi. u obliku fosilnih goriva: nafte, ugljena, uljnog škriljevca, treseta, itd. Ta se fosilna goriva iskopavaju u ogromnim količinama kako bi se zadovoljile energetske potrebe našeg industrijskog društva. Njegovim spaljivanjem mi, na neki način, dovršavamo ciklus ugljika. Drugim putem, migracija ugljika se odvija stvaranjem karbonatnog sustava u različitim ležištima, gdje CO2 prelazi u H2CO3, HCO, CO3. Uz pomoć kalcija (ili magnezija) otopljenog u vodi, karbonati (CaCO3) se talože biogenim i abiogenim putem. Nastaju debeli slojevi vapnenca. Prema riječima A.B. Ronov, omjer zakopanog ugljika u produktima fotosinteze prema ugljiku u karbonatnim stijenama je 1:4. Uz veliki ciklus kruženja ugljika, postoji i niz malih ciklusa ugljika na kopnu iu oceanu. Općenito, bez antropogenih intervencija, sadržaj ugljika u biogeokemijskim rezervoarima: biosferi (biomasa + tlo i detritus), sedimentnim stijenama, atmosferi i hidrosferi, održava se s visokim stupnjem postojanosti (prema T.A. Akimova, V.V. Haskin (1994.) ). Stalna izmjena ugljika, s jedne strane, između biosfere, as druge, između atmosfere i hidrosfere, određena je plinskom funkcijom žive tvari - procesima fotosinteze, disanja i destrukcije, a iznosi oko 6-1010 tona/god. Postoji protok ugljika u atmosferu i hidrosferu i tijekom vulkanske aktivnosti prosječno 4,5 106 t/god. Ukupna masa ugljika u fosilnim gorivima (nafta, plin, ugljen itd.) procjenjuje se na 3,2*1015 tona, što odgovara prosječnoj stopi akumulacije od 7 milijuna tona/god. Ta je količina beznačajna u usporedbi s masom cirkulirajućeg ugljika i, takoreći, ispala je iz ciklusa i izgubljena u njemu. Dakle, stupanj otvorenosti (nesavršenosti) ciklusa je 10"4, odnosno 0,01%, a prema tome, stupanj zatvorenosti je 99,99%. To znači, s jedne strane, da je svaki atom ugljika sudjelovao u ciklusu desetke tisuća puta prije ispadanja iz ciklusa završili u dubinama.S druge strane, tokovi sinteze i raspadanja organskih tvari u biosferi vrlo su precizno usklađeni jedni s drugima. 0,2% mobilnih zaliha ugljika je u stalnoj cirkulaciji. Ugljik iz biomase obnavlja se za 12 godina, u atmosferi - za 8 godina. Ciklus kisika. Kisik (O2) ima važnu ulogu u životu većine živih organizama na našem planetu. U kvantitativnom smislu, ovo je glavna komponenta žive tvari. Na primjer, ako uzmemo u obzir vodu sadržanu u tkivima, ljudsko tijelo sadrži 62,8% kisika i 19,4% ugljika. Općenito, u biosferi ovaj element, u usporedbi s ugljikom i vodikom, glavni je među jednostavnim tvarima. Unutar biosfere dolazi do brze izmjene kisika sa živim organizmima ili njihovim ostacima nakon smrti. Biljke, u pravilu, proizvode slobodni kisik, a životinje ga troše disanjem. Kao najrasprostranjeniji i najpokretljiviji element na Zemlji, kisik ne ograničava postojanje i funkcije ekosfere, iako dostupnost kisika za vodene organizme može biti privremeno ograničena. Ciklus kisika u biosferi izuzetno je složen jer s njim reagira velik broj organskih i anorganskih tvari. Kao rezultat toga, pojavljuju se mnogi epicikli, koji se javljaju između litosfere i atmosfere ili između hidrosfere i ova dva okoliša. Ciklus kisika je na neki način sličan obrnutom ciklusu ugljičnog dioksida. Kretanje jednog događa se u smjeru suprotnom od kretanja drugog. Potrošnja atmosferskog kisika i njegova zamjena primarnim proizvođačima odvija se relativno brzo. Dakle, potrebno je 2000 godina da se potpuno obnovi sav atmosferski kisik. Danas ne dolazi do nakupljanja kisika u atmosferi, a njegov sadržaj (20,946%) ostaje konstantan. U gornjim slojevima atmosfere, kada ultraljubičasto zračenje djeluje na kisik, nastaje ozon - O3. Oko 5% sunčeve energije koja dospije do Zemlje troši se na stvaranje ozona - oko 8,6 * 1015 W. Reakcije su lako reverzibilne. Kada se ozon raspada, oslobađa se ta energija koja održava visoke temperature u gornjoj atmosferi. Prosječna koncentracija ozona u atmosferi je oko 106 vol. %; maksimalna koncentracija O3 - do 4-10 "* vol.% - postiže se na visinama od 20-25 km (T.A. Akimova, V.V. Haskin, 1998.). Ozon služi kao neka vrsta UV filtera: blokira značajan dio jakih ultraljubičastih zraka. Vjerojatno je stvaranje ozonskog omotača bio jedan od uvjeta za izlazak života iz oceana i naseljavanje kopna. Većina kisika proizvedenog tijekom geoloških epoha nije ostala u atmosferi, već ga je fiksirala litosfera u obliku karbonata, sulfata, željeznih oksida itd. Ova masa je 590 * 1014 tona u odnosu na 39 * 1014 tona kisika, koji cirkulira u biosferi u obliku plina ili sulfata otopljenih u kontinentalnim i oceanskim vodama. Ciklus dušika. Dušik je esencijalni biogeni element, jer je dio proteina i nukleinskih kiselina. Ciklus dušika je jedan od najsloženijih, jer uključuje i plinsku i mineralnu fazu, a ujedno i najidealnije cikluse. Ciklus dušika usko je povezan s ciklusom ugljika. U pravilu, dušik slijedi nakon ugljika, zajedno s kojim sudjeluje u stvaranju svih proteinskih tvari. Atmosferski zrak, koji sadrži 78% dušika, neiscrpan je rezervoar. Međutim, većina živih organizama ne može izravno koristiti ovaj dušik. Da bi biljke apsorbirale dušik, on mora biti dio amonijevih (NH*) ili nitratnih (NO3) iona. Plinoviti dušik kontinuirano se ispušta u atmosferu kao rezultat rada denitrifikacijskih bakterija, a fiksacijske bakterije ga zajedno s modrozelenim algama (cijanofitima) neprestano apsorbiraju pretvarajući ga u nitrate. Ciklus dušika jasno je vidljiv na razini destruktora. Proteini i drugi oblici organskog dušika sadržani u biljkama i životinjama nakon njihove smrti izloženi su heterotrofnim bakterijama, aktinomicetama, gljivama (bioredukcijskim mikroorganizmima), koji proizvode potrebnu energiju redukcijom ovog organskog dušika pretvarajući ga u amonijak. U tlu se odvija proces nitrifikacije koji se sastoji od niza reakcija, gdje uz sudjelovanje mikroorganizama dolazi do oksidacije amonijevog iona (NH4+) u nitrit (NO~) ili nitrita u nitrat (N0~). Redukcija nitrita i nitrata u plinovite spojeve molekularni dušik (N2) ili dušikov oksid (N20) bit je procesa denitrifikacije. Stvaranje nitrata anorganskim putem u malim količinama stalno se događa u atmosferi vezanjem atmosferskog dušika s kisikom tijekom električnih pražnjenja tijekom grmljavinske oluje, a zatim pada s kišom na površinu tla. Drugi izvor atmosferskog dušika su vulkani, koji nadoknađuju gubitak dušika isključenog iz ciklusa tijekom sedimentacije ili taloženja na dnu oceana. Općenito, prosječna opskrba nitratnog dušika abiotskog podrijetla tijekom taloženja iz atmosfere u tlo ne prelazi 10 kg (godina/ha), slobodne bakterije proizvode 25 kg (godina/ha), dok simbioza Rhizobiuma s leguminoznim biljkama proizvodi prosječno 200 kg (godina/ha). Pretežni dio vezanog dušika denitrifikacijske bakterije prerađuju u N2 i vraćaju u atmosferu. Samo oko 10% amonificiranog i nitrificiranog dušika više biljke apsorbiraju iz tla i završava na raspolaganju višestaničnim predstavnicima biocenoza Ciklus fosfora. Ciklus fosfora u biosferi povezan je s metaboličkim procesima u biljkama i životinjama. Ovaj važan i neophodan element protoplazme, sadržan u kopnenim biljkama i algama 0,01-0,1%, životinje od 0,1% do nekoliko posto, cirkulira, postupno se pretvarajući iz organskih spojeva u fosfate, koje biljke opet mogu koristiti. Međutim, fosfor, za razliku od drugih biofilnih elemenata, tijekom migracije ne stvara plinoviti oblik. Rezervoar fosfora nije atmosfera, poput dušika, već mineralni dio litosfere. Glavni izvori anorganskog fosfora su magmatske stijene (apatiti) ili sedimentne stijene (fosforiti). Iz stijena se anorganski fosfor uključuje u cirkulaciju ispiranjem i otapanjem u kontinentalnim vodama. Ulazeći u kopnene ekosustave i tlo, biljke apsorbiraju fosfor iz vodene otopine u obliku anorganskog fosfatnog iona i uključuju se u različite organske spojeve, gdje se pojavljuje u obliku organskog fosfata. Fosfor se kreće hranidbenim lancima od biljaka do drugih organizama u ekosustavu. Fosfor se prenosi u vodene ekosustave tekućim vodama. Rijeke neprestano obogaćuju oceane fosfatima. Gdje fosfor postaje dio fitoplanktona. Neki spojevi fosfora migriraju unutar malih dubina, konzumirajući ih organizmi, dok se drugi dio gubi na velikim dubinama. Mrtvi ostaci organizama dovode do nakupljanja fosfora na različitim dubinama. Ciklus sumpora. Postoje brojni plinoviti spojevi sumpora, kao što su sumporovodik H2S i sumporov dioksid SO2. Međutim, pretežni dio ciklusa ovog elementa je sedimentne prirode i javlja se u tlu i vodi.Dostupnost anorganskog sumpora u ekosustavu je olakšana dobrom topljivošću mnogih sulfata u vodi. Biljke, apsorbirajući sulfate, reduciraju ih i proizvode aminokiseline koje sadrže sumpor (metionin, cistein, cistin) koje igraju važnu ulogu u razvoju tercijarne strukture proteina tijekom stvaranja disulfidnih mostova između različitih zona polipeptidnog lanca. Mnoge osnovne značajke biogeokemijskog ciklusa jasno su vidljive: S -> S -> SO2 - bezbojne, zelene i ljubičaste sumporne bakterije; - "H2S (anaerobna redukcija sulfata) - Desulfovibno; H2S - "SO2" (aerobna oksidacija sulfida) - tiobacil; organski S u SO i H2S. - aerobnih i anaerobnih heterotrofnih mikroorganizama. Primarna proizvodnja osigurava ugradnju sulfata u organsku tvar, a izlučivanje životinja služi kao način vraćanja sulfata u ciklus. Općenito, ekosustav zahtijeva manje sumpora u usporedbi s dušikom i fosforom. Stoga je sumpor rjeđe ograničavajući faktor za biljke i životinje. U isto vrijeme, ciklus sumpora je ključan u cjelokupnom procesu proizvodnje i razgradnje biomase. Na primjer, kada se željezni sulfidi stvaraju u sedimentima, fosfor prelazi iz netopljivog oblika u topljivi oblik i postaje dostupan organizmima. Ovo je potvrda kako je jedan ciklus reguliran drugim. 