Energijos srautas gamtoje. Materijos ir energijos ciklas gamtoje
Bet koks gyvenimas reikalauja nuolatinio energijos ir materijos srauto. Energija eikvojama pagrindinėms gyvybės reakcijoms vykdyti, medžiaga naudojama organizmų kūnams kurti. Natūralių ekosistemų egzistavimą lydi sudėtingi medžiagų ir energijos mainų procesai tarp gyvosios ir negyvosios gamtos. Šie procesai yra labai svarbūs ir priklauso ne tik nuo biotinių bendrijų sudėties, bet ir nuo jų fizinės aplinkos.
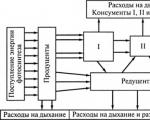
Energijos srautas bendruomenėje– tai jo perėjimas iš vieno lygio organizmų į kitą organinių junginių (maisto) cheminių ryšių pavidalu.
Medžiagos srautas (ciklas) – tai medžiagos judėjimas cheminių elementų ir jų junginių pavidalu nuo gamintojų iki skaidytojų, o po to (per chemines reakcijas, vykstančias nedalyvaujant gyviems organizmams) vėl pas gamintojus.
Medžiagos cirkuliacija ir energijos srautas nėra tapačios sąvokos, nors medžiagos judėjimui matuoti dažnai naudojami įvairūs energijos ekvivalentai (kalorijos, kilokalorijos, džauliai). Tai iš dalies paaiškinama tuo, kad visuose trofiniuose lygiuose, išskyrus pirmąjį, organizmų gyvybei reikalinga energija perduodama suvartoto maisto medžiagos pavidalu. Tiesiogiai Saulės spinduliuojamą energiją savo gyvybinei veiklai gali panaudoti tik augalai (gamintojai).
Griežtą ekosistemoje cirkuliuojančios medžiagos matavimą galima gauti atsižvelgiant į atskirų cheminių elementų, pirmiausia tų, kurie yra pagrindinė augalų ir gyvūnų ląstelių citoplazmos statybinė medžiaga, cirkuliaciją.
Skirtingai nuo medžiagų, kurios nuolat cirkuliuoja per skirtingus ekosistemos blokus ir visada gali vėl patekti į ciklą, energija organizme gali būti naudojama tik vieną kartą.
Remiantis fizikos dėsniais, energija gali keistis iš vienos formos (pvz., šviesos energija) į kitą (pavyzdžiui, potencialią maisto energiją), tačiau ji niekada nebekuriama ar sunaikinama. Negali būti vieno proceso, susieto su energijos transformacija, neprarandant jos dalies. Jo virsmų metu tam tikras energijos kiekis išsklaido šilumos pavidalu ir dėl to prarandamas. Dėl šios priežasties negali būti, pavyzdžiui, maisto medžiagų transformacijos į organizmo kūną sudarančias medžiagas, kurios vyksta šimtu procentų efektyvumu.
Visų ekosistemų egzistavimas priklauso nuo nuolatinio energijos srauto, kuris būtinas visiems organizmams gyvybinėms funkcijoms palaikyti ir savaiminiam dauginimuisi.
Tik apie pusę saulės srauto, patenkančio ant žaliųjų augalų, sugeria fotosintezės elementai, o tik maža dalis sugertos energijos (nuo 1/100 iki 1/20 dalies) sukaupiama veiklai būtinos energijos pavidalu. augalų audinių.
Tolstant nuo pirminio gamintojo, energijos srauto greitis (tai yra energijos kiekis, išreikštas energijos vienetais, perkeliamas iš vieno trofinio lygio į kitą) smarkiai sumažėja.
Energijos kiekio kritimas pereinant iš vieno trofinio lygio į aukštesnį nulemia pačių šių lygių skaičių. Apskaičiuota, kad bet kuris trofinis lygis gauna tik apie 10% (arba šiek tiek daugiau) ankstesnio lygio energijos. Todėl bendras trofinių lygių skaičius retai viršija 3–4.
Gyvosios medžiagos santykis skirtinguose trofiniuose lygiuose paprastai paklūsta tai pačiai taisyklei kaip ir gaunamos energijos santykis: kuo aukštesnis lygis, tuo mažesnė bendra biomasė ir ją sudarančių organizmų skaičius.
Įvairių organizmų grupių skaičiaus santykis leidžia suprasti bendruomenės stabilumą, nes biomasė ir kai kurių populiacijų skaičius kartu yra šios ir kitų rūšių organizmų gyvenamosios erdvės rodiklis. . Pavyzdžiui, medžių skaičius miške lemia ne tik bendrą juose esančios biomasės ir energijos pasiūlą, bet ir mikroklimatą, taip pat prieglaudų skaičių daugeliui vabzdžių ir paukščių.
Skaičių piramidės gali būti apverstos. Taip atsitinka, kai grobio populiacijos dauginimosi greitis yra didelis, ir net esant mažai biomasei tokia populiacija gali būti pakankamas maisto šaltinis plėšrūnams, kurių biomasė yra didesnė, bet reprodukcijos greitis mažas. Pavyzdžiui, ant vieno medžio (apverstos populiacijos piramidės) gali gyventi ir maitintis daug vabzdžių. Apversta biomasės piramidė būdinga vandens ekosistemoms, kur pirminiai gamintojai (fitoplanktoniniai dumbliai) labai greitai dalijasi ir dauginasi, o jų vartotojai (zooplanktoniniai vėžiagyviai) yra daug didesni, tačiau turi ilgą dauginimosi ciklą.
Ganyklų ir nuolaužų grandinės
Energija gali tekėti per bendruomenę įvairiais būdais. Jis atstovauja visų vartotojų maisto grandinei(vartotojų sistema) pridedant dar dvi nuorodas: tai negyvos organinės medžiagos Ir skaidančioji maisto grandinė(redukcijos sistema).
Energijos srautas iš augalų per žolėdžius(jie vadinami ganymu), vadinama pievų mitybos grandine.
Jų vartojamų organizmų liekanos, kurių vartotojai nenaudoja, papildo negyvas organines medžiagas. Jį sudaro išmatos, kuriose yra dalis nesuvirškinto maisto, taip pat gyvūnų lavonai, augmenijos likučiai (lapai, šakos, dumbliai) ir vadinami detritas.
Energijos srautas, atsirandantis iš negyvų organinių medžiagų ir einantis per irimo sistemą, vadinamas detrito maisto grandine.
Be panašumų, yra didelių skirtumų tarp ganyklų ir detrito maisto grandinių veikimo. Tai susideda iš to, kad in vartotojų sistemoje prarandamos išmatos ir negyvi organizmai, o redukcinėje sistemoje– Nr.
Anksčiau ar vėliau energija, esanti negyvoje organinėje medžiagoje, bus visiškai panaudota skaidytojų ir išsklaidoma kaip šiluma kvėpuojant, net jei tam reikės kelis kartus pereiti per skaidytojų sistemą. Išimtis yra tik tie atvejai, kai vietinės abiotinės sąlygos yra labai nepalankios irimo procesui (didelė drėgmė, amžinasis įšalas). Tokiais atvejais kaupiasi nepilnai apdorotų, daug energijos sunaudojančių medžiagų nuosėdos, kurios laikui bėgant ir tinkamomis sąlygomis virsta degiomis organinėmis fosilijomis – nafta, anglimis, durpėmis.
Medžiagų ciklas ekosistemoje
Natūralių ekosistemų vientisumas ypač aiškiai išryškėja turint omenyje jose cirkuliuojančių medžiagų srautus. Medžiaga gali būti perduodama uždarais ciklais (cirkuliacija), pakartotinai cirkuliuojant tarp organizmų ir aplinkos.
Cheminių elementų (tai yra medžiagų) žiediniai judėjimai (per žemę, orą, vandenį) vadinami biogeocheminiais ciklais arba cirkuliacijomis.
Sąlygiškai vadinami gyvybei būtini elementai ir ištirpusios druskos maistinių medžiagų(suteikia gyvybę) arba maistinių medžiagų. Tarp biogeninių elementų išskiriamos dvi grupės: makrotrofinės ir mikrotrofinės medžiagos.
Makrotrofinės medžiagos apima elementus, kurie sudaro gyvų organizmų audinių cheminį pagrindą. Tai apima: anglis, vandenilis, deguonis, azotas, fosforas, kalis, kalcis, magnis, siera.
Mikrotrofinės medžiagos apima elementus ir jų junginius, taip pat labai svarbius gyvoms sistemoms egzistuoti, tačiau labai mažais kiekiais. Tokios medžiagos dažnai vadinamos mikroelementai. Tai geležis, manganas, varis, cinkas, boras, natris, molibdenas, chloras, vanadis ir kobaltas. Nors mikrotrofiniai elementai yra būtini organizmams labai mažais kiekiais, jų trūkumas gali labai apriboti produktyvumą.
Maistinių medžiagų cirkuliaciją dažniausiai lydi jų cheminiai virsmai. Pavyzdžiui, nitratinis azotas gali būti paverstas baltyminiu azotu, tada paverčiamas karbamidu, paverčiamas amoniaku ir vėl susintetinamas į nitratą, veikiant mikroorganizmams. Denitrifikacijos ir azoto fiksavimo procesuose dalyvauja įvairūs biologiniai ir cheminiai mechanizmai.
Maistinių medžiagų atsargos yra įvairios. Kai kurių iš jų kaip gyvos biomasės sekvestravimo procesas sumažina abiotinėje aplinkoje likusį kiekį. O jei augalai ir kiti organizmai ilgainiui nesuirtų, išeikvotų maisto medžiagų atsargos ir gyvybė Žemėje nutrūktų. Iš to galime daryti išvadą heterotrofų, pirmiausia organizmų, veikiančių detritalinėse grandinėse, aktyvumas,– lemiamas veiksnys palaikant maistinių medžiagų ciklą ir gaminant produktus.
Pažvelkime į kai kuriuos skaitinius duomenis, nurodančius medžiagos perdavimo mastą, pereinant prie biogeocheminio anglies ciklo. Natūralus anglies šaltinis, kurį augalai naudoja organinių medžiagų sintezei, yra anglies dioksidas, kuris yra atmosferos dalis arba yra ištirpęs vandenyje. Fotosintezės metu anglies dioksidas (anglies dioksidas) paverčiamas organine medžiaga, kuri yra gyvūnų maistas. Kuro kvėpavimas, fermentacija ir deginimas grąžina anglies dioksidą į atmosferą.
Anglies atsargos mūsų planetos atmosferoje vertinamos 700 milijardų tonų, hidrosferoje – 50 000 milijardų tonų Kasmetiniais skaičiavimais, dėl fotosintezės augalų masės padidėjimas sausumoje ir vandenyje siekia 30 milijardų tonų ir Atitinkamai 150 milijardų tonų Anglies ciklas tęsiasi apie 300 metų.
Kitas pavyzdys yra fosforo ciklas. Pagrindinėse fosforo atsargose yra įvairių uolienų, kurios palaipsniui (dėl sunaikinimo ir erozijos) savo fosfatus išskiria į sausumos ekosistemas. Fosfatus suvartoja augalai ir jie juos naudoja organinėms medžiagoms sintetinti. Kai gyvūnų skerdenas skaido mikroorganizmai, fosfatai grąžinami į dirvą, o vėliau juos vėl panaudoja augalai. Be to, dalis fosfatų vandens telkiniais patenka į jūrą. Tai užtikrina fitoplanktono ir visų nuo jo priklausančių mitybos grandinių vystymąsi. Dalis jūros vandenyje esančio fosforo gali grįžti į žemę guano pavidalu.
ekosistemos biosferos energijos ciklas
Įvadas
Ekosistemų samprata ir struktūra
1 Ekosistemų samprata
2 Ekosistemų klasifikacija
3 Makroekosistemų zonavimas
4 Ekosistemos struktūra
Ekosistemų vientisumą užtikrinantys veiksniai
1 Medžiagų ciklai
2 Energijos srautas ekosistemose
3 Dinamiški procesai, užtikrinantys ekosistemų vientisumą ir tvarumą
4 Biosfera kaip pasaulinė ekosistema, užtikrinanti ekosistemų vientisumą ir tvarumą
Išvada
Įvadas
Ekosistema yra bet koks organizmų ir neorganinių komponentų rinkinys, kuriame gali vykti medžiagų cirkuliacija. Pasak N.F. Reimers (1990), ekosistema – tai bet kuri gyvų būtybių bendruomenė ir jos buveinė, sujungta į vieną funkcinę visumą, atsirandanti dėl tarpusavio priklausomybės ir priežasties-pasekmės ryšių, egzistuojančių tarp atskirų aplinkos komponentų. A. Tansley (1935) pasiūlė tokį santykį:
Yra mikroekosistemos, mezoekosistemos, makroekosistemos ir pasaulinė biosfera. Didelės sausumos ekosistemos vadinamos biomais. Ekosistemos nėra išsibarsčiusios netvarkoje, priešingai, jos yra sugrupuotos į gana taisyklingas zonas tiek horizontaliai (platumos), tiek vertikaliai (aukštyje).
Nuo pusiaujo iki ašigalių matoma tam tikra simetrija skirtingų pusrutulių biomų pasiskirstyme: atogrąžų miškuose, dykumose, stepėse, vidutinio klimato miškuose, spygliuočių miškuose, taigoje.
Kiekvieną ekosistemą sudaro du pagrindiniai komponentai: organizmai ir veiksniai iš jų negyvosios aplinkos. Organizmų (augalų, gyvūnų, mikrobų) visuma vadinama ekosistemos biota. Trofinės struktūros požiūriu (iš graikų kalbos trofe - mityba), ekosistemą galima suskirstyti į dvi pakopas: viršutinę, apatinę.
Kaip ekosistemos dalis išskiriami šie komponentai: neorganiniai, organiniai junginiai, oro, vandens ir substrato aplinka, gamintojai, autotrofiniai organizmai, vartotojai arba fagotrofai, skaidytojai ir detritivorės.
Saulės energija Žemėje sukelia du medžiagų ciklus: didelius, arba geologinius, ir mažus, biologinius (biotinius).
Ekosistemoje vyksta medžiagų ciklai. Labiausiai tiriami: anglies, deguonies, azoto, fosforo, sieros ir kt.
Nuo nuolatinio saulės energijos antplūdžio priklauso organizmų gyvybinės veiklos palaikymas ir medžiagų cirkuliacija ekosistemose, t.y. ekosistemų egzistavimas.
Ekosistemose nuolat kinta jų narių būklė ir gyvybinė veikla bei populiacijų santykis. Įvairūs pokyčiai, vykstantys bet kurioje bendruomenėje, skirstomi į du pagrindinius tipus: ciklinius ir progresyvius.
Pasaulinė ekosistema – tai biosfera, kuriai būdingas ekosistemų vientisumas ir stabilumas.
1. Ekosistemų samprata ir struktūra
1 Ekosistemų samprata
Gyvi organizmai ir jų negyva (abiotinė) aplinka yra neatskiriamai susiję vienas su kitu ir nuolat sąveikauja. Bet koks ekosistemos vienetas (biosistema) yra ekologinė sistema. Ekologinė sistema arba ekosistema yra pagrindinis funkcinis ekologijos vienetas, nes ji apima organizmus ir negyvąją aplinką – komponentus, kurie abipusiai įtakoja vienas kito savybes ir būtinas sąlygas gyvybei palaikyti tokia forma, kokia egzistuoja Žemėje. Pirmą kartą terminą „ekosistema“ 1935 metais pasiūlė anglų ekologas A. Tansley. Šiuo metu plačiai naudojamas toks ekosistemos apibrėžimas. Ekosistema yra bet koks organizmų ir neorganinių komponentų rinkinys, kuriame gali vykti medžiagų cirkuliacija. Pasak N.F. Reimers (1990), ekosistema – tai bet kuri gyvų būtybių bendruomenė ir jos buveinė, sujungta į vieną funkcinę visumą, atsirandanti dėl tarpusavio priklausomybės ir priežasties-pasekmės ryšių, egzistuojančių tarp atskirų aplinkos komponentų. Pabrėžtina, kad specifinės fizikinės ir cheminės aplinkos (biotopo) derinys su gyvų organizmų bendrija (biocenozė) formuoja ekosistemą. A. Tansley (1935) pasiūlė tokį santykį:
Ekosistema = biotopas + biocenozė
Pagal apibrėžimą V. N. Sukačiovo, biogeocenozė yra „vienarūšių gamtos reiškinių (atmosferos, uolienos, dirvožemio ir hidrologinių sąlygų) visuma tam tikrame žemės paviršiaus plote, kuri turi savo ypatingą šių jį sudarančių komponentų sąveikos specifiką ir tam tikro tipo materijos ir energijos mainai tarp savęs ir kitų gamtos reiškinių, atstovaujantys viduje prieštaringą dialektinę vienybę, nuolat judant ir vystantis.
Be ekosistemos A. Tensley ir biogeocenozės sąvokų V.N. Sukačiovas suformulavo F. Evanso (1956 m.) taisyklę, kuris pasiūlė vartoti terminą „ekosistema“ visiškai „be matmenų“ bet kokiai viršorganinei gyvai sistemai, sąveikaujančiai su aplinka. Tačiau daugelis autorių terminui „ekosistema“ suteikė biogeocenozės reikšmę, t.y. elementari ekosistema, o kartu ir aukščiau viršbiogeocenotinių darinių hierarchijoje iki biosferos ekosistemos.
2 Ekosistemų klasifikacija
Žemėje egzistuojančios ekosistemos yra įvairios. Yra mikroekosistemos (pavyzdžiui, pūvančio medžio kamienas), mezoekosistemos (miškas, tvenkinys ir kt.), makroekosistemos (žemynas, vandenynas ir kt.) ir globali biosfera.
Didelės sausumos ekosistemos vadinamos biomais. Kiekviename biome yra keletas mažesnių, tarpusavyje susijusių ekosistemų. Yra keletas ekosistemų klasifikacijų:
Visžalis atogrąžų miškas
Dykuma: žolė ir krūmas Chaparral - vietovės su lietingomis žiemomis ir sausomis vasaromis
Atogrąžų Graslencas ir Savana
Vidutinio klimato stepė
Vidutinio klimato lapuočių miškas
Borealiniai spygliuočių miškai
Tundra: arktinė ir alpinė
Gėlo vandens ekosistemų tipai:
Juostelė (nejudantys vandenys): ežerai, tvenkiniai ir kt.
Lotika (tekantys vandenys): upės, upeliai ir kt.
Pelkės: pelkės ir pelkėti miškai
Jūrų ekosistemų tipai
Atviras vandenynas (pelaginė)
Kontinentinio šelfo vandenys (pakrančių vandenys)
Augančios vietovės (derlingos vietovės su produktyvia žvejyba)
Estuarijos (pakrančių įlankos, sąsiauriai, upių žiotys, druskingos pelkės ir kt.). Sausumos biomai čia išsiskiria natūraliomis arba originaliomis augmenijos ypatybėmis, o vandens ekosistemų tipai – pagal geologinius ir fizinius ypatumus. Išvardyti pagrindiniai ekosistemų tipai reprezentuoja aplinką, kurioje vystėsi žmonių civilizacija, ir atstovauja pagrindinėms biotinėms bendruomenėms, palaikančioms gyvybę Žemėje.
3 Makroekosistemų zonavimas
Geografinio ekosistemų pasiskirstymo tyrimo galima imtis tik didelių ekologinių vienetų – makroekosistemų, kurios vertinamos žemyno mastu, lygiu. Ekosistemos nėra išsibarsčiusios netvarkoje, priešingai, jos yra sugrupuotos į gana taisyklingas zonas tiek horizontaliai (platumos), tiek vertikaliai (aukštyje). Tai patvirtina periodinis A.A geografinės zonos dėsnis. Grigorjeva - M.I. Budyko: keičiantis fizinėms-geografinėms Žemės zonoms, panašios kraštovaizdžio zonos ir kai kurios jų bendros savybės periodiškai kartojasi. Apie tai taip pat buvo kalbama svarstant gyvenimo aplinką žemė-oras. Įstatymo nustatytas periodiškumas pasireiškia tuo, kad sausumo indekso reikšmės įvairiose zonose svyruoja nuo 0 iki 4-5, tris kartus tarp ašigalių ir pusiaujo jie yra artimi vienybei. Šios vertės atitinka didžiausią biologinį kraštovaizdžio produktyvumą.
Periodiškas savybių pasikartojimas vieno hierarchinio lygio sistemų serijoje tikriausiai yra bendras visatos dėsnis, suformuluotas kaip periodiškumo dėsnis sisteminių agregatų struktūroje, arba sistemos periodinis dėsnis – specifinės vieno lygio natūralios sistemos ( Organizacijos polygis) sudaro periodinę arba pasikartojančią morfologiškai panašių struktūrų seriją viršutinėse ir apatinėse sistemos erdvės ir laiko ribose, už kurias sistemų egzistavimas tam tikrame lygyje tampa neįmanomas. Jie pereina į nestabilią būseną arba virsta kita sistemos struktūra, įskaitant kitą organizavimo lygį.
Temperatūra ir krituliai lemia pagrindinių sausumos biomų vietą žemės paviršiuje. Temperatūros ir kritulių tendencija tam tikroje vietovėje per gana ilgą laiką yra tai, ką mes vadiname klimatu. Klimatas skirtinguose pasaulio regionuose yra skirtingas. Metinis kritulių kiekis svyruoja nuo 0 iki 2500 mm ir daugiau. Temperatūros ir kritulių režimai derinami labai skirtingai.
Klimato sąlygų specifika savo ruožtu lemia konkretaus biomo vystymąsi.
Nuo pusiaujo iki ašigalių matoma tam tikra simetrija skirtingų pusrutulių biomų pasiskirstyme:
Atogrąžų miškai (šiaurinė Pietų Amerika, Centrinė Amerika, Pusiaujo Afrikos vakarinės ir centrinės dalys, Pietryčių Azija, šiaurės vakarų Australijos pakrantės zonos, Indijos ir Ramiojo vandenyno salos). Klimatas be sezonų kaitos (artumas iki pusiaujo), vidutinė metinė temperatūra viršija 17°C (dažniausiai 28°C), vidutinis metinis kritulių kiekis viršija 2400 mm.
Augalija: dominuoja miškai. Kai kurių rūšių medžiai yra iki 60 m aukščio, ant jų kamienų ir šakų auga epifitiniai augalai, kurių šaknys nepasiekia dirvos, sumedėję vynmedžiai, kurie įsišaknija dirvoje ir laipioja medžiais į viršūnes. Visa tai sudaro storą baldakimą.
Fauna: rūšių sudėtis turtingesnė nei visų kitų biomų kartu paėmus. Ypač daug yra varliagyvių, roplių ir paukščių (varlių, driežų, gyvačių, papūgų), beždžionių ir kitų smulkių žinduolių, egzotiškų ryškiaspalvių vabzdžių ir ryškiaspalvių žuvų rezervuaruose.
Savybės: dirvožemis dažnai yra plonas ir skurdus, daugiausia maistinių medžiagų yra įsišaknijusios augalijos paviršinėje biomasėje.
2. Savanos (subekvatorinė Afrika, Pietų Amerika, nemaža Pietų Indijos dalis). Klimatas didžiąją metų dalį yra sausas ir karštas. Gausūs krituliai drėgnuoju metų laiku. Temperatūra: vidutinė metinė-aukšta. Kritulių iškrenta 750-1650 mm/metus, daugiausia lietinguoju sezonu. Augalija – melsvieji (javai) augalai su retais lapuočiais medžiais. Fauna: stambūs žolėdžiai žinduoliai, tokie kaip antilopės, zebrai, žirafos, raganosiai, plėšrūnai – liūtai, leopardai, gepardai.
Dykumos (kai kurios Afrikos sritys, pavyzdžiui, Sachara; Viduriniai Rytai ir Centrinė Azija, Didysis baseinas ir JAV pietvakariai bei šiaurinė Meksika ir kt.). Klimatas labai sausas. Temperatūra – karštos dienos ir šaltos naktys. Kritulių yra mažiau nei 250 mm per metus. Augalija: reti krūmai, dažnai dygliuoti, kartais kaktusai ir žemos žolės, po retų liūčių greitai padengianti žemę žydinčiu kilimu. Augalai turi plačias paviršinių šaknų sistemas, kurios sulaiko drėgmę nuo retų kritulių, taip pat šaknis, kurios prasiskverbia į žemę iki gruntinio vandens lygio (30 m ir giliau). Fauna: įvairūs graužikai (kengūros žiurkės ir kt.), rupūžės, driežai, gyvatės ir kiti ropliai, pelėdos, ereliai, grifai, dideli paukščiai ir vabzdžiai.
4. Stepės (Šiaurės Amerikos centras, Rusija, tam tikros Afrikos ir Australijos sritys, į pietryčius nuo Pietų Amerikos). Klimatas sezoninis. Temperatūra – vasaros temperatūra svyruoja nuo vidutiniškai šiltos iki karštos, žiemos temperatūra žemesnė nei 0°C. Kritulių - 750-2000 mm/metus. Augalija: kai kuriose Šiaurės Amerikos prerijose vyrauja iki 2 m ir aukštesnės melsvažolės (javai) arba iki 50 cm, pavyzdžiui, Rusijos stepėse, drėgnose vietose pavieniai medžiai ir krūmai. Fauna: stambūs žolėdžiai žinduoliai – stumbrai, smaragiškiai antilopės (Šiaurės Amerika), laukiniai arkliai (Eurazija), kengūros (Australija), žirafos, zebrai, baltieji raganosiai, antilopės (Afrika); Plėšrūnai yra kojotai, liūtai, leopardai, gepardai, hienos, įvairūs paukščiai ir smulkūs žinduoliai, tokie kaip triušiai, žemės voverės ir aardvarkai.
5. Vidutinio klimato miškai (Vakarų Europa, Rytų Azija, rytų JAV). Klimatas - sezoninis, kai žiemos temperatūra žemesnė nei 0 0C. Krituliai - 750-2000 mm/metus. Augalija: vyrauja iki 35-45 m aukščio plačialapių lapuočių medžių miškai (ąžuolai, hikoriai, klevai), krūminis pomiškis, samanos, kerpės. Fauna: žinduoliai (baltauodegiai elniai, kiaulytės, meškėnai, oposumai, voverės, triušiai, vėgėlės), paukščiai (karselės, snapeliai, juodvarniai, pelėdos, sakalai), gyvatės, varlės, salamandros, žuvys (upėtakis, ešeriai, šamai ir kt.) . ), gausi dirvožemio mikrofauna (12.3 pav.). Biota prisitaikiusi prie sezoninio klimato: žiemojimo, migracijos, ramybės žiemos mėnesiais. 6. Spygliuočių miškai, taiga (šiauriniai Šiaurės Amerikos, Europos ir Azijos regionai). Klimatas ilgos ir šaltos žiemos, kai kurie krituliai iškrenta sniego pavidalu. Augalija: vyrauja visžaliai spygliuočių miškai, daugiausia eglės, pušys, kėniai. Fauna: stambūs žolėdžiai kanopiniai gyvūnai (elniai, šiaurės elniai), smulkūs žolėdžiai žinduoliai (kiškis, voverė, graužikai), vilkas, lūšis, lapė, juodasis lokys, grizlis, kurtiniai, audinės ir kiti plėšrūnai, daugybė kraują siurbiančių vabzdžių trumpą vasarą laikas. Klimatas labai šaltas su poliarinėmis dienomis ir poliarinėmis naktimis. Vidutinė metinė temperatūra žemesnė nei -5°C. Per kelias trumpos vasaros savaites žemė atšyla ne daugiau kaip vieno metro gylyje. Kritulių yra mažiau nei 250 mm per metus. Augalija: vyrauja lėtai augančios kerpės, samanos, žolės ir viksvos, žemaūgiai krūmai. Fauna: stambūs žolėdžiai kanopiniai gyvūnai (šiaurės elniai, muskuso jautis), smulkūs rausvantys žinduoliai (pavyzdžiui, lemingai ištisus metus), plėšrūnai, kurie žiemą įgauna maskuojančią baltą spalvą (arktinė lapė, lūšis, erminas, snieginė pelėda). Trumpą vasarą tundroje peri daug migruojančių paukščių, tarp jų ypač daug vandens paukščių, kurie minta čia gausiai aptinkamais vabzdžiais ir gėlavandeniais bestuburiais. Taip pat labai ryškus žemės ekosistemų vertikalus aukščio zonavimas, ypač ryškaus reljefo vietose. Drėgmė yra pagrindinis veiksnys, lemiantis biomo tipą. Esant pakankamai dideliam kritulių kiekiui, vystosi miško augalija. Temperatūra lemia miško tipą. Lygiai tokia pati situacija yra stepių ir dykumų biomuose. Augalijos tipų pokyčiai šaltuose regionuose vyksta esant mažesniam metiniam kritulių kiekiui, nes esant žemai temperatūrai, mažiau vandens prarandama garuojant. Temperatūra tampa pagrindiniu veiksniu tik labai šaltomis sąlygomis, kai yra amžinas įšalas. Taigi ekosistemų sudėtis labai priklauso nuo jų funkcinės „paskirties“ ir atvirkščiai. Pasak N.F. Reimers (1994), tai atsispindi ekologinio komplementarumo (komplementarumo) principu: jokia funkcinė ekosistemos dalis (ekologinis komponentas, elementas ir kt.) negali egzistuoti be kitų funkciškai vienas kitą papildančių dalių. Jai artimas ir jį plečiantis yra ekologinio sutapimo (atitikties) principas:. funkciškai papildydami vienas kitą, gyvieji ekosistemos komponentai tam išvysto atitinkamas adaptacijas, suderintas su abiotinės aplinkos sąlygomis, kurią iš esmės transformuoja tie patys organizmai (bioklimatas ir kt.), t.y. egzistuoja dviguba atitikmenų serija – tarp organizmų ir jų buveinės – išorinės ir sukurtos cenozės. 