2 Protok energije u ekosustavima Za razliku od tvari koje kontinuirano kruže kroz različite blokove ekosustava, a koje se uvijek mogu ponovno upotrijebiti i ući u ciklus, energija se može iskoristiti jednom, tj. postoji linearni tok energije kroz ekosustav. Jednosmjerni dotok energije kao univerzalni prirodni fenomen nastaje kao posljedica zakona termodinamike. Prvi zakon kaže da se energija može pretvoriti iz jednog oblika (kao što je svjetlost) u drugi (kao što je potencijalna energija hrane), ali se ne može stvoriti ili uništiti. Drugi zakon kaže da ne može postojati niti jedan proces povezan s transformacijom energije bez gubitka dijela energije. Određena količina energije u takvim transformacijama se rasipa u nedostupnu toplinsku energiju i time se gubi. Dakle: ne može doći do transformacije, na primjer, prehrambenih tvari u tvar koja čini tijelo organizma, a koja bi se dogodila sa 100% učinkovitosti. Dakle, živi organizmi su pretvarači energije. I svaki put kada se energija pretvara, dio se gubi u obliku topline. U konačnici, sva energija koja ulazi u biotski ciklus ekosustava rasipa se kao toplina. Živi organizmi zapravo ne koriste toplinu kao izvor energije za rad - oni koriste svjetlost i kemijsku energiju. Hranidbeni lanci i mreže, trofičke razine. Unutar ekosustava, tvari koje sadrže energiju stvaraju autotrofni organizmi i služe kao hrana za heterotrofe. Prehrambene veze su mehanizmi za prijenos energije iz jednog organizma u drugi. Tipičan primjer: životinja jede biljke. Ovu životinju, zauzvrat, može pojesti druga životinja. Na taj se način energija može prenositi kroz niz organizama - svaki sljedeći se hrani prethodnim koji ga opskrbljuje sirovinama i energijom. Ovaj slijed prijenosa energije naziva se hranidbeni (trofički) lanac ili hranidbeni lanac. Položaj svake karike u hranidbenom lancu je trofička razina. Prvu trofičku razinu, kao što je ranije navedeno, zauzimaju autotrofi ili takozvani primarni proizvođači. Organizmi druge trofičke razine nazivaju se primarni potrošači, treći - sekundarni potrošači itd. Općenito postoje tri vrste hranidbenih lanaca. Lanac ishrane mesoždera počinje s biljkama i kreće se od malih organizama prema sve većim organizmima. Na kopnu se prehrambeni lanci sastoje od tri do četiri karike. Jedan od najjednostavnijih prehrambenih lanaca izgleda ovako: biljka -> zec -> vuk proizvođač -" biljojed -> -> mesožder Sljedeći prehrambeni lanci također su široko rasprostranjeni: Biljni materijal (npr. nektar) - "muha -" -" pauk -> rovka -> sova. Sok ružinog grma -> lisna uš -> -> bubamara -> -> pauk - "ptica insektivorka -> ptica grabljivica. U vodenim, a posebno u morskim ekosustavima, lanci ishrane predatora obično su duži nego u kopnenim. Treća vrsta hranidbenih lanaca, počevši od mrtvih biljnih ostataka, lešina i životinjskih izlučevina, nazivaju se detritalnim (saprofitnim) hranidbenim lancima ili detritalnim lancima razgradnje. Listopadne šume igraju važnu ulogu u hranidbenim lancima kopnenih ekosustava, čiju većinu lišća ne jedu biljojedi i dio je otpalog lišća. Listove usitnjavaju brojni detritivori - gljivice, bakterije, kukci (primjerice, kolembola) itd., a zatim ih progutaju gliste koje ravnomjerno raspoređuju humus u površinskom sloju zemlje tvoreći tzv. Na ovoj razini gljive razvijaju micelij. Mikroorganizmi raspadanja koji dovršavaju lanac proizvode konačnu mineralizaciju mrtve organske tvari. Općenito, tipični hranidbeni lanci detrita naših šuma mogu se predstaviti na sljedeći način: Listnjak -> glista -> kos - kobac; Mrtva životinja - "ličinke strvine muhe -" žaba -> smuk. U dijagramima hranidbenog lanca o kojima se govori, svaki organizam je predstavljen kako se hrani drugim organizmima iste vrste. Stvarne prehrambene veze u ekosustavu mnogo su složenije, budući da se životinja može hraniti organizmima različitih vrsta iz istog hranidbenog lanca ili iz različitih prehrambenih lanaca, na primjer, grabežljivcima viših trofičkih razina. Životinje se često hrane i biljkama i drugim životinjama. Zovu se svejedi. Hranidbene mreže u ekosustavima vrlo su složene i možemo zaključiti da je energiji koja u njih ulazi potrebno dosta vremena da migrira iz jednog organizma u drugi. Ekološke piramide. Unutar svakog ekosustava, hranidbene mreže imaju dobro definiranu strukturu koju karakteriziraju priroda i broj organizama zastupljenih na svakoj razini različitih prehrambenih lanaca. Kako bi proučavali odnose između organizama u ekosustavu i kako bi ih grafički prikazali, obično koriste ekološke piramide, a ne dijagrame mreže hrane. Ekološke piramide izražavaju trofičku strukturu ekosustava u geometrijskom obliku. Izrađuju se u obliku pravokutnika iste širine, ali duljina pravokutnika mora biti proporcionalna vrijednosti objekta koji se mjeri. Odavde možete dobiti piramide brojeva, biomase i energije. Ekološke piramide odražavaju temeljne značajke bilo koje biocenoze kada prikazuju njezinu trofičku strukturu. Piramide brojeva. Oni predstavljaju najjednostavniju aproksimaciju proučavanja trofičke strukture ekosustava. Piramida biomase. Potpunije odražava prehrambene odnose u ekosustavu, budući da uzima u obzir ukupnu masu organizama (biomasu) svake trofičke razine. Piramida energije. Najosnovniji način prikazivanja veza između organizama na različitim trofičkim razinama je kroz energetske piramide. Oni predstavljaju učinkovitost pretvorbe energije i produktivnost hranidbenih lanaca i konstruirani su brojanjem količine energije (kcal) akumulirane po jedinici površine po jedinici vremena i koju koriste organizmi na svakoj trofičkoj razini. Sunčeva energija koju biljka primi samo se djelomično koristi u procesu fotosinteze. Energija fiksirana u ugljikohidratima predstavlja bruto proizvodnju ekosustava. Ugljikohidrati se koriste za izgradnju protoplazme i rast biljaka. Dio njihove energije troše na disanje. Konzumenti drugog reda (predatori) ne uništavaju cjelokupnu biomasu svojih žrtava. Štoviše, od količine koju uništavaju, samo se dio koristi za stvaranje biomase vlastite trofičke razine. Ostatak se uglavnom troši na energiju disanja i izlučuje se izmetom i izmetom. Godine 1942. R. Lindeman prvi je formulirao zakon piramide energija, koji se u udžbenicima često naziva "zakon od 10%". Prema tom zakonu, u prosjeku, ne više od 10% energije prelazi s jedne trofičke razine ekološke piramide na drugu razinu. Samo 10-20% početne energije prenosi se na sljedeće heterotrofe. Koristeći zakon energetske piramide, lako je izračunati da je količina energije koja dopire do tercijarnih mesoždera (trofička razina V) oko 0,0001 energije koju apsorbiraju proizvođači. Iz toga slijedi da se prijenos energije s jedne razine na drugu događa s vrlo niskom učinkovitošću. To objašnjava ograničeni broj karika u hranidbenom lancu, bez obzira na određenu biocenozu. E. Odum (1959.) u krajnje pojednostavljenom hranidbenom lancu - lucerna -> tele - "dijete je procijenilo transformaciju energije, ilustrirao veličinu njezinih gubitaka. Recimo, razmišljao je, postoje usjevi lucerne na površini od 4 hektara.Na ovom polju se hrane telad (pretpostavlja se da jedu samo lucernu), a dječak od 12 godina hrani se isključivo teletinom.Rezultati izračuna, prikazani u obliku tri piramide: brojeva, biomase i energije, pokazuju da lucerna koristi samo 0,24% ukupne sunčeve energije koja pada na polje; tele apsorbira 8% ovog proizvoda, a samo 0,7% biomase teleta osigurava razvoj djeteta tijekom godine. E. Odum je tako pokazao da se samo milijunti dio upadne sunčeve energije pretvara u biomasu mesoždera, u ovom slučaju pridonoseći povećanju tjelesne težine djeteta, a ostatak se gubi i rasipa u degradiranom obliku u okoliš. Gornji primjer jasno ilustrira vrlo nisku ekološku učinkovitost ekosustava i nisku učinkovitost transformacije u hranidbenim lancima. Možemo reći sljedeće: ako 1000 kcal (dan m 2) bilježe proizvođači, zatim 10 kcal (dan m 2) ulazi u biomasu biljojeda i samo 1 kcal (dan m 2) - u biomasu mesojeda. Budući da određenu količinu tvari svaka biocenoza može iskoristiti više puta, a dio energije jednom, svrsishodnije je reći da se u ekosustavu odvija kaskadni prijenos energije. Potrošači služe kao upravljačka i stabilizirajuća karika u ekosustavu. Potrošači stvaraju spektar raznolikosti u cenozi, sprječavajući monopol dominantnih. Pravilo kontrolne vrijednosti potrošača s pravom se može smatrati temeljnim. Prema kibernetičkim stajalištima, sustav upravljanja trebao bi biti složeniji po strukturi od upravljanog, tada postaje jasan razlog za mnoštvo vrsta imovine. Kontrolni značaj potrošača ima i energetsku osnovu. Protok energije kroz jednu ili drugu trofičku razinu ne može se apsolutno odrediti dostupnošću hrane u temeljnoj trofičkoj razini. Kao što je poznato, uvijek ostaje dovoljna “rezerva”, jer bi potpuno uništenje hrane dovelo do smrti potrošača. Ovi opći obrasci promatraju se u okviru populacijskih procesa, zajednica, razina ekološke piramide i biocenoza u cjelini. 