4 Ekosistemos struktūra Trofinės struktūros požiūriu (iš graikų kalbos trofe - mityba), ekosistemą galima suskirstyti į dvi pakopas: Viršutinė – autotrofinė (savaime maitinanti) pakopa, arba „žalia juosta“, įskaitant augalus ar jų dalis, kuriuose yra chlorofilo, kur vyrauja skaičiavimo energijos fiksavimas ir paprastų neorganinių junginių naudojimas. Apatinis – heterotrofinis (kitų maitinamas) sluoksnis arba „rudoji juosta“ iš dirvožemių ir nuosėdų, pūvančių medžiagų, šaknų ir kt., kuriame vyrauja kompleksinių junginių panaudojimas, transformacija ir skaidymas. Biologiniu požiūriu, kaip ekosistemos dalis, išskiriami šie komponentai: Ekosistemoje maisto ir energijos ryšiai tarp kategorijų visada yra nedviprasmiški ir eina tokia kryptimi: autotrofai - heterotrofai arba išsamesnė forma; autotrofai -> vartotojai -> skaidytojai (destruktoriai). Pagrindinis ekosistemų energijos šaltinis yra Saulė. Energijos srautas (pagal T.A. Akimova, V.V. Khaskin, 1994), kurį Saulė siunčia į Žemės planetą, per metus viršija 20 mln. EJ. Tik ketvirtadalis šio srauto priartėja prie atmosferos ribos. Iš jų apie 70% atsispindi, sugeria atmosferą ir išspinduliuoja ilgųjų bangų infraraudonųjų spindulių pavidalu. Saulės spinduliuotė, patenkanti į Žemės paviršių, yra 1,54 mln. EJ per metus. Šis didžiulis energijos kiekis 5000 kartų viršija visą XX amžiaus pabaigos žmonijos energiją ir 5,5 karto daugiau nei visų turimų organinės kilmės iškastinio kuro išteklių, sukauptų mažiausiai per 100 milijonų metų. Didžioji dalis saulės energijos, pasiekiančios planetos paviršių, tiesiogiai paverčiama šiluma, šildančiu vandeniu arba dirvožemiu, o tai savo ruožtu sušildo orą. Ši šiluma yra vandens ciklo, oro ir vandenyno srovių, lemiančių orus, varomoji jėga ir palaipsniui išleidžiama į kosmosą, kur ji prarandama. Norint nustatyti ekosistemų vietą šiame natūraliame energijos sraute, svarbu suvokti, kad kad ir kokios plačios ir sudėtingos jos būtų, jos naudoja tik nedidelę jos dalį. Tai suponuoja vieną iš pagrindinių ekosistemų funkcionavimo principų: jos egzistuoja dėl neteršiančios ir beveik amžinos saulės energijos, kurios kiekis yra gana pastovus ir gausus. Leiskite mums išsamiau pateikti kiekvieną iš išvardytų saulės energijos savybių: 2. Ekosistemų vientisumą užtikrinantys veiksniai 1 Medžiagų ciklai Abu ciklai yra tarpusavyje susiję ir atspindi tarsi vieną procesą. Apskaičiuota, kad visas atmosferoje esantis deguonis per organizmus (susijungia kvėpuojant ir išsiskiria fotosintezės metu) per 2000 metų, anglies dioksidas atmosferoje per 300 metų apsisuka priešinga kryptimi, o visas Žemėje esantis vanduo per 2 000 000 metų suyra ir atkurta fotosintezės ir kvėpavimo būdu. Abiotinių veiksnių ir ekosistemos gyvų organizmų sąveiką lydi nuolatinė medžiagų cirkuliacija tarp biotopo ir biocenozės kintančių organinių ir mineralinių junginių pavidalu. Cheminių elementų mainai tarp gyvų organizmų ir neorganinės aplinkos, kurių įvairios stadijos vyksta ekosistemoje, vadinamas biogeochemine cirkuliacija arba biogeocheminiu ciklu. Vandens ciklas. Reikšmingiausias ciklas Žemėje pagal perkeliamas mases ir energijos suvartojimą yra planetų hidrologinis ciklas – vandens ciklas. Kas sekundę į jį patenka 16,5 milijono m3 vandens ir tam išleidžiama daugiau nei 40 milijardų MW saulės energijos (pagal T.A. Akimova V.V. Haskin, (1994)). Tačiau šis ciklas yra ne tik vandens masių perkėlimas. Tai faziniai virsmai, tirpalų ir suspensijų susidarymas, nusodinimas, kristalizacija, fotosintezės procesai, taip pat įvairios cheminės reakcijos. Gyvenimas atsirado ir tęsiasi šioje aplinkoje. Vanduo yra pagrindinis elementas, būtinas gyvybei. Kiekybiškai tai yra labiausiai paplitęs neorganinis gyvosios medžiagos komponentas. Žmonėms vanduo sudaro 63% kūno svorio, grybuose - 80%, augaluose - 80-90%, o kai kuriose medūzose - 98%. Vanduo, kaip matysime šiek tiek vėliau, dalyvauja biologiniame cikle ir tarnauja kaip vandenilio ir deguonies šaltinis, sudarantis tik nedidelę viso jo tūrio dalį. Skystos, kietos ir garinės būsenos vandens yra visose trijose pagrindinėse biosferos dalyse: atmosferoje, hidrosferoje ir litosferoje. Visus vandenis vienija bendra „hidrosferos“ sąvoka. Hidrosferos komponentai yra tarpusavyje susiję nuolatiniais mainais ir sąveika. Vanduo, nuolat judėdamas iš vienos būsenos į kitą, daro mažus ir didelius ciklus. Vandens išgaravimas nuo vandenyno paviršiaus, vandens garų kondensacija atmosferoje ir krituliai vandenyno paviršiuje sudaro nedidelį ciklą. Kai vandens garus oro srovės nuneša į žemę, ciklas tampa daug sudėtingesnis. Šiuo atveju dalis kritulių išgaruoja ir grįžta į atmosferą, kita dalis maitina upes ir rezervuarus, bet galiausiai grįžta į vandenyną per upę ir požeminį nuotėkį, taip užbaigiant didelį ciklą. Biotinis (biologinis) ciklas. Biotinis (biologinis) ciklas reiškia medžiagų apykaitą tarp dirvožemio, augalų, gyvūnų ir mikroorganizmų. Biotinis (biologinis) ciklas – tai cheminių elementų srautas iš dirvožemio, vandens ir atmosferos į gyvus organizmus, gaunamų elementų pavertimas naujais kompleksiniais junginiais ir jų grįžimas atgal į gyvybės procesą, kasmet nukritus daliai organinių medžiagų. medžiaga arba visiškai negyvi organizmai, įtraukti į ekosistemos sudėtį. Dabar biotinį ciklą pateiksime cikline forma. Centrinį biotinį ciklą (pagal T.A. Akimovą, V.V., Khashhiną) sudarė primityvūs vienaląsčiai gamintojai (P) ir skaidytojai-destruktoriai (D). Mikroorganizmai geba greitai daugintis ir prisitaikyti prie įvairių sąlygų, pavyzdžiui, savo mityboje naudoti įvairiausius substratus – anglies šaltinius. Aukštesni organizmai tokių gebėjimų neturi. Išbaigtose ekosistemose jie gali egzistuoti mikroorganizmų pagrindu esančios struktūros pavidalu. Pirmiausia išsivysto daugialąsčiai augalai (P) – aukštesni gamintojai. Kartu su vienaląsčiais organizmais jie sukuria organines medžiagas fotosintezės procese, naudodami saulės spinduliuotės energiją. Vėliau sujungiami pirminiai vartotojai – žolėdžiai gyvūnai (T), o vėliau – mėsėdžiai vartotojai. Mes ištyrėme biotinį žemės ciklą. Tai visiškai taikoma vandens ekosistemų, pavyzdžiui, vandenyno, biotiniam ciklui. Visi organizmai biotiniame cikle užima tam tikrą vietą ir atlieka savo gaunamas energijos srauto šakų transformavimo ir biomasės perdavimo funkcijas. Vienaląsčių skaidytojų (destruktorių) sistema sujungia visus, nuasmenina jų substancijas ir uždaro bendrą ratą. Jie grąžina į abiotinę biosferos aplinką visus elementus, reikalingus naujoms ir naujoms revoliucijoms. Reikėtų pabrėžti svarbiausias biotinio ciklo ypatybes. Fotosintezė yra galingas natūralus procesas, kasmet į ciklą įtraukiantis didžiules biosferos medžiagų mases ir lemiantis didelį deguonies potencialą. Dėl anglies dioksido ir vandens sintetinama organinė medžiaga ir išsiskiria laisvasis deguonis. Tiesioginiai fotosintezės produktai yra įvairūs organiniai junginiai, ir apskritai fotosintezės procesas yra gana sudėtingas. Be fotosintezės, kurioje dalyvauja deguonis, vadinamosios deguonies fotosintezės, taip pat turėtume sutelkti dėmesį į fotosintezę be deguonies arba chemosintezę. Chemosintetiniai organizmai apima nitrifikatorius, karboksidobakterijas, sieros bakterijas, tioninės geležies bakterijas ir vandenilio bakterijas. Jie pavadinti pagal jų oksidacijos substratus, kurie gali būti NH3, NO2, CO, H2S, S, Fe2+, H2. Kai kurios rūšys yra privalomi chemolitoautotrofai, kitos yra fakultatyvinės. Pastarosios apima karboksidobakterijas ir vandenilio bakterijas. Chemosintezė būdinga giliavandenėms hidroterminėms angoms. Fotosintezė, išskyrus keletą išimčių, vyksta visame Žemės paviršiuje, sukuria didžiulį geocheminį efektą ir gali būti išreikšta visos anglies masės kiekiu, kasmet dalyvaujant visos biosferos organinės gyvosios medžiagos kūrime. Bendrasis medžiagų ciklas, susijęs su organinių medžiagų statyba fotosintezės būdu, taip pat apima cheminius elementus, tokius kaip N, P, S, taip pat metalus - K, Ca, Mg, Na, Al. Kai organizmas miršta, vyksta atvirkštinis procesas – organinių medžiagų skilimas vykstant oksidacijai, irimui ir kt. susidarant galutiniams skilimo produktams. Žemės biosferoje šis procesas lemia tai, kad gyvosios medžiagos biomasės kiekis yra šiek tiek pastovus. Ekosferos biomasė (2 10|2t) yra septyniomis eilėmis mažesnė už žemės plutos masę (2 .10|9t). Žemės augalai kasmet gamina 1,6,10"% arba 8% ekosferos biomasės organinės medžiagos. Destruktoriai, kurie sudaro mažiau nei 1% visos planetos organizmų biomasės, apdoroja organinių medžiagų masę, kuri yra 10 kartų didesnis nei jų pačių biomasė.Vidutiniškai biomasės atsinaujinimo laikotarpis yra 12,5 metų. Tarkime, kad gyvosios medžiagos masė ir biosferos produktyvumas nuo kambro iki dabar buvo vienodi (530 mln. metų), tada bendras organinių medžiagų, praėjusių pasaulinį biotinį ciklą ir sunaudotos gyvybei planetoje, kiekis bus 2,10 "2-5,ZL08/12,5=8,5L0|9t, o tai 4 kartus viršija žemės plutos masę. Dėl šių skaičiavimų N. S. Pechurkin (1988) rašė: „Galime sakyti, kad atomai, sudarantys mūsų kūną, buvo senovės bakterijose, dinozauruose ir mamutuose. Biogeninės atomų migracijos dėsnis V.I. Vernadskis teigia: „Cheminių elementų migracija žemės paviršiuje ir visoje biosferoje vyksta arba tiesiogiai dalyvaujant gyvajai medžiagai (biogeninė migracija), arba vyksta aplinkoje, kurios geocheminės savybės (O2, CO2, H2, ir tt) yra determinuota gyvoji medžiaga, tiek ta, kuri šiuo metu gyvena biosferoje, tiek ta, kuri veikė Žemę per visą geologinę istoriją. Į IR. Vernadskis 1928-1930 m giliuose apibendrinimuose apie procesus biosferoje jis pateikė idėją apie penkias pagrindines gyvosios medžiagos biogeochemines funkcijas. Pirmoji funkcija yra dujos. Antroji funkcija yra koncentracija. Trečioji funkcija yra redoksas. Ketvirtoji funkcija yra biocheminė. Penktoji funkcija – biogeocheminė žmonijos veikla, apimanti vis didesnį žemės plutoje esančios medžiagos kiekį pramonės, transporto ir žemės ūkio reikmėms. Biologinis ciklas kinta įvairiose gamtinėse zonose ir klasifikuojamas pagal rodiklių rinkinį: augalų biomasė, kraikas, kraikas, biomasėje fiksuotų elementų kiekis ir kt. Bendra biomasė didžiausia miško zonoje, o požeminių organų dalis miškuose yra mažiausia. Tai patvirtina ir biologinio ciklo intensyvumo indeksas – vado masės ir ją sudarančios vados dalies santykis. Anglies ciklas. Iš visų biogeocheminių ciklų anglies ciklas, be jokios abejonės, yra intensyviausias. Anglis dideliu greičiu cirkuliuoja tarp įvairių neorganinių terpių ir per maisto tinklus gyvų organizmų bendruomenėse. CO ir CO2 atlieka tam tikrą vaidmenį anglies cikle.Dažnai Žemės biosferoje anglis yra judriausia CO2 forma. Pirminio anglies dioksido šaltinis biosferoje yra vulkaninis aktyvumas, susijęs su pasaulietiniu mantijos degazavimu ir žemutiniais žemės plutos horizontais. CO2 migracija biosferoje vyksta dviem būdais. Pirmasis būdas yra jį absorbuoti fotosintezės metu, kai susidaro gliukozė ir kitos organinės medžiagos, iš kurių yra sukurti visi augalų audiniai. Vėliau jie pernešami maisto grandinėmis ir sudaro visų kitų gyvų būtybių audinius ekosistemoje. Ant paviršiaus mirštant augalams ir gyvūnams, susidaro organinių medžiagų oksidacija, susidarant CO2. Anglies atomai taip pat grąžinami į atmosferą deginant organines medžiagas. Svarbi ir įdomi anglies ciklo ypatybė yra ta, kad tolimose geologinėse epochose, prieš šimtus milijonų metų, nemaža dalis fotosintezės procesuose susidariusios organinės medžiagos nebuvo panaudota nei vartotojų, nei skaidytojų, o kaupėsi litosferoje. iškastinio kuro pavidalu: nafta, anglis, skalūnai, durpės ir kt. Šis iškastinis kuras išgaunamas didžiuliais kiekiais, kad būtų patenkinti mūsų pramonės visuomenės energijos poreikiai. Degindami jį tam tikra prasme užbaigiame anglies ciklą. Antruoju būdu anglies migracija vykdoma sukuriant karbonatų sistemą įvairiuose rezervuaruose, kur CO2 virsta H2CO3, HCO, CO3. Vandenyje ištirpusio kalcio (arba magnio) pagalba biogeniniais ir abiogeniniais keliais nusodinami karbonatai (CaCO3). Susidaro stori kalkakmenio sluoksniai. Pasak A.B. Ronov, fotosintezės produktuose palaidotos anglies ir karbonatinėse uolienose esančios anglies santykis yra 1:4. Kartu su dideliu anglies ciklu žemės paviršiuje ir vandenyne taip pat yra keletas mažų anglies ciklų. Apskritai, be antropogeninės intervencijos, anglies kiekis biogeocheminiuose rezervuaruose: biosferoje (biomasė + dirvožemis ir nuolaužos), nuosėdinėse uolienose, atmosferoje ir hidrosferoje yra palaikomas aukštu pastovumo laipsniu (pagal T. A. Akimovą, V. V. Haskiną (1994)). ). Nuolatinį anglies keitimąsi, viena vertus, tarp biosferos ir, kita vertus, tarp atmosferos ir hidrosferos, lemia gyvosios medžiagos dujinė funkcija – fotosintezės, kvėpavimo ir naikinimo procesai. 6-1010 t/metus.