3 Dinamički procesi koji osiguravaju cjelovitost i održivost ekosustava Cikličke promjene u zajednicama odražavaju dnevnu, sezonsku i dugoročnu periodičnost vanjskih uvjeta i manifestaciju endogenih ritmova organizama. Dnevna dinamika ekosustava uglavnom je povezana s ritmom prirodnih pojava i strogo je periodične prirode. Već smo smatrali da u svakoj biocenozi postoje skupine organizama čija se životna aktivnost odvija u različito doba dana. Neki su aktivni danju, drugi noću. Stoga dolazi do povremenih promjena u sastavu i omjeru pojedinih vrsta biocenoza pojedinog ekosustava, jer se pojedini organizmi na određeno vrijeme isključuju iz njega. Dnevnu dinamiku biocenoze osiguravaju i životinje i biljke. Kao što je poznato, intenzitet i priroda fizioloških procesa u biljkama mijenja se tijekom dana - fotosinteza se ne događa noću, često se cvjetovi u biljkama otvaraju samo noću i oprašuju ih noćne životinje, drugi su prilagođeni oprašivanju danju. Dnevna dinamika u biocenozama u pravilu je to izraženija što je veća razlika u temperaturi, vlažnosti i drugim čimbenicima okoliša između dana i noći. Značajnija odstupanja u biocenozama uočavaju se sezonskom dinamikom. To je zbog bioloških ciklusa organizama koji ovise o sezonskoj cikličnosti prirodnih pojava. Dakle, promjena godišnjeg doba značajno utječe na životnu aktivnost životinja i biljaka (hibernacija, zimski san, dijapauza i migracija kod životinja; razdoblja cvatnje, plodonošenja, aktivnog rasta, opadanja lišća i zimskog mirovanja kod biljaka). Slojevita struktura biocenoze često je podložna sezonskoj varijabilnosti. Pojedini slojevi biljaka mogu potpuno nestati u odgovarajućim godišnjim dobima, na primjer, zeljasti sloj koji se sastoji od jednogodišnjih biljaka. Trajanje bioloških godišnjih doba varira na različitim geografskim širinama. U tom smislu, sezonska dinamika biocenoza u arktičkim, umjerenim i tropskim zonama je različita. Najjasnije je izražen u ekosustavima umjerene klime i sjevernih geografskih širina. Dugoročna varijabilnost normalna je u životu svake biocenoze. Dakle, količina padalina koja pada u barabinskoj šumskoj stepi naglo varira iz godine u godinu; niz sušnih godina izmjenjuje se s dugotrajnim razdobljima obilnih oborina. To ima značajan utjecaj na biljke i životinje. U tom slučaju dolazi do razvoja ekoloških niša - funkcionalnog razgraničenja u novonastalom skupu ili njegovog dodavanja s malom raznolikošću. Dugoročne promjene u sastavu biocenoza također se ponavljaju u vezi s periodičnim promjenama opće cirkulacije atmosfere, što je pak uzrokovano povećanjem ili smanjenjem sunčeve aktivnosti. U procesu dnevne i sezonske dinamike, cjelovitost biocenoza obično nije narušena. Biocenoza doživljava samo periodične fluktuacije kvalitativnih i kvantitativnih karakteristika. Progresivne promjene u ekosustavu u konačnici dovode do zamjene jedne biocenoze drugom, s drugačijim skupom dominantnih vrsta. Razlozi za takve promjene mogu biti čimbenici izvan biocenoze koji djeluju dugo vremena u jednom smjeru, na primjer, sve veće onečišćenje vodenih tijela, sve veće isušivanje močvarnog tla kao rezultat melioracije, povećane ispaše itd. Te promjene iz jedne biocenoze u drugu nazivaju se egzogenetskim. U slučaju kada sve veći utjecaj čimbenika dovodi do postupnog pojednostavljivanja strukture biocenoze, osiromašenja njihovog sastava i smanjenja produktivnosti, takvi se pomaci nazivaju digresivnim ili digresivnim. Endogenetske promjene nastaju kao rezultat procesa koji se odvijaju unutar same biocenoze. Sukcesivna zamjena jedne biocenoze drugom naziva se ekološka sukcesija (od lat. - sukcesija - slijed, promjena). Sukcesija je proces samorazvoja ekosustava. Sukcesija se temelji na nedovršenosti biološkog ciklusa u određenoj biocenozi. Poznato je da živi organizmi, kao rezultat svoje vitalne aktivnosti, mijenjaju okoliš oko sebe, uklanjajući neke tvari iz njega i zasićujući ga metaboličkim produktima. Kada populacije postoje relativno dugo, mijenjaju svoj okoliš u nepovoljnom smjeru i, kao rezultat toga, bivaju istisnute populacijama drugih vrsta, za koje se rezultirajuće promjene okoliša pokazuju kao ekološki korisne. U biocenozi dolazi do promjene dominantnih vrsta. Ovdje je jasno vidljivo pravilo (princip) ekološkog udvostručavanja. Dugotrajno postojanje biocenoze moguće je samo ako su promjene u okolišu uzrokovane djelovanjem jednih živih organizama povoljne za druge sa suprotnim zahtjevima. Na temelju kompetitivnih interakcija vrsta tijekom sukcesije postupno se stvaraju stabilnije kombinacije koje odgovaraju specifičnim abiotičkim uvjetima okoliša. Primjer sukcesije koja dovodi do zamjene jedne zajednice drugom je zarastanje malog jezera praćeno pojavom močvare, a zatim šume na njegovom mjestu. Najprije se uz rubove jezera formira plutajući tepih - plutajući tepih od šaša, mahovina i drugog bilja. Jezero je stalno ispunjeno mrtvim biljnim ostacima – tresetom. Nastaje močvara, postupno obrasla šumom. Sukcesivni niz zajednica koje postupno i prirodno zamjenjuju jedna drugu u sukcesiji naziva se niz sukcesije. Sukcesije u prirodi vrlo su raznolike u opsegu. Mogu se uočiti u obalama s kulturama, koje su planktonske zajednice - razne vrste plutajućih algi i njihovi konzumenti - rotiferi, flagelati u lokvama i barama, močvarama, livadama, šumama, napuštenim obradivim površinama, trošnim stijenama itd. Postoji hijerarhija u organizaciji ekosustava Očituje se i u sukcesijskim procesima – veće transformacije biocenoza čine manje. U stabilnim ekosustavima s reguliranim ciklusom tvari stalno se odvijaju i lokalne sukcesivne promjene koje podržavaju složenu unutarnju strukturu zajednica. Vrste nasljednih promjena. Postoje dvije glavne vrste sukcesijskih promjena: 1 - uz sudjelovanje autotrofnih i heterotrofnih populacija; 2 - uz sudjelovanje samo heterotrofa. Sukcesija drugog tipa javlja se samo u uvjetima kada se stvara prethodna ili stalna zaliha organskih spojeva, zbog kojih postoji zajednica: u hrpama ili hrpama gnoja, u raspadajućoj biljnoj tvari, u rezervoarima zagađenim organskim tvarima itd. Proces sukcesije. Prema F. Clementsu (1916.), proces sukcesije sastoji se od sljedećih faza: Sukcesija sa smjenom vegetacije može biti primarna i sekundarna. Primarna sukcesija je proces razvoja i promjene ekosustava u prethodno nenaseljenim područjima, počevši od njihove kolonizacije. Klasičan primjer je stalno obraštavanje golih stijena s eventualnim razvojem šuma na njima. Dakle, u primarnim sukcesijama koje se javljaju na stijenama Uralskih planina, razlikuju se sljedeće faze: Sekundarna sukcesija je obnova ekosustava koji je nekoć postojao na određenom području. Počinje kada se uspostavljeni odnosi organizama u uspostavljenoj biocenozi poremete zbog vulkanske erupcije, požara, sječe, oranja itd. Pomaci koji dovode do obnove prethodne biocenoze nazivaju se u geobotanici demutacijskim. Primjer je dinamika raznolikosti vrsta na otoku Krakatoa nakon potpunog uništenja domaće flore i faune od strane vulkana. Drugi primjer je sekundarna sukcesija sibirske tamne crnogorične šume (tajga jela-cedar) nakon razornog šumskog požara. U opožarenijim područjima pionirske mahovine nastaju iz spora koje raznosi vjetar: 3-5 godina nakon požara najzastupljenija je „vatrena mahovina“ Funaria hygrometrica, Geratodon purpureus itd. Među višim biljkama vrlo brzo se pojavljuje ognjenica (Chamaenerion angustifolium). naseljava opožarena područja), koja već nakon 2-3 mjeseca u požaru obilno cvjeta, kao i mljevenu trstiku (Calamagrostis epigeios) i druge vrste. Promatraju se daljnje faze sukcesije: livada trske ustupa mjesto grmlju, zatim šuma breze ili jasike, mješovita šuma bora, šuma bora, šuma bora i cedra, te na kraju, nakon 250 godina, dolazi do obnove šume cedra i jele. . Sekundarne sukcesije u pravilu se odvijaju brže i lakše od primarnih, budući da se u narušenom staništu očuva profil tla, sjemenke, primordije i dio prethodne populacije i prijašnjih veza. Demutacija nije ponavljanje bilo koje faze primarne sukcesije. Climax ekosustav. Sukcesija završava fazom kada sve vrste ekosustava, dok se razmnožavaju, održavaju relativno konstantan broj i ne dolazi do daljnjih promjena u njihovom sastavu. Ovo ravnotežno stanje naziva se klimaks, a ekosustav klimaks. U različitim abiotskim uvjetima formiraju se različiti klimaksni ekosustavi. U vrućoj i vlažnoj klimi to će biti tropska prašuma, u suhoj i vrućoj klimi pustinja. Glavni biomi na zemlji su vrhunci ekosustava svojih odgovarajućih geografskih područja. Promjene u ekosustavu tijekom sukcesije. Produktivnost i biomasa. Kao što je već navedeno, sukcesija je prirodan, usmjeren proces i promjene koje se događaju u jednoj ili drugoj fazi karakteristične su za bilo koju zajednicu i ne ovise o njezinom sastavu vrsta ili zemljopisnom položaju. Postoje četiri glavne vrste nasljednih promjena: Također treba napomenuti da se promjena faza sukcesije odvija u skladu s određenim pravilima. Svaka faza priprema okolinu za pojavu sljedeće. Ovdje djeluje zakon slijeda prolaska razvojnih faza; faze razvoja prirodnog sustava mogu se nizati samo evolucijski utvrđenim (povijesno, okolišno uvjetovanim) redom, obično od relativno jednostavnih prema složenim, u pravilu, bez gubitka međufaza, ali vjerojatno s njihovim vrlo brzim prolaskom ili evolucijski fiksiranom odsutnošću. Kada se ekosustav približi stanju menopauze, u njemu se, kao i u svim ravnotežnim sustavima, usporavaju svi razvojni procesi. Ova situacija se odražava u zakonu sukcesijskog usporavanja: procesi koji se odvijaju u zrelim ravnotežnim ekosustavima koji su u stabilnom stanju, u pravilu imaju tendenciju usporavanja. U tom slučaju restorativni tip nasljeđivanja prelazi u njihov sekularni tijek, tj. samorazvoj se događa unutar granica menopauze ili nodalnog razvoja. Empirijski zakon usporavanja sukcesije posljedica je pravila G. Oduma i R. Pinkertona, odnosno pravila maksimalne energije za održanje zrelog sustava: sukcesija se odvija u smjeru temeljnog pomaka protoka energije prema povećanju njegove količina, usmjerena na održavanje sustava. Pravilo G. Oduma i R. Pinkertona, pak, temelji se na pravilu maksimalne energije u biološkim sustavima, koje je formulirao A. Lotka. Ovo su pitanje kasnije dobro razvili R. Margalef, Y. Odum i poznato je kao dokaz načela „nultog maksimuma“, ili minimiziranja rasta u zrelom ekosustavu: ekosustav u sukcesijskom razvoju teži stvaranju najveće biomase s najmanju biološku produktivnost. Lindeman (1942.) je eksperimentalno dokazao da je sukcesija popraćena povećanjem produktivnosti do zajednice klimaksa, u kojoj se pretvorba energije odvija najučinkovitije. Podaci iz istraživanja sukcesije hrastovih i jasenovih šuma pokazuju da u kasnijim fazama njihova produktivnost zapravo raste. Međutim, tijekom prijelaza u zajednicu klimaksa obično dolazi do smanjenja ukupne produktivnosti. Stoga je produktivnost u starim šumama niža nego u mladim šumama, koje zauzvrat mogu biti manje produktivne od zeljastih slojeva bogatijih vrstama koji im prethode. Slični padovi u produktivnosti primijećeni su u nekim vodenim sustavima. Nekoliko je razloga za to. Jedan od njih je da nakupljanje hranjivih tvari u rastućoj šumskoj biomasi može dovesti do smanjenja njihovog kruženja. Pad ukupne produktivnosti mogao bi jednostavno biti rezultat smanjene vitalnosti pojedinaca kako njihova prosječna dob u zajednici raste. Kako sukcesija napreduje, sve veći udio raspoloživih hranjivih tvari nakuplja se u biomasi zajednice, a sukladno tome njihov sadržaj u abiotskoj komponenti ekosustava (u tlu ili vodi) opada. Količina proizvedenog detritusa također se povećava. Glavni primarni potrošači nisu biljojedi, već detritivorni organizmi. Odgovarajuće promjene također se događaju u trofičkim mrežama. Detritus postaje glavni izvor hranjivih tvari. Tijekom sukcesije povećava se zatvorenost biogeokemijskih ciklusa tvari. Otprilike 10 godina od početka obnove vegetacijskog pokrova, otvorenost ciklusa se smanjuje sa 100 na 10%, a zatim se još više smanjuje, dostižući minimum u fazi klimaksa. Pravilo sve veće zatvorenosti biogeokemijskog ciklusa tvari tijekom sukcesije, može se pouzdano tvrditi, narušava se antropogenom transformacijom vegetacije i prirodnih ekosustava općenito. Nedvojbeno, to dovodi do dugog niza anomalija u biosferi i njezinim podjelama. Smanjenje raznolikosti vrsta tijekom menopauze ne znači i njen mali ekološki značaj. Raznolikost vrsta oblikuje sukcesiju, njen smjer i osigurava da stvarni prostor bude ispunjen životom. Nedovoljan broj vrsta koje čine kompleks ne bi mogao formirati sukcesivni niz, te bi postupno, s uništenjem klimaksnih ekosustava, došlo do potpune dezertifikacije planeta. Vrijednost raznolikosti je funkcionalna i statički i dinamički. Treba napomenuti da tamo gdje raznolikost vrsta nije dovoljna za formiranje biosfere, koja služi kao temelj normalnog prirodnog tijeka procesa sukcesije, a sam okoliš je oštro narušen, sukcesija ne doseže vrhunac faze, već završava nodalnom zajednicom – paraklimaksom, dugotrajnom ili kratkotrajnom izvedenom zajednicom. Što je dublji poremećaj okoliša pojedinog prostora, sukcesija ranijih faza prestaje. Kada se jedna ili skupina vrsta izgubi kao posljedica njihovog uništenja (antropogeni nestanak staništa, rjeđe izumiranje), postizanje menopauze nije potpuna obnova prirodnog okoliša. Zapravo, radi se o novom ekosustavu, jer su u njemu nastale nove veze, mnoge stare su izgubljene, a razvilo se drugačije "raspadanje" vrsta. Ekosustav se ne može vratiti u staro stanje, budući da se izgubljena vrsta ne može obnoviti. Kada se bilo koji abiotički ili biotički čimbenik promijeni, na primjer, s trajnim hlađenjem ili uvođenjem nove vrste, vrsta koja je slabo prilagođena novim uvjetima suočit će se s jednim od tri puta. Zakon evolucijsko-ekološke nepovratnosti kaže da se ekosustav koji je izgubio dio svojih elemenata ili je zamijenjen drugim kao rezultat neravnoteže ekoloških komponenti ne može vratiti u svoje prvobitno stanje tijekom sukcesije ako se tijekom promjena evolucijski (mikroevolucijski) ) došlo je do promjena ekoloških elemenata (očuvanih ili privremeno izgubljenih). U slučaju kada se neka vrsta izgubi u međufazama sukcesije, taj se gubitak može funkcionalno nadoknaditi, ali ne u potpunosti. Kada se raznolikost smanji preko kritične razine, tijek sukcesije je iskrivljen, i zapravo se ne može postići vrhunac identičan prošlosti. Za procjenu prirode obnovljenih ekosustava važan je zakon evolucijsko-ekološke nepovratnosti. Gubitkom elemenata, to su zapravo potpuno nove ekološke prirodne tvorevine s novonastalim uzorcima i vezama. Dakle, prijenos u prošlost vrste koja je tijekom reaklimatizacije ispala iz ekosustava ne znači i njezin mehanički povratak. To je zapravo uvođenje nove vrste u obnovljeni ekosustav.Zakon evolucijsko-ekološke nepovratnosti ističe smjer evolucije ne samo na razini biosustava, već i na svim ostalim hijerarhijskim razinama biote. 4 Biosfera kao globalni ekosustav koji osigurava cjelovitost i održivost ekosustava Biosfera je globalni ekosustav. Kao što je ranije navedeno, biosfera se dijeli na geobiosferu, hidrobiosferu i aerobiosferu (slika 2.4). Geobiosfera ima podjele u skladu s glavnim čimbenicima koji stvaraju okoliš: terabiosfera i litobiosfera - unutar geobiosfere, marinobiosfera (oceanobiosfera) i akvabiosfera - unutar hidrobiosfere. Te se formacije nazivaju subsferama. Vodeći okolišnotvorni čimbenik u njihovu nastanku je fizikalna faza životnog okoliša: zrak-voda u aerobiosferi, voda-slatkovodna i slana voda u hidrobiosferi, čvrsti zrak u terabiosferi i čvrsta voda u litobiosferi. Zauzvrat, svi oni padaju u slojeve: aerobiosfera - u tropobiosferu i altobiosferu; hidrobiosfera - na fotosferu, disfotosferu i afotosferu. Strukturotvorni čimbenici ovdje su, osim fizičkog okoliša, energija (svjetlost i toplina), posebni uvjeti za nastanak i razvoj života - evolucijski pravci prodiranja biote na kopno, u njegove dubine, u prostore iznad zemlja, bezdan oceana, nedvojbeno su različiti. Zajedno s apobiosferom, parabiosferom i drugim sub- i supra-biosferskim slojevima, oni čine takozvani „kolač života“ i geosferu (ekosferu) njegovog postojanja unutar granica megabiosfere. U sistemskom smislu, navedene tvorevine su veliki funkcionalni dijelovi gotovo univerzalnih ili subplanetarnih dimenzija. Znanstvenici vjeruju da u biosferi postoji najmanje 8-9 razina relativno neovisnih ciklusa tvari unutar međusobnog povezivanja 7 glavnih materijalno-energetskih ekoloških komponenti i 8. - informacijske. Globalni, regionalni i lokalni ciklusi tvari nisu zatvoreni i djelomično se "sijeku" unutar hijerarhije ekosustava. Ova materijalno-energetska, a dijelom i informacijska “sprega” osigurava cjelovitost ekoloških supersustava do biosfere u cjelini. Cjelovitost i održivost ekosustava. Biosfera se u većoj mjeri ne formira vanjskim čimbenicima, već unutarnjim obrascima. Najvažnije svojstvo biosfere je međudjelovanje živih i neživih bića, što se ogleda u zakonu biogene migracije atoma V. I. Vernadskog. Zakon biogene migracije atoma omogućuje čovječanstvu svjesno upravljanje biogeokemijskim procesima kako na Zemlji kao cjelini tako iu njezinim regijama. Količina žive tvari u biosferi, kao što je poznato, nije podložna značajnijim promjenama. Ovaj obrazac je formuliran u obliku zakona konstantnosti količine žive tvari od strane