Į atmosferą ir hidrosferą patenka anglies srautas ir vulkaninės veiklos metu vidutiniškai 4,5 106 t/metus. Bendra anglies masė iškastiniame kure (naftoje, dujose, anglyje ir kt.) yra 3,2*1015 tonų, o tai atitinka vidutinį 7 mln. tonų per metus kaupimo greitį. Šis kiekis yra nereikšmingas, palyginti su cirkuliuojančios anglies mase ir tarsi iškrito iš ciklo ir jame pasimetė. Vadinasi, ciklo atvirumo (netobulumo) laipsnis yra 10"4, arba 0,01%, ir atitinkamai uždarumo laipsnis yra 99,99%. Tai reiškia, viena vertus, kiekvienas anglies atomas dalyvavo cikle. dešimtis tūkstančių kartų, kol iškrito iš ciklo, atsidūrė gelmėse, kita vertus, organinių medžiagų sintezės ir irimo srautai biosferoje yra labai tiksliai suderinami vienas su kitu. 0,2 % mobiliosios anglies atsargų yra nuolatinėje cirkuliacijoje. Biomasės anglis atsinaujina per 12 metų, atmosferoje – per 8 metus. Deguonies ciklas. Deguonis (O2) vaidina svarbų vaidmenį daugelio mūsų planetos gyvų organizmų gyvenime. Kiekybine prasme tai yra pagrindinis gyvosios medžiagos komponentas. Pavyzdžiui, jei atsižvelgsime į audiniuose esantį vandenį, žmogaus kūne yra 62,8% deguonies ir 19,4% anglies. Apskritai biosferoje šis elementas, palyginti su anglimi ir vandeniliu, yra pagrindinis tarp paprastų medžiagų. Biosferoje vyksta greitas deguonies keitimasis su gyvais organizmais arba jų liekanomis po mirties. Augalai, kaip taisyklė, gamina laisvą deguonį, o gyvūnai jį sunaudoja kvėpuodami. Būdamas plačiausiai paplitęs ir judriausias elementas Žemėje, deguonis neriboja ekosferos egzistavimo ir funkcijų, nors deguonies prieinamumas vandens organizmams gali būti laikinai apribotas. Deguonies ciklas biosferoje yra labai sudėtingas, nes su juo reaguoja daug organinių ir neorganinių medžiagų. Dėl to atsiranda daug epiciklų, vykstančių tarp litosferos ir atmosferos arba tarp hidrosferos ir šių dviejų aplinkų. Deguonies ciklas tam tikra prasme yra panašus į atvirkštinį anglies dioksido ciklą. Vieno judėjimas vyksta priešinga kryptimi nei kitas. Atmosferos deguonis suvartojamas ir jį pakeičia pirminiai gamintojai gana greitai. Taigi, norint visiškai atnaujinti visą atmosferos deguonį, reikia 2000 metų. Šiais laikais deguonies kaupimasis atmosferoje nevyksta, o jo kiekis (20,946%) išlieka pastovus. Viršutiniuose atmosferos sluoksniuose ultravioletiniams spinduliams veikiant deguonį susidaro ozonas – O3. Ozono susidarymui išleidžiama apie 5% Žemę pasiekiančios saulės energijos – apie 8,6 * 1015 W. Reakcijos yra lengvai grįžtamos. Ozonui irstant išsiskiria ši energija, kuri palaiko aukštą temperatūrą viršutiniuose atmosferos sluoksniuose. Vidutinė ozono koncentracija atmosferoje yra apie 106 tūriai. %; maksimali O3 koncentracija - iki 4-10 "* tūrio % - pasiekiama 20-25 km aukštyje (T.A. Akimova, V.V. Haskin, 1998). Ozonas tarnauja kaip savotiškas UV filtras: jis blokuoja didelę dalį kietųjų ultravioletinių spindulių. Tikriausiai ozono sluoksnio susidarymas buvo viena iš sąlygų gyvybei išplaukti iš vandenyno ir kolonizuoti žemę. Didžioji dalis per geologines epochas susidariusio deguonies neliko atmosferoje, o buvo fiksuojama litosferos karbonatų, sulfatų, geležies oksidų ir kt. Ši masė yra 590 * 1014 tonų, palyginti su 39 * 1014 tonų deguonies, kuris cirkuliuoja biosferoje dujų arba sulfatų pavidalu, ištirpusių žemyniniuose ir vandenynų vandenyse. Azoto ciklas. Azotas yra būtinas biogeninis elementas, nes jis yra baltymų ir nukleorūgščių dalis. Azoto ciklas yra vienas sudėtingiausių, nes apima ir dujų, ir mineralų fazes, o kartu ir pačius idealiausius ciklus. Azoto ciklas yra glaudžiai susijęs su anglies ciklu. Paprastai azotas seka anglį, kartu su ja dalyvauja formuojant visas baltymines medžiagas. Atmosferos oras, kuriame yra 78% azoto, yra neišsenkantis rezervuaras. Tačiau dauguma gyvų organizmų negali tiesiogiai panaudoti šio azoto. Kad augalai pasisavintų azotą, jis turi būti amonio (NH*) arba nitrato (NO3) jonų dalis. Azoto dujos nuolat patenka į atmosferą dėl denitrifikuojančių bakterijų darbo, o fiksuojančios bakterijos kartu su melsvadumbliais (cianofitais) nuolat jį sugeria, paverčiant nitratais. Azoto ciklas aiškiai matomas naikintojų lygyje. Baltymus ir kitas organinio azoto formas, esančias augaluose ir gyvūnuose po jų mirties, veikia heterotrofinės bakterijos, aktinomicetai, grybai (bioredukuojantys mikroorganizmai), kurie gamina jiems reikalingą energiją redukuodami šį organinį azotą, paversdami jį amoniaku. Dirvožemyje vyksta nitrifikacijos procesas, susidedantis iš reakcijų grandinės, kur, dalyvaujant mikroorganizmams, vyksta amonio jonų (NH4+) oksidacija į nitritus (NO~) arba nitritai į nitratus (N0~). Denitrifikacijos proceso esmė yra nitritų ir nitratų redukcija į dujinius junginius – molekulinį azotą (N2) arba azoto oksidą (N20). Neorganinis nitratų susidarymas nedideliais kiekiais nuolat vyksta atmosferoje, perkūnijos metu elektros iškrovų metu atmosferos azotą surišant su deguonimi, o vėliau su lietumi iškritus ant dirvožemio paviršiaus. Kitas atmosferos azoto šaltinis yra ugnikalniai, kurie kompensuoja iš ciklo pašalinto azoto praradimą nusėdimo arba nusėdimo į vandenynų dugną metu. Apskritai vidutinis abiotinės kilmės nitratinio azoto pasiūla nusėdant iš atmosferos į dirvą neviršija 10 kg (metai/ha), laisvosios bakterijos gamina 25 kg (metai/ha), o Rhizobium simbiozė su ankštiniais augalais. užaugina vidutiniškai 200 kg (metai/ha). Didžiąją fiksuoto azoto dalį denitrifikuojančios bakterijos perdirba į N2 ir grąžina į atmosferą. Tik apie 10% amonifikuoto ir nitrifikuoto azoto iš dirvožemio pasisavina aukštesni augalai ir patenka į daugialąsčių biocenozių atstovų žinią. Fosforo ciklas. Fosforo ciklas biosferoje yra susijęs su medžiagų apykaitos procesais augaluose ir gyvūnuose. Šis svarbus ir reikalingas protoplazmos elementas, kurio sausumos augaluose ir dumbliuose yra 0,01–0,1%, gyvūnuose nuo 0,1% iki kelių procentų, cirkuliuoja, palaipsniui iš organinių junginių virsdamas fosfatais, kuriuos vėl gali naudoti augalai. Tačiau fosforas, skirtingai nei kiti biofiliniai elementai, migracijos metu nesudaro dujų formos. Fosforo rezervuaras yra ne atmosfera, kaip azotas, o mineralinė litosferos dalis. Pagrindiniai neorganinio fosforo šaltiniai yra magminės uolienos (apatitai) arba nuosėdinės uolienos (fosforitai). Iš uolienų neorganinis fosforas dalyvauja apyvartoje, išplaudamas ir ištirpdamas žemyniniuose vandenyse. Patekęs į sausumos ekosistemas ir dirvožemį, fosforą augalai pasisavina iš vandeninio tirpalo neorganinių fosfatų jonų pavidalu ir yra įtrauktas į įvairius organinius junginius, kur atsiranda organinio fosfato pavidalu. Fosforas maisto grandinėmis juda iš augalų į kitus ekosistemos organizmus. Fosforas į vandens ekosistemas pernešamas tekančiomis vandenimis. Upės nuolat praturtina vandenynus fosfatais. Kur fosforas tampa fitoplanktono dalimi. Kai kurie fosforo junginiai migruoja sekliuose gyliuose, juos suvartoja organizmai, o kita dalis prarandama didesniame gylyje. Negyvos organizmų liekanos lemia fosforo kaupimąsi skirtinguose gyliuose. Sieros ciklas. Yra daug dujinių sieros junginių, tokių kaip vandenilio sulfidas H2S ir sieros dioksidas SO2. Tačiau vyraujanti šio elemento ciklo dalis yra nuosėdinio pobūdžio ir atsiranda dirvožemyje bei vandenyje.Neorganinės sieros prieinamumą ekosistemoje palengvina geras daugelio sulfatų tirpumas vandenyje. Augalai, absorbuodami sulfatus, juos redukuoja ir gamina sieros turinčias aminorūgštis (metioniną, cisteiną, cistiną), kurios vaidina svarbų vaidmenį formuojant tretinę baltymų struktūrą formuojant disulfidinius tiltelius tarp skirtingų polipeptidinės grandinės zonų. Aiškiai matomos daugelis pagrindinių biogeocheminio ciklo ypatybių: S -> S -> SO2 - bespalvės, žalios ir violetinės sieros bakterijos; - "H2S (anaerobinis sulfato redukavimas) - Desulfovibno; H2S - "SO2" (aerobinė sulfido oksidacija) - tiobacila; organinis S SO ir H2S. atitinkamai aerobiniai ir anaerobiniai heterotrofiniai mikroorganizmai. Pirminė gamyba užtikrina sulfato įtraukimą į organines medžiagas, o gyvūnų išskyrimas yra būdas grąžinti sulfatą į ciklą. Apskritai ekosistemai reikia mažiau sieros nei azoto ir fosforo. Taigi siera rečiau yra ribojantis veiksnys augalams ir gyvūnams. Tuo pačiu metu sieros ciklas yra pagrindinis viso biomasės gamybos ir skaidymo proceso veiksnys. Pavyzdžiui, kai nuosėdose susidaro geležies sulfidai, fosforas iš netirpios formos pereina į tirpią ir tampa prieinamas organizmams. Tai patvirtinimas, kaip vieną ciklą reguliuoja kitas. 2 Energijos srautas ekosistemose Skirtingai nuo medžiagų, kurios nuolat cirkuliuoja per skirtingus ekosistemos blokus, kurias visada galima panaudoti pakartotinai ir patekti į ciklą, energija gali būti panaudota vieną kartą, t.y. vyksta linijinis energijos srautas per ekosistemą. Vienpusis energijos antplūdis, kaip universalus gamtos reiškinys, atsiranda dėl termodinamikos dėsnių. Pirmasis dėsnis teigia, kad energiją galima paversti iš vienos formos (pavyzdžiui, šviesos) į kitą (pavyzdžiui, potencialią maisto energiją), tačiau jos negalima sukurti ar sunaikinti. Antrasis dėsnis teigia, kad negali būti vieno proceso, susieto su energijos transformacija, neprarandant jos dalies. Tam tikras energijos kiekis tokiose transformacijose išsisklaido į neprieinamą šiluminę energiją, todėl prarandama. Vadinasi: negali būti 100% efektyvumo, pavyzdžiui, maisto medžiagų transformacijos į organizmo kūną sudarančią medžiagą. Taigi gyvi organizmai yra energijos keitikliai. Ir kiekvieną kartą, kai energija virsta, dalis jos prarandama šilumos pavidalu. Galiausiai visa energija, patenkanti į biotinį ekosistemos ciklą, išsisklaido kaip šiluma. Gyvi organizmai iš tikrųjų nenaudoja šilumos kaip energijos šaltinio darbui atlikti – jie naudoja šviesą ir cheminę energiją. Maisto grandinės ir tinklai, trofiniai lygiai. Ekosistemoje energijos turinčias medžiagas sukuria autotrofiniai organizmai ir jos yra heterotrofų maistas. Maisto ryšiai – tai energijos perdavimo iš vieno organizmo į kitą mechanizmai. Tipiškas pavyzdys: gyvūnas valgo augalus. Šį gyvūną, savo ruožtu, gali valgyti kitas gyvūnas. Tokiu būdu energija gali būti perduodama per daugybę organizmų – kiekvienas paskesnis minta ankstesniuoju, kuris aprūpina jį žaliavomis ir energija. Ši energijos perdavimo seka vadinama maisto (trofine) grandine arba maisto grandine. Kiekvienos maisto grandinės grandies vieta yra trofinis lygis. Pirmąjį trofinį lygį, kaip minėta anksčiau, užima autotrofai arba vadinamieji pirminiai gamintojai. Antrojo trofinio lygio organizmai vadinami pirminiais vartotojais, trečiojo – antriniais vartotojais ir kt. Paprastai yra trijų tipų maisto grandinės.Mėsėdžių maisto grandinė prasideda nuo augalų ir pereina nuo mažų organizmų prie vis didesnių organizmų. Sausumoje maisto grandinės susideda iš trijų ar keturių grandžių. Viena iš paprasčiausių maisto grandinių atrodo taip: augalas -> kiškis -> vilkas gamintojas -" žolėdis -> -> mėsėdis Taip pat plačiai paplitusios šios maisto grandinės: Augalinė medžiaga (pvz., nektaras) - "musė -" -" voras -> vėgėlė -> pelėda. Rožių krūmo sula -> amaras -> -> boružėlė -> -> voras - "vabzdžiaėdis paukštis -> plėšrus paukštis. Vandens ir ypač jūrų ekosistemose plėšrūnų mitybos grandinės paprastai būna ilgesnės nei sausumos. Trečiojo tipo mitybos grandinės, pradedant negyvų augalų liekanomis, skerdenomis ir gyvūnų ekskrementais, vadinamos detritinėmis (saprofitinėmis) maisto grandinėmis arba detritalų skilimo grandinėmis. Lapuočių miškai vaidina svarbų vaidmenį sausumos ekosistemų detrito maisto grandinėse, kurių didžioji dalis lapijos nesuvalgoma žolėdžių ir yra nukritusių lapų pakratų dalis. Lapus susmulkina daugybė detritivorių – grybų, bakterijų, vabzdžių (pavyzdžiui, kolembolų) ir kt., o vėliau juos suryja sliekai, kurie tolygiai paskirsto humusą paviršiniame žemės sluoksnyje, sudarydami vadinamąjį mulą. Šiame lygyje grybai vystosi grybiena. Skildami mikroorganizmai, užbaigiantys grandinę, gamina galutinę negyvų organinių medžiagų mineralizaciją. Apskritai mūsų miškams būdingos detrito mitybos grandinės gali būti pavaizduotos taip: Lapų kraikas -> sliekas -> juodvarnis - žvirblis; Nugaišęs gyvūnas – „medžių musių lervos –“ žolinė varlė –> paprastoji žolinė gyvatė. Aptartose maisto grandinės diagramose kiekvienas organizmas vaizduojamas kaip maitinantis kitais tos pačios rūšies organizmais. Tikrieji maisto ryšiai ekosistemoje yra daug sudėtingesni, nes gyvūnas gali maitintis skirtingų tipų organizmais iš tos pačios mitybos grandinės arba iš skirtingų mitybos grandinių, pavyzdžiui, viršutinių trofinių lygių plėšrūnais. Gyvūnai dažnai minta ir augalais, ir kitais gyvūnais. Jie vadinami visaėdžiais. Maisto tinklai ekosistemose yra labai sudėtingi, todėl galime daryti išvadą, kad į jas patenkanti energija ilgai migruoja iš vieno organizmo į kitą. Ekologinės piramidės. Kiekvienoje ekosistemoje maisto tinklai turi aiškiai apibrėžtą struktūrą, kuriai būdinga organizmų, atstovaujamų kiekviename įvairių maisto grandinių lygmenyje, pobūdžiu ir skaičiumi. Norėdami ištirti organizmų ryšius ekosistemoje ir pavaizduoti juos grafiškai, jie dažniausiai naudoja ekologines piramides, o ne maisto tinklo diagramas. Ekologinės piramidės geometrine forma išreiškia ekosistemos trofinę struktūrą. Jie sukonstruoti tokio paties pločio stačiakampių pavidalu, tačiau stačiakampių ilgis turi būti proporcingas matuojamo objekto vertei. Iš čia galite gauti skaičių, biomasės ir energijos piramides. Ekologinės piramidės atspindi pagrindines bet kokios biocenozės savybes, kai parodo jos trofinę struktūrą. Skaičių piramidės. Jie yra paprasčiausias ekosistemos trofinės struktūros tyrimo priartėjimas. Biomasės piramidė. Tiksliau atspindi mitybos santykius ekosistemoje, nes atsižvelgiama į bendrą kiekvieno trofinio lygio organizmų masę (biomasę). Energijos piramidė. Svarbiausias būdas parodyti ryšius tarp skirtingų trofinių lygių organizmų yra energijos piramidės. Jie atspindi maisto grandinių energijos konversijos efektyvumą ir produktyvumą ir yra sudaryti skaičiuojant energijos kiekį (kcal), sukauptą paviršiaus vienetui per laiko vienetą ir sunaudojamą organizmų kiekviename trofiniame lygmenyje. Augalo gaunama saulės energija fotosintezės procese panaudojama tik iš dalies. Angliavandeniuose fiksuota energija atspindi bendrą ekosistemos gamybą. Angliavandeniai naudojami protoplazmai kurti ir augalų augimui. Dalis jų energijos skiriama kvėpavimui. Antros eilės vartotojai (plėšrūnai) nesunaikina visos savo aukų biomasės. Be to, tik dalis jų sunaikinamo kiekio panaudojama jų pačių trofinio lygio biomasei sukurti. Likusi dalis daugiausia išleidžiama kvėpavimo energijai ir išsiskiria su ekskrementais ir ekskrementais. 1942 metais R. Lindemanas pirmą kartą suformulavo energijų piramidės dėsnį, kuris vadovėliuose dažnai vadinamas „10% dėsniu“. Pagal šį dėsnį vidutiniškai iš vieno ekologinės piramidės trofinio lygio į kitą pereina ne daugiau kaip 10 % energijos. Tik 10-20% pradinės energijos perduodama vėlesniems heterotrofams. Taikant energijos piramidės dėsnį, nesunku apskaičiuoti, kad tretinius mėsėdžius (trofinis lygis V) pasiekiančios energijos kiekis yra apie 0,0001 gamintojų absorbuojamos energijos. Iš to išplaukia, kad energijos perkėlimas iš vieno lygio į kitą vyksta labai mažu efektyvumu. Tai paaiškina ribotą grandžių skaičių maisto grandinėje, nepaisant konkrečios biocenozės. E. Odumas (1959) itin supaprastintoje mitybos grandinėje - liucerna -> veršelis - "vaikas įvertino energijos virsmą, iliustravo jos nuostolių mastą. Tarkime, samprotavo jis, liucernos pasėlių yra plote, 4 ha.Šiame lauke minta veršeliai (manoma, kad minta tik liucerna), o 12 metų berniukas – vien veršienos.Skaičiavimų rezultatai pateikti trijų piramidžių pavidalu: skaičiai, biomasė ir energija nurodo, kad liucerna sunaudoja tik 0,24 % visos į lauką patenkančios saulės energijos, veršelis sugeria 8 % šio produkto ir tik 0,7 % veršelio biomasės užtikrina vaiko vystymąsi per metus. Taigi E. Odumas parodė, kad tik viena milijoninė saulės energijos dalis paverčiama mėsėdžių biomase, šiuo atveju prisidedant prie vaiko svorio padidėjimo, o likusi dalis prarandama ir suardyta forma išsisklaido aplinkoje. Aukščiau pateiktas pavyzdys aiškiai iliustruoja labai žemą ekologinį ekosistemų efektyvumą ir žemą transformacijos maisto grandinėse efektyvumą. Galime teigti: jei 1000 kcal (dieną m 2) užfiksavo gamintojai, tada 10 kcal (dieną m 2) patenka į žolėdžių gyvūnų biomasę ir tik 1 kcal (dieną m 2) – į mėsėdžių biomasę. Kadangi tam tikrą medžiagos kiekį kiekviena biocenozė gali panaudoti pakartotinai, o dalį energijos – vieną kartą, tikslingiau sakyti, kad ekosistemoje vyksta kaskadinis energijos perdavimas. Vartotojai yra valdymo ir stabilizavimo grandis ekosistemoje. Vartotojai sukuria cenozės įvairovės spektrą, užkertant kelią dominuojančių asmenų monopolijai. Kontroliuojančios vartotojų vertės taisyklė pagrįstai gali būti laikoma gana pagrindine. Remiantis kibernetikos pažiūromis, valdymo sistema turėtų būti sudėtingesnės struktūros nei valdoma, tada paaiškėja instrumentų tipų įvairovės priežastis. Kontroliuojanti vartotojų reikšmė turi ir energetinį pagrindą. Energijos srauto per vieną ar kitą trofinį lygį negali visiškai nulemti maisto prieinamumas pagrindiniame trofiniame lygyje. Kaip žinoma, visada lieka pakankamai „rezervo“, nes visiškas maisto sunaikinimas lemtų vartotojų mirtį. Šie bendrieji modeliai stebimi populiacijos procesų, bendruomenių, ekologinės piramidės lygių ir visos biocenozių kontekste. 3 Dinamiški procesai, užtikrinantys ekosistemų vientisumą ir tvarumą Cikliniai bendrijų pokyčiai atspindi kasdienį, sezoninį ir ilgalaikį išorinių sąlygų periodiškumą bei endogeninių organizmų ritmų pasireiškimą. Kasdienė ekosistemų dinamika daugiausia siejama su gamtos reiškinių ritmu ir yra griežtai periodinio pobūdžio. Jau svarstėme, kad kiekvienoje biocenozėje yra organizmų grupių, kurių gyvybė vyksta skirtingu paros metu. Vieni aktyvūs dieną, kiti – naktį. Vadinasi, periodiškai keičiasi tam tikros ekosistemos atskirų biocenozės tipų sudėtis ir santykis, nes atskiri organizmai tam tikram laikui iš jos išjungiami. Kasdienę biocenozės dinamiką užtikrina ir gyvūnai, ir augalai. Kaip žinoma, fiziologinių procesų intensyvumas ir pobūdis augaluose kinta dienos metu – naktį fotosintezė nevyksta, dažnai žiedai augaluose atsiveria tik naktį ir juos apdulkina naktiniai gyvūnai, kiti prisitaikę apdulkinti dieną. Dienos dinamika biocenozėse, kaip taisyklė, yra ryškesnė, tuo didesnis temperatūros, drėgmės ir kitų aplinkos veiksnių skirtumas tarp dienos ir nakties. Esant sezoninei dinamikai, pastebimi reikšmingesni biocenozių nuokrypiai. Taip yra dėl biologinių organizmų ciklų, kurie priklauso nuo sezoninio gamtos reiškinių cikliškumo. Taigi sezonų kaita daro didelę įtaką gyvūnų ir augalų gyvenimo veiklai (žiemos miegas, žiemos miegas, žvėrių svyravimai ir migracija; augalų žydėjimo, derėjimo, aktyvaus augimo, lapų kritimo ir žiemos ramybės periodai). Pakopinė biocenozės struktūra dažnai priklauso nuo sezoniškumo. Atskiri augalų sluoksniai atitinkamais metų laikais gali visiškai išnykti, pavyzdžiui, žolinis sluoksnis, susidedantis iš vienmečių augalų. Biologinių sezonų trukmė įvairiose platumose skiriasi. Šiuo atžvilgiu sezoninė biocenozių dinamika Arkties, vidutinio klimato ir atogrąžų zonose skiriasi. Aiškiausiai tai išreikšta vidutinio klimato ekosistemose ir šiaurinėse platumose. Ilgalaikis kintamumas yra normalus bet kurios biocenozės gyvenime. Taigi kritulių kiekis Barabinsko miško stepėje kasmet smarkiai svyruoja, daug sausų metų keičiasi su ilgalaikiais gausių kritulių laikotarpiais. Tai daro didelę įtaką augalams ir gyvūnams. Tokiu atveju atsiranda ekologinių nišų plėtra – funkcinė demarkacija besiformuojančiame rinkinyje arba jo papildymas su maža įvairove. Ilgalaikiai biocenozių sudėties pokyčiai taip pat kartojasi dėl periodinių bendros atmosferos cirkuliacijos pokyčių, kuriuos savo ruožtu sukelia saulės aktyvumo padidėjimas arba sumažėjimas. Kasdienės ir sezoninės dinamikos procese biocenozių vientisumas paprastai nepažeidžiamas. Biocenozė patiria tik periodinius kokybinių ir kiekybinių savybių svyravimus. Progresuojantys ekosistemos pokyčiai galiausiai lemia, kad viena biocenozė pakeičiama kita, su skirtinga dominuojančių rūšių rinkiniu. Tokių pokyčių priežastys gali būti išoriniai biocenozei veiksniai, kurie ilgą laiką veikia viena kryptimi, pavyzdžiui, didėjanti vandens telkinių tarša, didėjantis pelkių dirvožemių džiūvimas dėl melioracijos, padidėjęs ganymas ir kt. Šie vienos biocenozės pokyčiai į kitą vadinami egzogenetiniais. Tuo atveju, kai didėjanti veiksnio įtaka lemia laipsnišką biocenozės struktūros supaprastinimą, jų sudėties išeikvojimą ir produktyvumo mažėjimą, tokie poslinkiai vadinami digresuojančiais arba mažėjančiais. Endogenetiniai pokyčiai atsiranda dėl procesų, vykstančių pačioje biocenozėje. Eilinis vienos biocenozės pakeitimas kita vadinama ekologine sukcesija (iš lot. – sukcesija – seka, kaita). Susekcija yra ekosistemų savaiminio vystymosi procesas. Perėjimas grindžiamas biologinio ciklo neužbaigtumu tam tikroje biocenozėje. Yra žinoma, kad gyvi organizmai dėl savo gyvybinės veiklos keičia juos supančią aplinką, pašalindami iš jos kai kurias medžiagas ir prisotindami medžiagų apykaitos produktais. Kai populiacijos egzistuoja gana ilgą laiką, jos keičia savo aplinką nepalankia linkme ir dėl to yra išstumtos kitų rūšių populiacijų, kurioms dėl to atsirandantys aplinkos pokyčiai yra ekologiškai naudingi. Taigi biocenozėje keičiasi dominuojančios rūšys. Čia aiškiai matoma ekologinio dubliavimo taisyklė (principas). Ilgalaikis biocenozės egzistavimas įmanomas tik tuo atveju, jei aplinkos pokyčiai, kuriuos sukelia vienų gyvų organizmų veikla, yra palankūs kitiems, kuriems keliami priešingi reikalavimai. Remiantis konkurencine rūšių sąveika sukcesijos metu, palaipsniui formuojasi stabilesni deriniai, atitinkantys specifines abiotines aplinkos sąlygas. Vienos bendruomenės pakeitimo kita pavyzdys yra nedidelio ežero užaugimas, po kurio atsiranda pelkė, o vėliau – miškas. Pirmiausia ežero pakraščiais suformuojamas plaukiojantis kilimas – plaukiojantis viksvų, samanų ir kitų augalų kilimas. Ežeras nuolat prisipildo negyvų augalų liekanų – durpių. Susidaro pelkė, pamažu apauganti mišku. Viena po kitos einančios bendruomenių serijos, palaipsniui ir natūraliai pakeičiančios viena kitą, vadinamos seka. Susekėjimai gamtoje yra labai įvairaus masto. Juos galima stebėti pakrantėse su kultūromis, kurios yra planktoninės bendrijos – įvairių rūšių plūduriuojantys dumbliai ir jų vartotojai – rotiferiai, žiogeliai balose ir tvenkiniuose, pelkėse, pievose, miškuose, apleistose ariamose žemėse, atvėsusiose uolose ir kt. Yra hierarchija. ekosistemų organizacijoje Tai pasireiškia ir sukcesijos procesais – didesnės biocenozių transformacijos susidaro iš smulkesnių. Stabiliose ekosistemose su reguliuojamu medžiagų ciklu taip pat nuolat vyksta vietiniai sukcesijos pokyčiai, palaikantys sudėtingą vidinę bendruomenių struktūrą. Seksualinių pokyčių tipai. Yra du pagrindiniai nuoseklių pokyčių tipai: 1 - dalyvaujant autotrofinėms ir heterotrofinėms populiacijoms; 2 - dalyvaujant tik heterotrofams. Antrojo tipo sukcesija atsiranda tik tokiomis sąlygomis, kai susidaro preliminarus organinių junginių tiekimas arba nuolatinis aprūpinimas, dėl kurio egzistuoja bendrija: mėšlo krūvose ar krūvose, irstančioje augalinėje medžiagoje, organinėmis medžiagomis užterštuose rezervuaruose ir kt. Perėjimo procesas. Pasak F. Clementso (1916), paveldėjimo procesas susideda iš šių etapų: Susekimas pasikeitus augmenijai gali būti pirminis ir antrinis. Pirminis sukcesija yra ekosistemų vystymosi ir kaitos procesas anksčiau negyvenamose teritorijose, pradedant jų kolonizavimu. Klasikinis pavyzdys yra nuolatinis plikų uolienų užteršimas ir galiausiai ant jų besiformuojantys miškai. Taigi Uralo kalnų uolose vykstančiose pirminėse sukcesijose išskiriami šie etapai: Antrinė sukcesija yra ekosistemos, kuri kažkada egzistavo tam tikroje vietovėje, atkūrimas. Jis prasideda, kai nusistovėjusioje biocenozėje sutrinka nusistovėję organizmų ryšiai dėl ugnikalnio išsiveržimo, gaisro, kirtimo, arimo ir pan. Poslinkiai, vedantys į ankstesnės biocenozės atkūrimą, geobotanikoje vadinami demutaciniais. Pavyzdys yra rūšių įvairovės dinamika Krakatau saloje po to, kai ugnikalnis visiškai sunaikino vietinę florą ir fauną. Kitas pavyzdys – antrinis Sibiro tamsiųjų spygliuočių miško (eglių-kedro taigos) sukcesija po niokojančio miško gaisro. Labiau išdegintose vietose pionierinės samanos atsiranda nuo vėjo perpučiamų sporų: praėjus 3-5 metams po gaisro, gausiausios „ugnies samanos“ yra Funaria hygrometrica, Geratodon