V.I. Vernadsky: količina žive tvari u biosferi za određeno geološko razdoblje je konstanta. U praksi, ovaj zakon je kvantitativna posljedica zakona unutrašnje dinamičke ravnoteže za globalni ekosustav – biosferu. Budući da je živa tvar, u skladu sa zakonom biogene migracije atoma, energetski posrednik između Sunca i Zemlje, njezina količina ili mora biti konstantna, ili se njezina energetska svojstva moraju mijenjati. Zakon fizičkog i kemijskog jedinstva žive tvari (sva živa tvar Zemlje je fizički i kemijski jedinstvena i isključuje značajne promjene u potonjem svojstvu. Stoga je kvantitativna stabilnost neizbježna za živu tvar planeta. Ona je u potpunosti karakteristična za broj vrsta. Živa tvar, kao akumulator sunčeve energije, mora istodobno reagirati i na vanjske (kozmičke) utjecaje i na unutarnje promjene. Smanjenje ili povećanje količine žive tvari na jednom mjestu biosfere trebalo bi dovesti do upravo suprotnog procesa na drugom mjestu, jer oslobođene hranjive tvari ostatak žive tvari može asimilirati ili će se uočiti njihov nedostatak. Ovdje moramo uzeti u obzir brzinu procesa koja je u slučaju antropogenih promjena mnogo manja od izravnog remećenja prirode od strane čovjeka. Osim postojanosti i postojanosti količine žive tvari, koja se ogleda u zakonu fizikalno-kemijskog jedinstva žive tvari, u živoj prirodi postoji stalno očuvanje informacijske i somatske strukture, unatoč tome što se ona mijenja. donekle s tijekom evolucije. To je svojstvo uočio Yu. Goldsmith (1981.) i nazvalo ga je zakon očuvanja strukture biosfere - informacijske i somatske ili prvi zakon ekodinamike. Kako bi očuvala strukturu biosfere, živa bića nastoje postići stanje zrelosti ili ekološku ravnotežu. Za biosferu i druge razine ekoloških sustava vrijedi zakon želje za menopauzom - drugi zakon ekodinamike Yu.Goldsmitha, iako ima specifičnosti - biosfera je zatvoreniji sustav od svojih pododjela. Jedinstvo žive tvari biosfere i homologija strukture njezinih podsustava dovode do činjenice da su živi elementi različitih geoloških dobi i izvornog geografskog podrijetla koji su nastali na njoj zamršeno evolucijski isprepleteni. Ispreplitanje elemenata različite prostorno-vremenske geneze na svim ekološkim razinama biosfere odražava pravilo ili princip heterogeneze žive tvari. Taj dodatak nije kaotičan, već je podložan načelima ekološke komplementarnosti, ekološke usklađenosti (kongruencije) i drugim zakonitostima. U okviru ekodinamike Yu.Goldsmitha, to je njezin treći zakon - princip ekološkog reda, ili ekološkog uzajamnosti, koji ukazuje na globalno svojstvo zbog utjecaja cjeline na njezine dijelove, obrnuti utjecaj diferenciranih dijelova na razvoj cjeline i sl. što ukupno dovodi do očuvanja stabilnosti biosfere kao cjeline. Uzajamno pomaganje u okviru ekološkog poretka, odnosno sistemskog uzajamnosti, potvrđuje zakon uređenosti popunjavanja prostora i prostorno-vremenske izvjesnosti: popunjavanje prostora unutar prirodnog sustava, zbog međudjelovanja njegovih podsustava, uređeno je u takav način koji omogućuje da se homeostatska svojstva sustava realiziraju uz minimalne kontradikcije između dijelova unutar njega. Iz ovog zakona proizlazi da je dugoročno postojanje nesreća koje su "nepotrebne" prirodi, uključujući i njoj tuđe ljudske tvorevine, nemoguće. Pravila uzajamnog uređenja sustava u biosferi uključuju i načelo komplementarnosti sustava, koje kaže da podsustavi jednog prirodnog sustava svojim razvojem daju preduvjet za uspješan razvoj i samoregulaciju ostalih podsustava uključenih u isti sustav. Četvrti zakon ekodinamike Yu. Goldsmitha uključuje zakon samokontrole i samoregulacije živih bića: živi sustavi i sustavi pod upravljačkim utjecajem živih bića sposobni su za samokontrolu i samoregulaciju u procesu svog prilagođavanje promjenama u okolini. U biosferi se samokontrola i samoregulacija odvijaju tijekom kaskadnih i lančanih procesa opće interakcije - tijekom borbe za postojanje prirodne selekcije (u najširem smislu ovog pojma), prilagodbe sustava i podsustava, široke koevolucije itd. Štoviše, svi ti procesi dovode do pozitivnih rezultata “sa stajališta” prirode - očuvanje i razvoj ekosustava biosfere i nje u cjelini. Spojna karika između generalizacija strukturalne i evolucijske prirode je pravilo automatskog održavanja globalnog staništa: živa tvar u tijeku samoregulacije i interakcije s abiotičkim čimbenicima autodinamički održava životni okoliš pogodan za svoj razvoj. Taj je proces ograničen promjenama u kozmičkoj i globalnoj ekosferi i događa se u svim ekosustavima i biosustavima planeta, kao kaskada samoregulacije koja doseže globalne razmjere. Pravilo automatskog održavanja globalnog staništa proizlazi iz biogeokemijskih načela V.I. Vernadskog, pravila za očuvanje staništa vrsta, relativnu unutarnju konzistentnost i služi kao konstanta prisutnosti konzervativnih mehanizama u biosferi i ujedno potvrđuje pravilo sustavno-dinamičke komplementarnosti. O kozmičkom utjecaju na biosferu svjedoči zakon prelamanja kozmičkih utjecaja: kozmički čimbenici, koji utječu na biosferu, a posebno na njezine dijelove, podložni su promjenama ekosfere planeta i stoga, u smislu snage i vremena , manifestacije mogu oslabiti i pomaknuti se ili čak potpuno izgubiti svoj učinak. Generalizacija je ovdje važna zbog činjenice da često postoji tok sinkronih učinaka sunčeve aktivnosti i drugih kozmičkih čimbenika na Zemljine ekosustave i organizme koji je nastanjuju. Valja napomenuti da se mnogi procesi na Zemlji i u njezinoj biosferi, iako podložni utjecaju svemira, pretpostavljaju cikluse Sunčeve aktivnosti s intervalima od 1850, 600, 400, 178, 169, 88, 83, 33, 22, 16. , 11, 5 (11 ,1), 6,5 i 4,3 godine, sama biosfera i njezini dijelovi ne moraju nužno reagirati istom cikličnosti u svim slučajevima. Kozmički utjecaji sustava biosfere mogu se potpuno ili djelomično blokirati. Zaključak Ekosustavi su osnovna funkcionalna cjelina u ekologiji, budući da uključuju organizme i neživi okoliš - komponente koje međusobno utječu na svojstva i nužne uvjete za održanje života u obliku kakav postoji na Zemlji. Kombinacija specifičnog fizikalno-kemijskog okoliša (biotopa) sa zajednicom živih organizama (biocenoza) čini ekosustav. Postoje mikroekosustavi, mezoekosustavi i globalni ekosustav – biosfera. Ekosustavi nisu raštrkani u neredu, naprotiv, grupirani su u prilično pravilne zone, vodoravno (po širini) i okomito (po visini). Primarni izvor energije za ekosustave je Sunce. Sunčevo zračenje koje pada na Zemljinu površinu iznosi 1,54 milijuna EJ godišnje. Većina sunčeve energije koja dospije na površinu planeta pretvara se izravno u toplinu, zagrijavajući vodu ili tlo, što zauzvrat zagrijava zrak. Ekosustavi postoje zahvaljujući nezagađujućoj i gotovo vječnoj sunčevoj energiji, čija je količina relativno stalna i izdašna. Sunčeva energija na Zemlji uzrokuje dva ciklusa tvari: veliki, ili geološki, i mali, biološki (biotički). Oba ciklusa su međusobno povezana i predstavljaju takoreći jedan proces. Postojanje biogeokemijskih ciklusa, odnosno biogeokemijskih ciklusa, stvara mogućnost samoregulacije (homeostaze) sustava, što ekosustavu daje stabilnost: nevjerojatnu konstantnost postotnog sadržaja različitih elemenata. Postoji dosta ciklusa tvari. Najznačajniji ciklus na Zemlji u pogledu prenesenih masa i potrošnje energije je planetarni hidrološki ciklus ili ciklus vode. Biotički (biološki) ciklus odnosi se na kruženje tvari između tla, biljaka, životinja i mikroorganizama. Od svih biogeokemijskih ciklusa, ciklus ugljika je bez sumnje najintenzivniji. Većina proizvedenog kisika tijekom geoloških epoha nije ostala u atmosferi, već ga je litosfera fiksirala u obliku karbonata, sulfata, željeznih oksida itd. Ova masa je 590x1014 tona naspram 39x1014 tona kisika, koji cirkulira u biosferi u obliku plina ili sulfata otopljenih u kontinentalnim i oceanskim vodama. Dušik je esencijalni biogeni element, jer je dio proteina i nukleinskih kiselina. Ciklus dušika je jedan od najsloženijih, jer uključuje i plinsku i mineralnu fazu, a ujedno i najidealnije cikluse. Ciklus fosfora u biosferi povezan je s metaboličkim procesima u biljkama i životinjama. Ovaj važan i neophodan element protoplazme, sadržan u kopnenim biljkama i algama 0,01-0,1%, životinje od 0,1% do nekoliko posto, cirkulira, postupno se pretvarajući iz organskih spojeva u fosfate, koje biljke opet mogu koristiti. Postoje brojni plinoviti spojevi sumpora, kao što su sumporovodik H2S i sumporov dioksid SO2. Održavanje vitalne aktivnosti organizama i kruženje tvari u ekosustavima, odnosno postojanje ekosustava, ovisi o stalnom protoku energije potrebne svim organizmima za njihov život i samorazmnožavanje. Za razliku od tvari koje kontinuirano kruže kroz različite blokove ekosustava, a koje se uvijek mogu ponovno upotrijebiti i ući u ciklus, energija se može upotrijebiti jednokratno, odnosno postoji linearan tok energije kroz ekosustav. Unutar ekosustava, tvari koje sadrže energiju stvaraju autotrofni organizmi i služe kao hrana za heterotrofe. Prehrambene veze su mehanizmi za prijenos energije iz jednog organizma u drugi. Unutar svakog ekosustava, hranidbene mreže imaju dobro definiranu strukturu koju karakteriziraju priroda i broj organizama zastupljenih na svakoj razini različitih prehrambenih lanaca. Formiranje ekosustava je dinamičan proces. U ekosustavima se stalno događaju promjene u stanju i životnoj aktivnosti njihovih članova te u omjeru populacija. Različite promjene koje se događaju u bilo kojoj zajednici mogu se podijeliti u dvije glavne vrste: cikličke i progresivne. Sukcesija je prirodan, usmjeren proces i promjene koje se događaju u jednoj ili drugoj fazi karakteristične su za svaku zajednicu i ne ovise o njezinom sastavu vrsta ili geografskom položaju. Globalni ekosustav je biosfera. Kao što je ranije navedeno, biosfera se dijeli na geobiosferu, hidrobiosferu i aerobiosferu. Biosfera se u većoj mjeri ne formira vanjskim čimbenicima, već unutarnjim obrascima. Najvažnije svojstvo biosfere je međudjelovanje živih i neživih bića, što se odražava u zakonu biogene migracije atoma V.I. Vernadski. Ekosustavi koji postoje na Zemlji su raznoliki. Postoje mikroekosustavi (na primjer, deblo trulog drveta), mezoekosustavi (šuma, ribnjak itd.), makroekosustavi (kontinent, ocean itd.) i globalna biosfera. Veliki kopneni ekosustavi nazivaju se biomi. Svaki biom sadrži niz manjih, međusobno povezanih ekosustava. Postoji nekoliko klasifikacija ekosustava: zimzelene tropske kišne šume; pustinja: travnata i grmljasta; tropski travnjaci i savane; umjerena stepa; listopadne šume umjerenog pojasa: borealne crnogorične šume. Tundra: arktička i alpska. Vrste slatkovodnih ekosustava: vrpca (stojeće vode): jezera, bare itd.; lotic (vode koje teku): rijeke, potoci itd.; močvare: močvare i močvarne šume; Vrste morskih ekosustava: otvoreni ocean (pelagikal); vode epikontinentalnog pojasa (obalne vode); područja uzdizanja (plodna područja s produktivnim ribolovom); estuari (obalni zaljevi, tjesnaci, riječna ušća, slane močvare itd.). Svaki ekosustav ima dvije glavne komponente: organizme i čimbenike iz njihove nežive okoline. Ukupnost organizama (biljke, životinje, mikrobi) naziva se biota ekosustava. Načini interakcije između različitih kategorija organizama su njegova biotička struktura. Sunčeva energija na Zemlji uzrokuje dva ciklusa tvari: veliki, ili geološki, koji se najjasnije očituje u ciklusu vode i kruženju atmosfere, i mali, biološki (biotički), koji se razvija na temelju velikog i sastoji se od kontinuiranog, ciklički, ali neujednačeni u vremenu i prostoru, te praćeni manje ili više značajnim gubicima u prirodnoj preraspodjeli tvari, energije i informacija unutar ekoloških sustava različitih razina organizacije. Oba ciklusa su međusobno povezana i predstavljaju takoreći jedan proces. Međudjelovanje abiotskih čimbenika i živih organizama ekosustava prati kontinuirano kruženje tvari između biotopa i biocenoze u obliku izmjeničnih organskih i mineralnih spojeva. Razmjena kemijskih elemenata između živih organizama i anorganskog okoliša, čije se različite faze odvijaju unutar ekosustava, naziva se biogeokemijski ciklus ili biogeokemijski ciklus. Postojanje takvih ciklusa stvara mogućnost samoregulacije (homeostaze) sustava, što ekosustavu daje stabilnost: nevjerojatnu konstantnost postotka različitih elemenata. Ovdje vrijedi princip funkcioniranja ekosustava: stjecanje resursa i odlaganje otpada odvija se u okviru ciklusa svih elemenata. Popis korištene literature 1.Bigon M.I. Ekologija, pojedinci, populacije i zajednice. M.: 1989. - 290 str. 2.Beysenova A.S., Shildebaev Zh.B., Sautbaeva G.Z. Ekologija. Almaty: "Gylym, 2001. - 201 str. 3.Akimova T.A., Khaskin V.V. Ekologija. M.: Izdavačka kuća UNITY, 1998. - 233 str. .Gorelov A.A. Ekologija. Tečaj predavanja. M.: "Centar" 1997. - 280 str. .Stadnitsky G.V., Rodionov A.I. Ekologija. M.: Izdavačka kuća UNITY, 1996. - 272 str. .Sadanov A.K., Svanbaeva Z.S., Ekologija. Almaty: "Agrouniversity", 1999. - 197 str. .Shilov I.A. Ekologija. M.: Viša škola, 2000. - 348 str. .Reimers N.F. Ekologija (Teorija, zakoni, pravila, principi i hipoteze). M.: "Mlada Rusija", 1994. - 260 str. .Tsvetkova L.I., Alekseev M.I., Usenov B.P. i dr. Ekologija. Udžbenik za tehnička sveučilišta. M.; ASV, St. Petersburg: Khimizdat, 1999., 185 str. .Girusov E.V. Ekologija i ekonomika upravljanja okolišem. M.: Pravo i pravo, JEDINSTVO. 1998. - 232 str. .Vronski V.A. Primijenjena ekologija: Udžbenik. Korist. Rostov na Donu: 1995. - 197 str. .Budyko M.I. Globalna ekologija. M.: Mysl, 1977.-248 str. .Alekseenko V.A. Geokemija okoliša. Udžbenik. M.: Logos, 2000. - 410 str. .Petrov K.M. Opća ekologija S-P. Tutorial. "Kemija", 1997. - 218 str. .Andersen J.M. Ekologija i znanost o okolišu. Biosfera, ekosustav, čovjek. L., 1985. 376 str. .Girenok F.I. Ekologija. Civilizacija. Noosfera.M.: 191987. - 281 str. .U potrazi za ravnotežom: ekologija u sustavu društveno-političkih prioriteta M.: 1992. - 427 str. .Povijest interakcije društva i prirode: Čimbenici i koncept. M.: ANSSSR 1990. .Stepanovskikh A.S. Primijenjena ekologija: Zaštita okoliša. Udžbenik za sveučilišta. M.: JEDINSTVO DANA. 2003. 751 str. Slični radovi - Kruženje tvari i protok energije u ekosustavima
Treba jasno definirati pojmove “protok materije” i “protok energije”. Tijek tvari je kretanje u obliku kemijskih elemenata i njihovih spojeva od proizvođača do razlagača (kroz ili bez potrošača). Protok energije je prijenos energije u obliku kemijskih veza organskih spojeva (hrane) duž prehrambenih lanaca s jedne trofičke razine na drugu (višu).
Treba napomenuti da, za razliku od tvari koje stalno cirkuliraju kroz različite blokove ekosustava i uvijek mogu ponovno ući u ciklus, dolazna energija može se koristiti samo jednom.
Kao univerzalni prirodni fenomen, jednosmjerni tok energije određen je zakonima termodinamike. Prema prvom zakonu, energija može prelaziti iz jednog oblika (energija svjetlosti) u drugi (potencijalna energija hrane), ali se više nikada ne stvara i ne nestaje bez traga.
Drugi zakon termodinamike kaže da ne može postojati niti jedan proces povezan s transformacijom energije bez gubitka dijela energije. Iz tog razloga ne može doći do transformacije sa 100% učinkovitosti npr. hrane u tvar od koje se sastoji tijelo organizma.
Dakle, funkcioniranje svih ekosustava određeno je stalnim protokom energije, koja je neophodna svim organizmima za održavanje njihovog postojanja i samoreprodukciju.
U ekosustavima postoje i konkurentski odnosi. S tog aspekta od velikog je interesa zakon maksimizacije energije (G. Odum - E. Odum): u konkurenciji s drugim ekosustavima opstaje onaj koji najbolje pridonosi opskrbi energijom i na najučinkovitiji način koristi njezinu najveću količinu. (konzervira). Sukladno zakonu, u tu svrhu sustav: 1) stvara akumulatore (skladišta) visokokvalitetne energije (na primjer, rezerve masti); 2) troši određenu količinu akumulirane energije da osigura opskrbu novom energijom; 3) osigurava kruženje raznih tvari; 4) stvara regulatorne mehanizme koji podržavaju stabilnost sustava i njegovu sposobnost prilagodbe promjenjivim uvjetima; 5) uspostavlja razmjene s drugim sustavima potrebne za zadovoljenje potreba za posebnim vrstama energije.
Potrebno je istaknuti važnu okolnost: zakon maksimizacije energije vrijedi i u odnosu na informaciju, stoga se (prema N. F. Reimersu) može smatrati i zakonom maksimizacije energije i informacija: sustav koji najviše doprinosi primanje, proizvodnja i učinkovito korištenje energije i informacija.
Napomenimo da maksimalna opskrba tvari, kao takva, još ne jamči uspjeh sustava u konkurentskoj skupini drugih sličnih sustava.
Prethodno je napomenuto da između organizama biocenoze nastaju i uspostavljaju se jaki hranidbeni odnosi ili hranidbeni lanac. Potonji se sastoji od tri glavne karike: proizvođača, potrošača i razlagača.
Lanci ishrane koji započinju fotosintetskim organizmima nazivaju se lancima ispaše (ili ispaše), a lanci koji počinju mrtvom biljnom tvari, lešinama i životinjskim izmetom nazivaju se lancima detrita.
Položaj svake karike u hranidbenom lancu naziva se trofička razina; karakteriziraju ga različiti intenziteti protoka tvari i energije. Prva trofička razina uvijek su proizvođači; drugi - potrošači biljojedi; treći - mesožderi, koji žive od biljojeda; četvrta razina - konzumiranje drugih mesoždera itd.
Postoje konzumenti prvog, drugog, trećeg i četvrtog reda koji zauzimaju različite razine u hranidbenom lancu (slika 9).
Riža. 9.
Očito je da veliku ulogu u tome igra prehrambena specijalizacija potrošača. Vrste sa širokim rasponom prehrane mogu se uključiti u prehrambene lance na različitim trofičkim razinama. Prehrana, na primjer, osobe uključuje i biljnu hranu i meso biljojeda i mesojeda. Stoga djeluje u različitim prehrambenim lancima kao potrošač I, II ili III reda.
Budući da se energija gubi pri prijenosu energije s jedne razine na drugu, energetski lanac ne može biti dug: obično se sastoji od 4...6 karika (Tablica 1).