purpureus ir kt. Iš aukštesnių augalų ugniažolė (Chamaenerion angustifolium) labai greitai. kolonizuoja išdegusias vietas ), kuri jau po 2-3 mėnesių gausiai žydi ugnyje, taip pat žemė nendrinė žolė (Calamagrostis epigeios) ir kitos rūšys. Pastebimos tolimesnės sukcesijos fazės: nendrinė pieva užleidžia vietą krūmams, tada beržynams ar drebulynams, mišriam pušynui, pušynui, pušynui ir kedro miškui, o galiausiai po 250 metų atsistato kedro eglynas. . Antrinė sukcesija, kaip taisyklė, vyksta greičiau ir lengviau nei pirminė, nes pažeistoje buveinėje išsaugomas dirvožemio profilis, sėklos, pradmenys ir dalis ankstesnės populiacijos bei ankstesnės jungtys. Demutacija nėra jokio pirminio paveldėjimo etapo kartojimas. Climax ekosistema. Sukcesija baigiasi etapu, kai visos ekosistemos rūšys, besidaugindamos, išlaiko santykinai pastovų skaičių ir daugiau jos sudėties nekeičiama. Ši pusiausvyros būsena vadinama kulminacija, o ekosistema – kulminacija. Esant skirtingoms abiotinėms sąlygoms, susidaro skirtingos kulminacijos ekosistemos. Karštame ir drėgname klimate tai bus atogrąžų miškas, sausame ir karštame – dykuma. Pagrindiniai žemės biomai yra jų atitinkamų geografinių sričių kulminacijos ekosistemos. Ekosistemos pokyčiai sukcesijos metu. Produktyvumas ir biomasė. Kaip jau minėta, sukcesija yra natūralus, kryptingas procesas, o viename ar kitame etape vykstantys pokyčiai yra būdingi bet kuriai bendrijai ir nepriklauso nuo jos rūšinės sudėties ar geografinės padėties. Yra keturi pagrindiniai nuoseklių pakeitimų tipai: Pažymėtina ir tai, kad paveldėjimo fazės keičiasi laikantis tam tikrų taisyklių. Kiekviena fazė paruošia aplinką kito atsiradimui. Čia veikia vystymosi fazių eigos dėsnis; natūralios sistemos vystymosi fazės gali sekti tik evoliuciškai fiksuota (istoriškai, aplinkosaugos nulemta) tvarka, paprastai nuo gana paprastos iki sudėtingos, kaip taisyklė, be nuostolių. tarpinių stadijų, bet galbūt su labai greitu jų praeinimu arba evoliuciškai fiksuotu nebuvimu. Kai ekosistema artėja prie menopauzės, joje, kaip ir visose pusiausvyros sistemose, visi vystymosi procesai sulėtėja. Šią situaciją atspindi nuoseklumo sulėtėjimo dėsnis: procesai, vykstantys brandžiose pusiausvyros ekosistemose, kurios yra stabilios būklės, paprastai yra linkusios lėtėti. Tokiu atveju atkuriamasis paveldėjimo tipas keičiasi į jų pasaulietinę eigą, t.y. savaiminis vystymasis vyksta menopauzės ar mazgų vystymosi ribose. Empirinis nuoseklumo sulėtėjimo dėsnis yra G. Odumo ir R. Pinkertono taisyklės arba maksimalios energijos taisyklės, skirtos išlaikyti brandžią sistemą, pasekmė: seka vyksta esminio energijos srauto poslinkio link jos didinimo. kiekis, skirtas sistemai palaikyti. G. Odumo ir R. Pinkertono taisyklė savo ruožtu remiasi A. Lotkos suformuluota maksimalios energijos biologinėse sistemose taisykle. Šį klausimą vėliau gerai išplėtojo R. Margalef, Y. Odum ir jis žinomas kaip „nulinio maksimumo“ principo arba augimo sumažinimo brandžioje ekosistemoje įrodymas: nuosekliai besivystanti ekosistema linkusi formuoti didžiausią biomasę mažiausias biologinis produktyvumas. Lindemanas (1942) eksperimentiškai įrodė, kad paveldėjimą lydi produktyvumo padidėjimas iki kulminacijos bendruomenės, kurioje energijos konversija vyksta efektyviausiai. Ąžuolynų ir ąžuolynų miškų sukcesijos tyrimų duomenys rodo, kad vėlesniuose tarpsniuose jų produktyvumas iš tiesų didėja. Tačiau perėjus į kulminacijos bendruomenę, paprastai sumažėja bendras produktyvumas. Taigi produktyvumas senuose miškuose yra mažesnis nei jaunuolynų, kurie savo ruožtu gali būti mažiau produktyvūs nei prieš juos esantys rūšimis turtingesni žoliniai sluoksniai. Panašus produktyvumo sumažėjimas buvo pastebėtas kai kuriose vandens sistemose. Tam yra keletas priežasčių. Viena iš jų yra ta, kad augančioje miško biomasėje susikaupus maistinėms medžiagoms gali sumažėti jų ciklas. Bendro produktyvumo mažėjimas gali būti tiesiog sumažėjusio asmenų gyvybingumo rezultatas, nes didėja jų vidutinis amžius bendruomenėje. Vykstant sukcesijai, bendrijos biomasėje kaupiasi vis didesnė turimų maisto medžiagų dalis, atitinkamai mažėja jų kiekis abiotiniame ekosistemos komponente (dirvožemyje ar vandenyje). Taip pat didėja susidarančių detritų kiekis. Pagrindiniai pirminiai vartotojai yra ne žolėdžiai, o detritivoriai organizmai. Atitinkami pokyčiai vyksta ir trofiniuose tinkluose. Detritas tampa pagrindiniu maistinių medžiagų šaltiniu. Vykstant sukcesijai, didėja medžiagų biogeocheminių ciklų uždarumas. Maždaug 10 metų nuo augalijos dangos atkūrimo pradžios ciklų atvirumas sumažėja nuo 100 iki 10%, o vėliau dar labiau sumažėja, pasiekdamas minimumą kulminacijos fazėje. Galima drąsiai teigti, kad didėjančio medžiagų biogeocheminio ciklo uždarumo sukcesijos metu taisyklę pažeidžia antropogeninė augalijos ir natūralių ekosistemų transformacija apskritai. Be jokios abejonės, tai lemia ilgą biosferos ir jos padalijimo anomalijų seriją. Rūšių įvairovės sumažėjimas menopauzės metu nereiškia, kad jos ekologinė reikšmė maža. Rūšių įvairovė formuoja sukcesiją, jos kryptį ir užtikrina, kad tikroji erdvė prisipildytų gyvybės. Nepakankamas rūšių, sudarančių kompleksą, skaičius negalėjo sudaryti eilės, ir palaipsniui, sunaikinus kulminacijos ekosistemas, planeta dykumėtų. Įvairovės reikšmė yra funkcionali ir statiškai, ir dinamiškai. Pažymėtina, kad kai rūšių įvairovės nepakanka, kad susidarytų biosfera, kuri yra normalios natūralios sukcesijos proceso eigos pagrindas, o pati aplinka yra smarkiai sutrikdyta, sukcesija nepasiekia kulminacijos fazės, o baigiasi mazgine bendruomene – paraklimaksa, ilgalaike arba trumpalaike išvestine bendruomene. Kuo gilesnis tam tikros erdvės aplinkos sutrikimas, tuo ankstesnės fazės baigiasi. Kai dėl jų sunaikinimo (antropogeninio buveinių nykimo, rečiau išnykimo) netenkama viena ar grupė rūšių, menopauzės pasiekimas nėra visiškas natūralios aplinkos atkūrimas. Tiesą sakant, tai nauja ekosistema, nes joje atsirado naujų ryšių, dingo daug senų, susiformavo kitoks rūšių „nušrimas“. Ekosistema negali grįžti į savo senąją būseną, nes prarastos rūšies negalima atkurti. Pasikeitus bet kuriam abiotiniam ar biotiniam veiksniui, pavyzdžiui, dėl nuolatinio aušinimo arba įvedant naują rūšį, rūšis, kuri yra prastai prisitaikiusi prie naujų sąlygų, susidurs su vienu iš trijų būdų. Evoliucinio-ekologinio negrįžtamumo dėsnis teigia, kad ekosistema, kuri dėl ekologinių komponentų disbalanso prarado dalį savo elementų arba buvo pakeista kitu, sukcesijos metu negali grįžti į pradinę būseną, jei pokyčių metu evoliucinė (mikroevoliucinė) ) įvyko ekologinių elementų pakitimų (išsaugotų arba laikinai prarastų). Tuo atveju, kai kai kurios rūšys prarandamos tarpinėse sukcesijos fazėse, šį praradimą galima funkciškai kompensuoti, bet ne visiškai. Kai įvairovė mažėja virš kritinio lygio, sekimo eiga iškreipiama ir iš tikrųjų neįmanoma pasiekti kulminacijos, identiškos praeičiai. Norint įvertinti atkurtų ekosistemų prigimtį, svarbus evoliucinio-ekologinio negrįžtamumo dėsnis. Praradus elementus, tai iš tikrųjų yra visiškai nauji ekologiški gamtos dariniai su naujai susiformavusiais raštais ir ryšiais. Taigi, rūšies, kuri iškrito iš ekosistemos reaklimatizacijos metu, perkėlimas į praeitį nereiškia jos mechaninio sugrįžimo. Tai iš tikrųjų yra naujos rūšies įvedimas į atnaujintą ekosistemą.Evoliucinio-ekologinio negrįžtamumo dėsnis pabrėžia evoliucijos kryptį ne tik biosistemų lygmenyje, bet ir visuose kituose hierarchiniuose biotos lygiuose. 4 Biosfera kaip pasaulinė ekosistema, užtikrinanti ekosistemų vientisumą ir tvarumą Biosfera yra pasaulinė ekosistema. Kaip minėta anksčiau, biosfera skirstoma į geobiosferą, hidrobiosferą ir aerobiosferą (2.4 pav.). Geobiosfera skirstoma pagal pagrindinius aplinką formuojančius veiksnius: terrabiosfera ir litobiosfera – geobiosferoje, marinobiosfera (okeanobiosfera) ir akvabiosfera – hidrobiosferoje. Šie dariniai vadinami subsferomis. Pagrindinis aplinką formuojantis veiksnys jų formavimuisi yra fizinė gyvenamosios aplinkos fazė: oras-vanduo aerobiosferoje, vanduo - gėlas ir sūrus vanduo hidrobiosferoje, kietas oras terabiosferoje ir kietas vanduo litobiosferoje. Savo ruožtu jie visi patenka į sluoksnius: aerobiosfera – į tropobiosferą ir altobiosferą; hidrobiosfera – į fotosferą, disfotosferą ir afotosferą. Struktūrą formuojantys veiksniai čia, be fizinės aplinkos, energijos (šviesos ir šilumos), ypatingų gyvybės formavimosi ir evoliucijos sąlygų – evoliucinės biotos skverbimosi į žemę, į jos gelmes, į erdves virš žemės. Žemė, vandenyno bedugnė, neabejotinai skiriasi. Kartu su apobiosfera, parabiosfera ir kitais sub- ir viršbiosferos sluoksniais jie sudaro vadinamąjį „gyvybės sluoksnį“ ir jos egzistavimo geosferą (ekosferą) megabiosferos ribose. Sistemine prasme išvardyti dariniai yra didelės funkcinės, praktiškai universalių arba subplanetinių matmenų dalys. Mokslininkai mano, kad biosferoje yra mažiausiai 8-9 lygiai santykinai nepriklausomų medžiagų ciklų, susijusių su 7 pagrindinių medžiagų ir energijos ekologinių komponentų jungtimi, o 8-asis - informacinis. Globalūs, regioniniai ir vietiniai medžiagų ciklai nėra uždari ir iš dalies „kertasi“ ekosistemų hierarchijoje. Ši materialinė ir energetinė, o iš dalies ir informacinė „sujungimas“ užtikrina ekologinių supersistemų vientisumą iki pat visos biosferos. Ekosistemų vientisumas ir tvarumas. Biosferą labiau formuoja ne išoriniai veiksniai, o vidiniai modeliai. Svarbiausia biosferos savybė yra gyvų ir negyvų daiktų sąveika, kurią atspindi V. I. Vernadskio atomų biogeninės migracijos dėsnis. Biogeninės atomų migracijos dėsnis leidžia žmonijai sąmoningai valdyti biogeocheminius procesus tiek visoje Žemėje, tiek jos regionuose. Gyvos medžiagos kiekis biosferoje, kaip žinoma, nepastebimai keičiasi. Šį modelį gyvosios medžiagos kiekio pastovumo dėsnio forma suformulavo V. I. Vernadskis: gyvosios medžiagos kiekis biosferoje tam tikru geologiniu laikotarpiu yra pastovus. Praktiškai šis dėsnis yra kiekybinė vidinės dinaminės pusiausvyros dėsnio pasekmė globaliai ekosistemai – biosferai. Kadangi gyvoji medžiaga pagal biogeninės atomų migracijos dėsnį yra energijos tarpininkė tarp Saulės ir Žemės, jos kiekis arba turi būti pastovus, arba turi keistis energetinės charakteristikos. Gyvosios medžiagos fizinės ir cheminės vienybės dėsnis (visa gyvoji Žemės medžiaga yra fiziškai ir chemiškai vieninga ir neįtraukia reikšmingų pastarosios savybės pokyčių. Vadinasi, kiekybinis stabilumas planetos gyvajai medžiagai yra neišvengiamas. Jis visiškai būdingas rūšių skaičius. Gyvoji medžiaga, kaip saulės energijos kaupiklis, turi vienu metu reaguoti ir į išorinius (kosminius) poveikius, ir į vidinius pokyčius. Gyvosios medžiagos kiekio sumažėjimas arba padidėjimas vienoje biosferos vietoje turėtų sukelti visiškai priešingą procesą kitoje vietoje, nes išsiskiriančias maistines medžiagas gali pasisavinti likusi gyvoji medžiaga arba bus pastebėtas jų trūkumas. Čia reikia atsižvelgti į proceso greitį, kuris antropogeninės kaitos atveju yra daug mažesnis nei tiesioginis žmogaus trikdymas gamtai. Be gyvosios medžiagos kiekio pastovumo ir pastovumo, kuris atsispindi gyvosios medžiagos fizinės ir cheminės vienybės dėsnyje, gyvojoje gamtoje nuolat išsaugoma informacinė ir somatinė struktūra, nepaisant to, kad ji keičiasi. šiek tiek su evoliucijos eiga. Šią savybę pastebėjo Yu.Goldsmithas (1981) ir ją pavadino biosferos sandaros – informacinio ir somatinio – išsaugojimo dėsniu arba pirmuoju ekodinamikos dėsniu. Siekdami išsaugoti biosferos struktūrą, gyviai stengiasi pasiekti brandos būseną arba ekologinę pusiausvyrą. Menopauzės troškimo dėsnis – antrasis Yu.Goldsmitho ekodinamikos dėsnis, taikomas biosferai ir kitiems ekologinių sistemų lygmenims, nors yra specifikos – biosfera yra uždaresnė sistema nei jos padaliniai. Gyvosios biosferos materijos vienovė ir jos posistemių struktūros homologija lemia tai, kad joje atsiradę skirtingų geologinių amžių ir originalios geografinės kilmės gyvieji elementai yra evoliuciškai sudėtingai susipynę. Skirtingos erdvės ir laiko genezės elementų susipynimas visuose ekologiniuose biosferos lygiuose atspindi gyvosios medžiagos heterogenezės taisyklę arba principą. Šis papildymas nėra chaotiškas, jam galioja ekologinio papildomumo, ekologinio atitikimo (kongruencijos) principai