1. Tipični dijagrami prehrambenih lanaca (prema V. M. Ivoninu, 1996.)
Međutim, takvi se lanci u svom čistom obliku obično ne nalaze u prirodi, jer ista vrsta može biti prisutna istovremeno u različitim vezama. To je zbog činjenice da u prirodi ima malo monofaga, mnogo su češći oligofagi i polifagi. Na primjer, grabežljivci koji se hrane raznim biljojedima i mesojedima karike su u mnogim lancima. Kao rezultat toga, u svakoj biocenozi evolucijski se formiraju kompleksi hranidbenih lanaca, koji predstavljaju jednu cjelinu. Tako nastaju elektroenergetske mreže koje su vrlo složene.
Dakle, možemo zaključiti da je hranidbeni lanac glavni kanal prijenosa energije u zajednici (između biljaka – proizvođača, životinja – konzumenata i mikroorganizama – razlagača) (sl. 10.).Već na dijagramu je jasno da je ideja o lanci ishrane i trofičke razine prilično su apstrakcija. Linearni lanac s jasno odvojenim razinama može se stvoriti u laboratoriju. Ali u prirodi doista postoje trofičke mreže u kojima mnoge populacije pripadaju nekoliko trofičkih razina odjednom. Isti organizam troši i životinje i biljke; grabežljivac se može hraniti potrošačima prvog i drugog reda; mnoge životinje jedu i žive i mrtve biljke.
Zbog složenosti trofičkih odnosa, gubitak jedne vrste često nema gotovo nikakav učinak na zajednicu. Drugi “korisnici” počinju konzumirati hranu izumrlih vrsta,

Riža. 10.
vrste koje se njime hrane pronalaze nove izvore hrane: općenito se u zajednici održava ravnoteža.
Energija koju apsorbiraju proizvođači teče kroz prehrambene lance i postupno se troši. Na kraju hranidbenog lanca količina energije je uvijek manja nego na početku. Tijekom fotosinteze biljke vežu u prosjeku samo oko 1% sunčeve energije koja ih pogodi. Životinja koja je pojela biljku ne probavlja dio hrane i izbacuje je u obliku izmeta. Obično se probavi 20...60% biljne hrane; Apsorbirana energija koristi se za održavanje života životinje. Funkcioniranje stanica i organa popraćeno je oslobađanjem topline, pri čemu se znatan dio energije iz hrane ubrzo rasipa u okoliš. Relativno mali dio hrane koristi se za izgradnju novog tkiva i stvaranje masnih rezervi. Dalje, grabežljivac koji je pojeo biljojeda i predstavlja treću trofičku razinu dobiva samo onu energiju iz energije koju je akumulirala biljka koja se zadržava u tijelu njenog plijena (druga razina) u obliku povećanja biomase.
Poznato je da se u svakoj fazi prijenosa tvari i energije u hranidbenom lancu gubi približno 90% energije, a tek oko desetina odlazi do sljedećeg potrošača, odnosno prijenosa energije u prehrambenim vezama. organizama poštuje "pravilo deset posto" (Lindemannovo načelo). Na primjer, količina energije koja dopire do tercijarnih mesoždera (peta trofična razina) iznosi samo oko 10 -4 energije koju apsorbiraju proizvođači. Time se objašnjava ograničeni broj (5...6) karika (razina) u hranidbenom lancu, bez obzira na složenost sastava vrsta biocenoze.

Riža. jedanaest.
S obzirom na tok energije u ekosustavima, također je lako razumjeti zašto se biomasa smanjuje kako se trofička razina povećava. Ovdje se očituje treći temeljni princip funkcioniranja ekosustava: što je veća biomasa populacije, to bi trebala biti niža trofička razina koju zauzima, ili inače: na kraju dugih hranidbenih lanaca ne može biti velika biomasa.
Tri osnovna načela funkcioniranja ekosustava navedena gore - ciklus hranjivih tvari, protok sunčeve energije i smanjenje biomase s povećanjem trofičke razine - mogu se prikazati u obliku općeg dijagrama (slika 11). Posložimo li organizme prema njihovim hranidbenim odnosima, navodeći za svaki od njih “ulaz” i “izlaz” energije i hranjivih tvari, postaje očito da se hranjive tvari kontinuirano recikliraju unutar ekosustava, a tok energije prolazi kroz njega.
Održavanje vitalne aktivnosti organizama i kruženje tvari u ekosustavima, odnosno postojanje ekosustava, ovisi o stalnom protoku energije potrebne svim organizmima za njihov život i samorazmnožavanje.
Za razliku od tvari koje kontinuirano kruže kroz različite blokove ekosustava, a koje se uvijek mogu ponovno upotrijebiti i ući u ciklus, energija se može upotrijebiti samo jednom, odnosno postoji linearan tok energije kroz ekosustav.
Jednosmjerni dotok energije kao univerzalni prirodni fenomen nastaje kao posljedica zakona termodinamike. Prvi zakon navodi da se energija može pretvoriti iz jednog oblika (kao što je svjetlost) u drugi (kao što je potencijalna energija hrane), ali se ne može stvoriti ili uništiti. Drugi zakon navodi da ne može postojati niti jedan proces povezan s transformacijom energije a da se dio nje ne izgubi. Određena količina energije u takvim transformacijama se rasipa u nedostupnu toplinsku energiju i time se gubi. Dakle, ne može doći do transformacija npr. prehrambenih tvari u tvari koje čine tijelo organizma, a koje se događaju sa 100 postotnom učinkovitošću.
Dakle, živi organizmi su pretvarači energije. I svaki put kada se energija pretvara, dio se gubi u obliku topline. U konačnici, sva energija koja ulazi u biotski ciklus ekosustava rasipa se kao toplina. Živi organizmi zapravo ne koriste toplinu kao izvor energije za rad - oni koriste svjetlost i kemijsku energiju.
Ekološke piramide. Unutar svakog ekosustava, hranidbene mreže imaju dobro definiranu strukturu koju karakteriziraju priroda i broj organizama zastupljenih na svakoj razini različitih prehrambenih lanaca. Kako bi proučavali odnose između organizama u ekosustavu i kako bi ih grafički prikazali, obično koriste ekološke piramide, a ne dijagrame mreže hrane. Ekološke piramide izražavaju trofičku strukturu ekosustava u geometrijskom obliku. Izrađuju se u obliku pravokutnika iste širine, ali duljina pravokutnika mora biti proporcionalna vrijednosti objekta koji se mjeri. Odavde možete dobiti piramide brojeva, biomase i energije.
Ekološke piramide odražavaju temeljne značajke bilo koje biocenoze kada prikazuju njezinu trofičku strukturu:
Njihova je visina proporcionalna duljini dotičnog hranidbenog lanca, tj. broju trofičkih razina koje sadrži;
Njihov oblik manje-više odražava učinkovitost energetskih transformacija tijekom prijelaza s jedne razine na drugu.
Piramide brojeva. Oni predstavljaju najjednostavniju aproksimaciju proučavanja trofičke strukture ekosustava. U tom slučaju prvo se broji broj organizama na određenom području, grupira ih po trofičkim razinama i prikazuje u obliku pravokutnika čija je duljina (ili površina) proporcionalna broju organizama koji žive na određenom području ( ili u određenom volumenu, ako se radi o vodenom ekosustavu). Utvrđeno je osnovno pravilo da u svakom okolišu ima više biljaka nego životinja, više biljojeda nego mesojeda, više insekata nego ptica itd.
Populacijske piramide odražavaju gustoću organizama na svakoj trofičkoj razini. Velika je raznolikost u izgradnji raznih populacijskih piramida. Često su naopačke.
Na primjer, u šumi ima znatno manje drveća (primarnih proizvođača) nego kukaca (biljojeda).
Piramida biomase. Potpunije odražava prehrambene odnose u ekosustavu, budući da uzima u obzir ukupnu masu organizama (biomasa) svakoj trofičkoj razini. Pravokutnici u piramidama biomase predstavljaju masu organizama na svakoj trofičkoj razini po jedinici površine ili volumena. Oblik piramide biomase često je sličan obliku piramide stanovništva. Karakteristično je smanjenje biomase na svakoj sljedećoj trofičkoj razini.
Piramide biomase, kao i brojevi, mogu biti ne samo ravne, već i obrnute. Obrnute piramide biomase karakteristične su za vodene ekosustave u kojima se primarni proizvođači, poput fitoplanktonskih algi, vrlo brzo dijele, a njihovi potrošači - zooplanktonski rakovi - puno su veći, ali imaju dug ciklus razmnožavanja. Osobito se to odnosi na slatkovodne okoliše, gdje primarnu produktivnost osiguravaju mikroskopski organizmi čija je stopa metabolizma povećana, tj. biomasa je niska, produktivnost visoka.
Piramida energije. Najosnovniji način prikazivanja veza između organizama na različitim trofičkim razinama je kroz energetske piramide. Oni predstavljaju učinkovitost pretvorbe energije i produktivnost hranidbenih lanaca i konstruirani su brojanjem količine energije (kcal) akumulirane po jedinici površine po jedinici vremena i koju koriste organizmi na svakoj trofičkoj razini. Stoga je relativno lako odrediti količinu energije pohranjene u biomasi, ali je teže procijeniti ukupnu količinu energije apsorbirane na svakoj trofičkoj razini. Konstruirajući graf (sl. 12.28), možemo ustvrditi da destruktori, čija se važnost čini malom u piramidi biomase, i obrnuto u piramidi stanovništva; primaju značajan dio energije prolazeći kroz ekosustav. Štoviše, samo dio sve te energije ostaje u organizmima na svakoj trofičkoj razini ekosustava i pohranjuje se u biomasu, a ostatak se koristi za zadovoljenje metaboličkih potreba živih bića: održavanje postojanja, rast, reprodukcija. Životinje također troše značajnu količinu energije za rad mišića.
R. Lindeman je prvi put formulirao 1942 zakon energetske piramide, koji se u udžbenicima često naziva "zakon od 10%". Prema ovom zakonu od jedne trofička razina ekološke piramide U prosjeku, ne više od 10% energije prelazi na drugu razinu.
Potrošači služe kao upravljačka i stabilizirajuća karika u ekosustavu. Potrošači stvaraju spektar raznolikosti u cenozi, sprječavajući monopol dominantnih. Pravilo kontrole vrijednosti potrošača s pravom se može smatrati prilično temeljnim. Prema kibernetičkim pogledima, upravljački sustav trebao bi biti složeniji po strukturi od upravljanog, tada postaje jasan razlog mnoštva tipova potrošača. Kontrolni značaj potrošača ima i energetsku osnovu. Protok energije kroz jednu ili drugu trofičku razinu ne može se apsolutno odrediti dostupnošću hrane u temeljnoj trofičkoj razini. Kao što je poznato, uvijek ostaje dovoljna “rezerva”, jer bi potpuno uništenje hrane dovelo do smrti potrošača. Ovi opći obrasci promatraju se u okviru populacijskih procesa, zajednica, razina ekološke piramide i biocenoza u cjelini.