ir kiti dėsniai. Yu. Goldsmitho ekodinamikos rėmuose tai yra trečiasis jos dėsnis – ekologinės tvarkos principas, arba ekologinis abipusiškumas, nurodantis visuotinę savybę dėl visumos įtakos jos dalims, atvirkštinę diferencijuotų dalių įtaką visumos vystymąsi ir pan., o tai iš viso lemia visos biosferos išsaugojimo stabilumą. Abipusę pagalbą ekologinės tvarkos rėmuose, arba sisteminį abipusiškumą, patvirtina erdvės užpildymo tvarkingumo ir erdvės-laikinio tikrumo dėsnis: erdvės užpildymas natūralioje sistemoje dėl jos posistemių sąveikos yra įsakomas tokiu būdu, kuris leidžia realizuoti homeostatines sistemos savybes su minimaliais prieštaravimais tarp joje esančių dalių. Iš šio dėsnio išplaukia, kad gamtai „nereikalingų“ nelaimingų atsitikimų, įskaitant jai svetimus žmogaus kūrinius, ilgalaikis egzistavimas yra neįmanomas. Biuzialistinės sistemų tvarkos taisyklės biosferoje taip pat apima sistemų komplementarumo principą, teigiantį, kad vienos natūralios sistemos posistemės savo raidoje sudaro prielaidą sėkmingam kitų į tą pačią sistemą įtrauktų posistemių vystymuisi ir savireguliacijai. Ketvirtasis Yu. Goldsmitho ekodinamikos dėsnis apima gyvų būtybių savikontrolės ir savireguliacijos dėsnį: gyvos sistemos ir sistemos, veikiančios kontroliuojančią gyvų būtybių įtaką, savo procese sugeba susitvardyti ir reguliuotis. prisitaikymas prie aplinkos pokyčių. Biosferoje savikontrolė ir savireguliacija vyksta bendrosios sąveikos kaskadinių ir grandininių procesų metu – kovojant už natūralios atrankos egzistavimą (plačiausia šios sąvokos prasme), adaptuojant sistemas ir posistemes, vykstant plačiajai koevoliucijai. ir kt. Be to, visi šie procesai lemia teigiamus rezultatus „gamtos požiūriu“ - biosferos ir visos jos ekosistemų išsaugojimą ir vystymąsi. Jungiamoji grandis tarp struktūrinio ir evoliucinio pobūdžio apibendrinimų yra automatinio globalios buveinės palaikymo taisyklė: gyvoji medžiaga savireguliacijos ir sąveikos su abiotiniais veiksniais eigoje autodinamiškai palaiko jai vystytis tinkamą gyvybės aplinką. Šį procesą riboja kosminės ir globalios ekosferos masto pokyčiai ir jis vyksta visose planetos ekosistemose ir biosistemose kaip savireguliacijos kaskada, pasiekianti pasaulinį mastą. Automatinio globalios buveinės priežiūros taisyklė išplaukia iš V.I. biogeocheminių principų. Vernadsky, rūšių buveinių išsaugojimo taisyklės, santykinis vidinis nuoseklumas ir tarnauja kaip konservatyvių mechanizmų buvimo biosferoje konstanta ir tuo pačiu patvirtina sistemos dinaminio papildomumo taisyklę. Kosminį poveikį biosferai liudija kosminių poveikių lūžio dėsnis: kosminius veiksnius, turinčius įtakos biosferai ir ypač jos padaliniams, keičia planetos ekosfera, todėl jų stiprumas ir laikas. , apraiškos gali susilpnėti ir pasislinkti arba net visiškai prarasti savo poveikį. Apibendrinimas čia svarbus dėl to, kad dažnai vyksta sinchroninis Saulės aktyvumo ir kitų kosminių veiksnių poveikis Žemės ekosistemoms ir joje gyvenantiems organizmams. Pažymėtina, kad daugelis procesų Žemėje ir jos biosferoje, nors ir priklausomi nuo kosmoso, saulės aktyvumo ciklai laikomi 1850, 600, 400, 178, 169, 88, 83, 33, 22, 16 intervalais. , 11, 5 (11 ,1), 6,5 ir 4,3 metų, pati biosfera ir jos skyriai nebūtinai visais atvejais turi reaguoti vienodai cikliškai. Kosminis biosferos sistemos poveikis gali būti visiškai arba iš dalies užblokuotas. Išvada Ekosistemos yra pagrindinis funkcinis ekologijos vienetas, nes jas sudaro organizmai ir negyvoji aplinka – komponentai, kurie abipusiškai įtakoja vienas kito savybes ir būtinas sąlygas gyvybei palaikyti tokia forma, kokia egzistuoja Žemėje. Konkrečios fizikinės ir cheminės aplinkos (biotopo) derinys su gyvų organizmų bendruomene (biocenozė) sudaro ekosistemą. Yra mikroekosistemos, mezoekosistemos ir pasaulinė ekosistema – biosfera. Ekosistemos nėra išsibarsčiusios netvarkoje, priešingai, jos yra sugrupuotos į gana taisyklingas zonas tiek horizontaliai (platumos), tiek vertikaliai (aukštyje). Pagrindinis ekosistemų energijos šaltinis yra Saulė. Saulės spinduliuotė, patenkanti į Žemės paviršių, yra 1,54 mln. EJ per metus. Didžioji dalis saulės energijos, pasiekiančios planetos paviršių, tiesiogiai paverčiama šiluma, šildančiu vandeniu arba dirvožemiu, o tai savo ruožtu sušildo orą. Ekosistemos egzistuoja dėl neteršiančios ir beveik amžinos saulės energijos, kurios kiekis yra gana pastovus ir gausus. Saulės energija Žemėje sukelia du medžiagų ciklus: didelius, arba geologinius, ir mažus, biologinius (biotinius). Abu ciklai yra tarpusavyje susiję ir atspindi tarsi vieną procesą. Biogeocheminių ciklų, arba biogeocheminių ciklų, egzistavimas sukuria galimybę sistemos savireguliacijai (homeostazei), o tai suteikia ekosistemai stabilumo: nuostabų įvairių elementų procentinio kiekio pastovumą. Yra gana daug medžiagų ciklų. Reikšmingiausias ciklas Žemėje pagal perkeliamas mases ir energijos suvartojimą yra planetos hidrologinis ciklas arba vandens ciklas. Biotinis (biologinis) ciklas reiškia medžiagų apykaitą tarp dirvožemio, augalų, gyvūnų ir mikroorganizmų. Iš visų biogeocheminių ciklų anglies ciklas, be jokios abejonės, yra intensyviausias. Didžioji dalis per geologines epochas susidariusio deguonies neliko atmosferoje, o buvo fiksuojama litosferos karbonatų, sulfatų, geležies oksidų ir kt. pavidalu. Ši masė yra 590x1014 tonų, palyginti su 39x1014 tonų deguonies, kuris cirkuliuoja biosferoje. dujų arba sulfatų pavidalu, ištirpusių žemyniniuose ir vandenynų vandenyse. Azotas yra būtinas biogeninis elementas, nes jis yra baltymų ir nukleorūgščių dalis. Azoto ciklas yra vienas sudėtingiausių, nes apima ir dujų, ir mineralų fazes, o kartu ir pačius idealiausius ciklus. Fosforo ciklas biosferoje yra susijęs su medžiagų apykaitos procesais augaluose ir gyvūnuose. Šis svarbus ir reikalingas protoplazmos elementas, kurio sausumos augaluose ir dumbliuose yra 0,01–0,1%, gyvūnuose nuo 0,1% iki kelių procentų, cirkuliuoja, palaipsniui iš organinių junginių virsdamas fosfatais, kuriuos vėl gali naudoti augalai. Yra daug dujinių sieros junginių, tokių kaip vandenilio sulfidas H2S ir sieros dioksidas SO2. Organizmų gyvybinės veiklos ir medžiagų cirkuliacijos ekosistemose palaikymas, t.y., ekosistemų egzistavimas, priklauso nuo nuolatinio energijos srauto, reikalingo visiems organizmams jų gyvybinėms funkcijoms ir savaiminiam dauginimuisi. Skirtingai nuo medžiagų, kurios nuolat cirkuliuoja per skirtingus ekosistemos blokus, kurias visada galima pakartotinai panaudoti ir patekti į ciklą, energiją galima panaudoti vieną kartą, t.y., ekosistemoje vyksta linijinis energijos srautas. Ekosistemoje energijos turinčias medžiagas sukuria autotrofiniai organizmai ir jos yra heterotrofų maistas. Maisto ryšiai – tai energijos perdavimo iš vieno organizmo į kitą mechanizmai. Kiekvienoje ekosistemoje maisto tinklai turi aiškiai apibrėžtą struktūrą, kuriai būdinga organizmų, atstovaujamų kiekviename įvairių maisto grandinių lygmenyje, pobūdžiu ir skaičiumi. Ekosistemų formavimasis yra dinamiškas procesas. Ekosistemose nuolat kinta jų narių būklė ir gyvybinė veikla bei populiacijų santykis. Įvairūs pokyčiai, vykstantys bet kurioje bendruomenėje, skirstomi į du pagrindinius tipus: ciklinius ir progresyvius. Sukcesija yra natūralus, kryptingas procesas, o viename ar kitame etape vykstantys pokyčiai yra būdingi bet kuriai bendruomenei ir nepriklauso nuo jos rūšinės sudėties ar geografinės padėties. Pasaulinė ekosistema yra biosfera. Kaip minėta anksčiau, biosfera skirstoma į geobiosferą, hidrobiosferą ir aerobiosferą. Biosferą labiau formuoja ne išoriniai veiksniai, o vidiniai modeliai. Svarbiausia biosferos savybė yra gyvų ir negyvų daiktų sąveika, kuri atsispindi V.I. atomų biogeninės migracijos dėsnyje. Vernadskis. Žemėje egzistuojančios ekosistemos yra įvairios. Yra mikroekosistemos (pavyzdžiui, pūvančio medžio kamienas), mezoekosistemos (miškas, tvenkinys ir kt.), makroekosistemos (žemynas, vandenynas ir kt.) ir globali biosfera. Didelės sausumos ekosistemos vadinamos biomais. Kiekviename biome yra keletas mažesnių, tarpusavyje susijusių ekosistemų. Yra keletas ekosistemų klasifikacijų: visžaliai atogrąžų miškai; dykuma: žolinė ir krūminė; atogrąžų žolės ir savanos; vidutinio klimato stepė; vidutinio klimato lapuočių miškas: borealiniai spygliuočių miškai. Tundra: arktinė ir alpinė. Gėlavandenių ekosistemų tipai: juostelė (nejudantis vanduo): ežerai, tvenkiniai ir kt.; lotikai (tekantys vandenys): upės, upeliai ir kt.; pelkės: pelkės ir pelkėti miškai; Jūrų ekosistemų tipai: atvirasis vandenynas (pelaginė); kontinentinio šelfo vandenys (pakrančių vandenys); pakilimo plotai (derlingi plotai su produktyvia žvejyba); estuarijos (pakrančių įlankos, sąsiauriai, upių žiotys, druskingos pelkės ir kt.). Kiekvieną ekosistemą sudaro du pagrindiniai komponentai: organizmai ir veiksniai iš jų negyvosios aplinkos. Organizmų (augalų, gyvūnų, mikrobų) visuma vadinama ekosistemos biota. Skirtingų kategorijų organizmų sąveikos būdai yra jų biotinė struktūra. Saulės energija Žemėje sukelia du medžiagų ciklus: didelį, arba geologinį, aiškiausiai pasireiškiantį vandens cikle ir atmosferos cirkuliacijoje, ir mažąjį, biologinį (biotinį), besivystantį didelio ir susidedančio iš ištisinio, cikliškas, bet netolygus laike ir erdvėje, lydimas daugiau ar mažiau reikšmingų natūralaus materijos, energijos ir informacijos persiskirstymo įvairių organizavimo lygių ekologinėse sistemose nuostolių. Abu ciklai yra tarpusavyje susiję ir atspindi tarsi vieną procesą. Abiotinių veiksnių ir ekosistemos gyvų organizmų sąveiką lydi nuolatinė medžiagų cirkuliacija tarp biotopo ir biocenozės kintančių organinių ir mineralinių junginių pavidalu. Cheminių elementų mainai tarp gyvų organizmų ir neorganinės aplinkos, kurių įvairios stadijos vyksta ekosistemoje, vadinamas biogeocheminiu ciklu arba biogeocheminiu ciklu. Tokių ciklų egzistavimas sukuria galimybę sistemos savireguliacijai (homeostazei), o tai suteikia ekosistemai stabilumo: nuostabų įvairių elementų procento pastovumą. Čia galioja ekosistemų funkcionavimo principas: išteklių įsigijimas ir atliekų šalinimas vyksta visų elementų ciklo rėmuose. Naudotos literatūros sąrašas 1.Bigonas M.I. Ekologija, individai, populiacijos ir bendruomenės. M.: 1989. - 290 p. 2.Beysenova A.S., Shildebaev Zh.B., Sautbaeva G.Z. Ekologija. Almata: „Gylym, 2001. – 201 p. 3.Akimova T.A., Khaskin V.V. Ekologija. M.: Vienybės leidykla, 1998. - 233 p. .Gorelovas A.A. Ekologija. Paskaitų kursas. M.: "Centras" 1997. - 280 p. .Stadnitsky G.V., Rodionovas A.I. Ekologija. M.: Vienybės leidykla, 1996. - 272 p. .Sadanov A.K., Svanbaeva Z.S., Ekologija. Almata: "Agrouniversitetas", 1999. - 197 p. .Šilovas I.A. Ekologija. M.: Aukštoji mokykla, 2000. - 348 p. .Reimersas N.F. Ekologija (teorija, dėsniai, taisyklės, principai ir hipotezės). M.: "Jaunoji Rusija", 1994. - 260 p. .Tsvetkova L.I., Aleksejevas M.I., Usenovas B.P. ir kt.. Ekologija. Vadovėlis technikos universitetams. M.; ASV, Sankt Peterburgas: Khimizdat, 1999, 185 p. .Girusovas E.V. Aplinkos vadybos ekologija ir ekonomika. M.: Teisė ir teisė, VIENYBĖ. 1998 - 232 p. .Vronskis V.A. Taikomoji ekologija: Vadovėlis. Nauda. Rostovas prie Dono: 1995. - 197 p. .Budyko M.I. Pasaulinė ekologija. M.: Mysl, 1977.-248 p. .Alekseenko V.A. Aplinkos geochemija. Vadovėlis. M.: Logos, 2000 - 410 p. .Petrovas K.M. Bendroji S-P ekologija. Pamoka. "Chemija", 1997. - 218 p. .Andersenas J.M. Ekologija ir aplinkos mokslas. Biosfera, ekosistema, žmogus. L., 1985. 376 p. .Girenok F.I. Ekologija. Civilizacija. Noosfera.M.: 191987. - 281 p. .Ieškant pusiausvyros: ekologija socialinių politinių prioritetų sistemoje M.: 1992. - 427 p. .Visuomenės ir gamtos sąveikos istorija: veiksniai ir samprata. M.: ANSSSR 1990 m. .Stepanovskikh A.S. Taikomoji ekologija: Aplinkos apsauga. Vadovėlis universitetams. M.: VIENYBĖ DANA. 2003. 751 p. Panašūs darbai į - Medžiagų ir energijos srautų ciklas ekosistemose
Sąvokos „medžiagos srautas“ ir „energijos srautas“ turėtų būti aiškiai apibrėžtos. Medžiagos srautas yra judėjimas cheminių elementų ir jų junginių pavidalu nuo gamintojų iki skaidytojų (per vartotojus arba be jų). Energijos srautas – tai energijos perdavimas organinių junginių (maisto) cheminių ryšių pavidalu išilgai maisto grandinių iš vieno trofinio lygio į kitą (aukštesnį).