Kako biste koristili preglede prezentacije, stvorite Google račun i prijavite se na njega: https://accounts.google.com
Naslovi slajdova:
Kruženje materije i energije u prirodi
Kruženje tvari su ponavljajući procesi preobrazbe i kretanja tvari u prirodi, koji su više ili manje ciklični. Sve tvari na našem planetu su u procesu kruženja. Postoje dva glavna ciklusa u prirodi: Veliki (geološki) Mali (biogeokemijski)
Veliki ciklus tvari Veliki ciklus traje milijunima godina i uzrokovan je međudjelovanjem sunčeve energije s dubinskom energijom Zemlje. Povezano s geološkim procesima, stvaranjem i razaranjem stijena i naknadnim kretanjem produkata razaranja.
Mali ciklus tvari Mali ciklus (biogeokemijski) događa se unutar biosfere, na razini biocenoze. Njegova bit je nastajanje žive tvari iz anorganskih spojeva tijekom procesa fotosinteze i pretvaranje organske tvari tijekom razgradnje natrag u anorganske spojeve. Biogeokemijski ciklusi – Vernadsky V.I.
Ciklus vode Tr otjecanje inf Isparavanje vode Kondenzacija pare Otjecanje oborina Transpiracija infiltracija
Transpiracija je proces kretanja vode kroz biljku i njezino isparavanje kroz vanjske organe biljke, kao što su listovi, stabljike i cvjetovi. Voda je neophodna za život biljaka, ali samo mali dio vode dobavljene kroz korijenje koristi se izravno za rast i metabolizam.
Kruženje vode
Kruženje vode Većina vode koncentrirana je u oceanima. Voda koja isparava s njihove površine opskrbljuje prirodne i umjetne kopnene ekosustave. Što je neko područje bliže oceanu, to je više oborina. Zemlja neprestano vraća vodu u ocean: dio vlage isparava, najaktivnije u šumama, dio skupljaju rijeke: primaju kišnicu i otopljenu vodu. Izmjena vlage između oceana i kopna zahtijeva vrlo visoke troškove energije: otprilike 30% sunčeve energije koja dospije na Zemlju troši se na to.
Utjecaj čovjeka na kruženje vode Kruženje vode u biosferi prije razvoja civilizacije bilo je u ravnoteži, tj. ocean je dobio onoliko vode iz rijeka koliko je potrošio isparavanjem. S razvojem civilizacije taj se ciklus počeo remetiti. Konkretno, šume isparavaju sve manje vode jer... njihova površina se smanjuje, a površina tla, naprotiv, raste, jer Povećava se površina poljoprivrednog zemljišta koje se navodnjava. zemlje. Rijeke južnih krajeva postale su plitke. Voda lošije isparava s površine oceana, jer... značajan dio prekriven je uljnim slojem. Sve to pogoršava opskrbu biosfere vodom.
Suše su sve češće, a pojavljuju se i džepovi ekoloških katastrofa. Na primjer, katastrofalna suša traje više od 35 godina u Africi, u zoni Sahela - polupustinjskom području koje odvaja Saharu od sjevernih zemalja kontinenta. Slatka voda koja se s kopna vraća u ocean i druge vodene površine često je zagađena. Voda mnogih ruskih rijeka postala je praktički neprikladna za piće. Udio svježe vode dostupan živim organizmima je vrlo mali, stoga se mora koristiti štedljivo i ne zagađivati! Svaka četvrta osoba na planeti nema čistu pitku vodu. U mnogim dijelovima svijeta nema dovoljno vode za industrijsku proizvodnju i navodnjavanje.
Različite komponente hidrosfere sudjeluju u ciklusu vode na različite načine i različitim brzinama. Za potpunu obnovu vode u ledenjacima potrebno je 8000 godina, podzemnih voda – 5000 godina, oceana – 3000 godina, tla – 1 godina. Atmosferska para i riječne vode potpuno se obnavljaju za 10-12 dana. Kruženje vode u prirodi traje oko milijun godina.
Ciklus kisika Kisik je dio najčešćih elemenata u biosferi. Sadržaj kisika u atmosferi je gotovo 21%. Kisik je dio molekula vode i živih organizama (proteini, masti, ugljikohidrati, nukleinske kiseline). Kisik proizvode proizvođači (zelene biljke). Ozon ima važnu ulogu u ciklusu kisika. Ozonski omotač se nalazi na nadmorskoj visini od 20-30 km iznad razine mora. Na sadržaj kisika u atmosferi utječu 2 glavna procesa: 1) fotosinteza 2) razgradnja organske tvari, tijekom koje se ona troši.
Ciklus kisika je spor proces. Potrebno je oko 2000 godina da se sav kisik u atmosferi potpuno obnovi. Za usporedbu: potpuna obnova ugljičnog dioksida u atmosferi događa se za oko 3 godine. Većina živih organizama koristi kisik za disanje. Kisik se koristi pri izgaranju goriva u motorima s unutarnjim izgaranjem, u ložištima termoelektrana, u zrakoplovnim i raketnim motorima itd. Dodatna antropogena potrošnja može poremetiti ravnotežu ciklusa kisika. Do sada biosfera nadoknađuje ljudsku intervenciju: gubitke nadoknađuju zelene biljke. S daljnjim smanjenjem šumskih površina i sagorijevanjem sve više goriva, sadržaj kisika u atmosferi počet će se smanjivati.
TO JE VAŽNO!!! Kada se sadržaj kisika u zraku smanji na 16%, čovjekovo zdravlje se pogoršava (osobito strada srce), na 7% osoba gubi svijest, a na 3% nastupa smrt.
Ciklus ugljika
Ciklus ugljika Ugljik je osnova organskih spojeva, ulazi u sastav svih živih organizama u obliku bjelančevina, masti i ugljikohidrata. Ugljik ulazi u atmosferu u obliku ugljičnog dioksida. U atmosferi, gdje je koncentriran najveći dio ugljičnog dioksida, stalno se odvija izmjena: biljke apsorbiraju ugljični dioksid tijekom fotosinteze, a svi organizmi ga oslobađaju tijekom disanja. Do 50% ugljika u obliku CO 2 vraćaju u atmosferu razlagači – mikroorganizmi tla. Ugljik napušta ciklus kao kalcijev karbonat.
Utjecaj čovjeka na kruženje ugljika Tehnogeno djelovanje čovjeka remeti prirodnu ravnotežu kruženja ugljika: 1) pri izgaranju organskog goriva godišnje se u atmosferu emitira oko 6 milijardi tona CO 2: a) Proizvodnja električne energije u termoelektranama b ) Ispušni plinovi automobila 2) uništavanje šuma. Tijekom proteklih 100 godina razina ugljičnog dioksida u atmosferi stalno i brzo raste. Ugljični dioksid + metan + vodena para + ozon + dušikovi oksidi = staklenički plin. Posljedica je efekt staklenika - globalno zatopljenje, što može dovesti do prirodnih katastrofa velikih razmjera.
Kruženje dušika U slobodnom obliku dušik je sastavni dio zraka - 78%. Dušik je jedan od najvažnijih elemenata za život organizama. Dušik je dio svih proteina. Molekula dušika je vrlo jaka, zbog čega većina organizama ne može apsorbirati atmosferski dušik. Dušik apsorbiraju živi organizmi samo u obliku spojeva s vodikom i kisikom. Do fiksacije dušika u kemijske spojeve dolazi kao posljedica vulkanske i grmljavinske aktivnosti, ali najviše kao posljedica aktivnosti mikroorganizama – fiksatora dušika (bakterije koje fiksiraju dušik i modrozelene alge).
Dušik ulazi u korijenje biljaka u obliku nitrata, koji se koriste za sintezu organske tvari (proteina). Životinje unose dušik putem biljne ili životinjske hrane. Dušik se vraća u atmosferu razgradnjom mrtvog organskog materijala. Bakterije u tlu razlažu bjelančevine na anorganske tvari – plinove – amonijak, dušikove okside, koji ulaze u atmosferu. Dušik koji ulazi u vodena tijela također prolazi kroz hranidbeni lanac biljka-životinja-mikroorganizam i vraća se u atmosferu.
Ljudski utjecaj na kruženje dušika Tehnogene ljudske aktivnosti remete prirodnu ravnotežu kruženja dušika. Prilikom oranja zemljišta aktivnost mikroorganizama - fiksatora dušika smanjuje se gotovo 5 puta, stoga se smanjuje sadržaj dušika u tlu, što dovodi do smanjenja plodnosti tla. Stoga ljudi unose višak nitrata u tlo, koji su uključeni u mineralna gnojiva. Velika količina dušikovih oksida ulazi u atmosferu prilikom izgaranja i prerade plina, nafte i ugljena te pada u obliku kiselih kiša. Ponovno uspostavljanje prirodnog ciklusa dušika moguće je smanjenjem proizvodnje dušičnih gnojiva, smanjenjem industrijskih emisija dušikovih oksida u atmosferu itd.
Ciklus fosfora
Za razliku od ciklusa vode, ugljika, dušika i kisika, koji su zatvoreni, ciklus fosfora je otvoren, jer fosfor ne stvara hlapljive spojeve koji se ispuštaju u atmosferu. Fosfor se nalazi u stijenama, odakle ulazi u ekosustave tijekom prirodnog razaranja stijena ili prilikom primjene fosfornih gnojiva na polja. Biljke apsorbiraju anorganske spojeve fosfora, a životinje koje se hrane tim biljkama nakupljaju fosfor u svojim tkivima. Nakon razgradnje mrtvih tijela životinja i biljaka, nije sav fosfor uključen u ciklus. Dio se ispire iz tla u vodene površine (rijeke, jezera, mora) i taloži se na dno. Fosfor se u malim količinama vraća na kopno s ribom koju ulove ljudi.
Ljudski utjecaj na ciklus fosfora Prijenos fosfora s kopna u ocean značajno se povećao pod utjecajem čovjeka. Uništavanjem šuma i oranjem tla povećava se volumen toka površinske vode, a osim toga ulaze i fosforna gnojiva koja se s polja nanose u rijeke i jezera. Budući da su rezerve fosfora na kopnu ograničene, a njegov povrat iz oceana otežan, u budućnosti može doći do nedostatka fosfora u poljoprivredi, što će uzrokovati smanjenje prinosa (prvenstveno žitarica).