Pažymėtina, kad skirtingai nuo medžiagų, kurios nuolat cirkuliuoja per skirtingus ekosistemos blokus ir visada gali vėl patekti į ciklą, gaunama energija gali būti panaudota tik vieną kartą.
Kaip universalus gamtos reiškinys, vienpusis energijos srautas yra nulemtas termodinamikos dėsnių. Pagal pirmąjį dėsnį energija gali pereiti iš vienos formos (šviesos energija) į kitą (potencialią maisto energiją), tačiau ji niekada nesusikuria ir neišnyksta be pėdsakų.
Antrasis termodinamikos dėsnis teigia, kad negali būti vieno proceso, susieto su energijos transformacija, neprarandant jos dalies. Dėl šios priežasties negalima 100% efektyvumo paversti, pavyzdžiui, maisto medžiaga, iš kurios susideda organizmo kūnas.
Taigi visų ekosistemų funkcionavimą lemia nuolatinis energijos srautas, būtinas visiems organizmams savo egzistavimui ir savaiminiam dauginimuisi.
Ekosistemose taip pat egzistuoja konkurenciniai santykiai. Šiuo aspektu labai domina energijos maksimizavimo dėsnis (G. Odum - E. Odum): konkuruojant su kitomis ekosistemomis, išlieka ta, kuri geriausiai prisideda prie energijos tiekimo ir efektyviausiai panaudoja jos maksimalų kiekį. (konservai). Pagal įstatymą, šiam tikslui sistema: 1) sukuria kokybiškos energijos (pavyzdžiui, riebalų atsargų) kaupiklius (saugyklas); 2) išleidžia tam tikrą sukauptos energijos kiekį naujos energijos tiekimui užtikrinti; 3) užtikrina įvairių medžiagų cirkuliaciją; 4) sukuria reguliavimo mechanizmus, kurie palaiko sistemos stabilumą ir gebėjimą prisitaikyti prie kintančių sąlygų; 5) nustato mainus su kitomis sistemomis, reikalingomis specialių energijos rūšių poreikiui patenkinti.
Būtina pabrėžti svarbią aplinkybę: energijos maksimizavimo dėsnis galioja ir informacijos atžvilgiu, todėl (pagal N. F. Reimersą) jį taip pat galima laikyti energijos ir informacijos maksimizavimo dėsniu: sistema, kuri labiausiai prisideda prie to, kad 2010 m. energijos ir informacijos gavimas, gamyba ir efektyvus naudojimas.
Atkreipkime dėmesį į tai, kad didžiausias medžiagos tiekimas dar negarantuoja sistemos sėkmės konkurencinėje kitų panašių sistemų grupėje.
Anksčiau buvo pastebėta, kad tarp biocenozės organizmų atsiranda ir užsimezga stiprūs mitybos ryšiai arba maisto grandinė. Pastaroji susideda iš trijų pagrindinių grandžių: gamintojų, vartotojų ir skaidytojų.
Maisto grandinės, kurios prasideda fotosintetiniais organizmais, vadinamos ganymo (arba ganymo) grandinėmis, o grandinės, kurios prasideda nuo negyvų augalų medžiagų, skerdenų ir gyvūnų ekskrementų, vadinamos detritalinėmis grandinėmis.
Kiekvienos maisto grandinės grandies vieta vadinama trofiniu lygiu; jam būdingas skirtingas medžiagų ir energijos srauto intensyvumas. Pirmasis trofinis lygis visada yra gamintojai; antrasis – žolėdžiai vartotojai; trečiasis - mėsėdžiai, gyvenantys iš žolėdžių formų; ketvirtas lygis – kitų mėsėdžių valgymas ir kt.
Yra pirmos, antros, trečios ir ketvirtos eilės vartotojai, užimantys skirtingus maisto grandinės lygius (9 pav.).
Ryžiai. 9.
Akivaizdu, kad didelį vaidmenį čia vaidina vartotojų maisto produktų specializacija. Rūšys, turinčios platų mitybos spektrą, gali būti įtrauktos į skirtingų trofinių lygių maisto grandines. Pavyzdžiui, žmogaus mityba apima ir augalinį maistą, ir žolėdžių bei mėsėdžių mėsą. Todėl jis įvairiose maisto grandinėse veikia kaip I, II ar III užsakymų vartotojas.
Kadangi perkeliant energiją iš vieno lygio į kitą prarandama energija, galios grandinė negali būti ilga: dažniausiai ji susideda iš 4...6 grandžių (1 lentelė).
1. Tipinės mitybos grandinių schemos (pagal V. M. Ivonin, 1996)
Tačiau tokios grynos grandinės gamtoje paprastai nerandamos, nes tos pačios rūšys vienu metu gali būti skirtingose grandyse. Taip yra dėl to, kad gamtoje yra nedaug monofagų, daug dažniau pasitaiko oligofagų ir polifagų. Pavyzdžiui, plėšrūnai, mintantys įvairiais žolėdžiais ir mėsėdžiais gyvūnais, yra daugelio grandinių grandys. Dėl to kiekvienoje biocenozėje evoliuciškai susidaro maisto grandinių kompleksai, sudarantys vieną visumą. Taip kuriami elektros tinklai, kurie yra labai sudėtingi.
Taigi galime daryti išvadą, kad maisto grandinė yra pagrindinis energijos perdavimo kanalas bendruomenėje (tarp augalų – gamintojų, gyvūnų – vartotojų ir mikroorganizmų – skaidytojų) (10 pav.) Jau diagramoje aišku, kad idėja maisto grandinės ir trofiniai lygiai yra greičiau abstrakcija. Laboratorijoje galima sukurti linijinę grandinę su aiškiai atskirtais lygiais. Tačiau gamtoje tikrai yra trofinių tinklų, kuriuose daug populiacijų vienu metu priklauso keliems trofiniams lygiams. Tas pats organizmas sunaudoja tiek gyvūnai, tiek augalai; plėšrūnas gali maitintis pirmos ir antros eilės vartotojais; daugelis gyvūnų valgo ir gyvus, ir negyvus augalus.
Dėl trofinių santykių sudėtingumo vienos rūšies praradimas dažnai beveik neturi įtakos bendruomenei. Kiti „vartotojai“ pradeda vartoti išnykusių rūšių maistą,

Ryžiai. 10.
juo mintančios rūšys randa naujų maisto šaltinių: apskritai bendruomenėje išlaikoma pusiausvyra.
Gamintojų sugeriama energija teka maisto grandinėmis ir palaipsniui suvartojama. Maisto grandinės pabaigoje energijos kiekis visada yra mažesnis nei pradžioje. Fotosintezės metu augalai vidutiniškai suriša tik apie 1% saulės energijos, kuri patenka į juos. Gyvūnas, suėdęs augalą, dalies maisto nesuvirškina ir išskiria jį ekskrementų pavidalu. Paprastai suvirškinama 20...60% augalinio maisto; Sugerta energija naudojama gyvūno gyvybei palaikyti. Ląstelių ir organų funkcionavimą lydi šilumos išsiskyrimas, t. y. didelė maisto energijos dalis greitai išsisklaido aplinkoje. Palyginti nedidelė maisto dalis naudojama naujiems audiniams kurti ir riebalų atsargoms kurti. Toliau plėšrūnas, suėdęs žolėdį ir atstovaujantis trečiam trofiniam lygiui, gauna tik tą energiją iš augalo sukauptos energijos, kuri pasilieka grobio kūne (antrasis lygis) biomasės padidėjimo pavidalu.
Yra žinoma, kad kiekviename etape medžiagos ir energijos perdavimo maisto grandinėje metu prarandama apie 90% energijos ir tik apie dešimtadalį jos pereina kitam vartotojui, t.y. energijos perdavimas maisto jungtyse. organizmų paklūsta „dešimties procentų taisyklei“ (Lindemanno principas). Pavyzdžiui, tretinius mėsėdžius (penktą trofinį lygį) pasiekiančios energijos kiekis sudaro tik apie 10 -4 gamintojų sugeriamos energijos. Tai paaiškina ribotą (5...6) grandžių (lygių) skaičių maisto grandinėje, neatsižvelgiant į biocenozės rūšinės sudėties sudėtingumą.

Ryžiai. vienuolika.
Atsižvelgiant į energijos srautą ekosistemose, taip pat nesunku suprasti, kodėl biomasė mažėja didėjant trofiniam lygiui. Čia pasireiškia trečiasis pagrindinis ekosistemų funkcionavimo principas: kuo didesnė populiacijos biomasė, tuo žemesnį jos trofinį lygį turėtų užimti arba kitaip: ilgų mitybos grandinių gale negali būti didelės biomasės.
Aukščiau išvardinti trys pagrindiniai ekosistemos funkcionavimo principai – maistinių medžiagų ciklas, saulės energijos srautas ir biomasės mažėjimas didėjant trofiniam lygiui – gali būti pateikiami bendros diagramos pavidalu (11 pav.). Jei organizmus išdėstysime pagal jų mitybos santykius, kiekvienam iš jų nurodydami energijos ir maistinių medžiagų „įvedimą“ ir „išvedimą“, paaiškėja, kad ekosistemoje maistinės medžiagos yra nuolat perdirbamos, o per ją teka energijos srautas.
Organizmų gyvybinės veiklos ir medžiagų cirkuliacijos ekosistemose palaikymas, t.y., ekosistemų egzistavimas, priklauso nuo nuolatinio energijos srauto, reikalingo visiems organizmams jų gyvybinėms funkcijoms ir savaiminiam dauginimuisi.
Skirtingai nuo medžiagų, kurios nuolat cirkuliuoja per skirtingus ekosistemos blokus, kurias visada galima pakartotinai panaudoti ir patekti į ciklą, energija gali būti naudojama tik vieną kartą, t.y., ekosistemoje vyksta linijinis energijos srautas.
Vienpusis energijos antplūdis, kaip universalus gamtos reiškinys, atsiranda dėl termodinamikos dėsnių. Pirmasis įstatymas teigia, kad energiją galima paversti iš vienos formos (pavyzdžiui, šviesa) į kitą (pavyzdžiui, potencialią maisto energiją), tačiau jos negalima sukurti ar sunaikinti. Antrasis įstatymas teigia, kad negali būti vieno proceso, susieto su energijos transformacija, neprarandant dalies jos. Tam tikras energijos kiekis tokiose transformacijose išsisklaido į neprieinamą šiluminę energiją, todėl prarandama. Vadinasi, negali būti 100 procentų efektyvumo vykstančių, pavyzdžiui, maistinių medžiagų transformacijų į organizmo kūną sudarančią medžiagą.
Taigi gyvi organizmai yra energijos keitikliai. Ir kiekvieną kartą, kai energija virsta, dalis jos prarandama šilumos pavidalu. Galiausiai visa energija, patenkanti į biotinį ekosistemos ciklą, išsisklaido kaip šiluma. Gyvi organizmai iš tikrųjų nenaudoja šilumos kaip energijos šaltinio darbui atlikti – jie naudoja šviesą ir cheminę energiją.
Ekologinės piramidės. Kiekvienoje ekosistemoje maisto tinklai turi aiškiai apibrėžtą struktūrą, kuriai būdinga organizmų, atstovaujamų kiekviename įvairių maisto grandinių lygmenyje, pobūdžiu ir skaičiumi. Norėdami ištirti organizmų ryšius ekosistemoje ir pavaizduoti juos grafiškai, jie dažniausiai naudoja ekologines piramides, o ne maisto tinklo diagramas. Ekologinės piramidės geometrine forma išreiškia ekosistemos trofinę struktūrą. Jie sukonstruoti tokio paties pločio stačiakampių pavidalu, tačiau stačiakampių ilgis turi būti proporcingas matuojamo objekto vertei. Iš čia galite gauti skaičių, biomasės ir energijos piramidės.
Ekologinės piramidės atspindi pagrindines bet kokios biocenozės savybes, kai parodo jos trofinę struktūrą:
Jų aukštis yra proporcingas atitinkamos maisto grandinės ilgiui, t.y., joje esančių trofinių lygių skaičiui;
Jų forma daugiau ar mažiau atspindi energijos transformacijų efektyvumą pereinant iš vieno lygio į kitą.
Skaičių piramidės. Jie yra paprasčiausias ekosistemos trofinės struktūros tyrimo priartėjimas. Tokiu atveju pirmiausia suskaičiuojamas organizmų skaičius tam tikroje teritorijoje, sugrupuojamas pagal trofinius lygius ir pateikiamas stačiakampio pavidalu, kurio ilgis (arba plotas) yra proporcingas tam tikroje teritorijoje gyvenančių organizmų skaičiui ( arba tam tikrame tūryje, jei tai vandens ekosistema). Nustatyta pagrindinė taisyklė, kad bet kurioje aplinkoje yra daugiau augalų nei gyvūnų, daugiau žolėdžių nei mėsėdžių, daugiau vabzdžių nei paukščių ir pan.
Populiacijos piramidės atspindi organizmų tankį kiekviename trofiniame lygmenyje. Įvairių gyventojų piramidžių statybos yra labai įvairios. Dažnai jie būna apversti.
Pavyzdžiui, miške medžių (pirminių gamintojų) yra žymiai mažiau nei vabzdžių (žolėdžių).
Biomasės piramidė. Tiksliau atspindi maisto santykius ekosistemoje, nes atsižvelgiama į bendrą organizmų masę (biomasė) kiekvienas trofinis lygis. Stačiakampiai biomasės piramidėse rodo organizmų masę kiekviename trofiniame lygyje ploto arba tūrio vienetui. Biomasės piramidės forma dažnai yra panaši į gyventojų piramidės formą. Būdingas biomasės sumažėjimas kiekviename trofiniame lygyje.
Biomasės piramidės, kaip ir skaičiai, gali būti ne tik tiesios, bet ir apverstos. Apverstos biomasės piramidės būdingos vandens ekosistemoms, kuriose pirminiai gamintojai, pavyzdžiui, fitoplanktoniniai dumbliai, labai greitai dalijasi, o jų vartotojai – zooplanktoniniai vėžiagyviai – yra daug didesni, tačiau turi ilgą dauginimosi ciklą. Tai ypač pasakytina apie gėlo vandens aplinką, kur pirminį produktyvumą užtikrina mikroskopiniai organizmai, kurių medžiagų apykaitos greitis yra padidėjęs, ty biomasė maža, produktyvumas didelis.
Energijos piramidė. Svarbiausias būdas parodyti ryšius tarp skirtingų trofinių lygių organizmų yra energijos piramidės. Jie atspindi maisto grandinių energijos konversijos efektyvumą ir produktyvumą ir yra sudaryti skaičiuojant energijos kiekį (kcal), sukauptą paviršiaus vienetui per laiko vienetą ir sunaudojamą organizmų kiekviename trofiniame lygmenyje. Taigi, palyginti nesunku nustatyti biomasėje sukauptos energijos kiekį, tačiau sunkiau įvertinti bendrą energijos kiekį, sugertą kiekviename trofiniame lygmenyje. Sukūrę grafiką (12.28 pav.), galime teigti, kad destruktoriai, kurių svarba biomasės piramidėje atrodo maža, o gyventojų piramidėje – atvirkščiai; gauna didelę dalį energijos, einančios per ekosistemą. Be to, tik dalis visos šios energijos lieka organizmuose kiekviename trofiniame ekosistemos lygyje ir kaupiasi biomasėje, o likusi dalis naudojama gyvų būtybių medžiagų apykaitos poreikiams tenkinti: egzistavimui, augimui, dauginimuisi. Gyvūnai taip pat nemažai energijos išeikvoja raumenų darbui.
R. Lindemanas pirmą kartą suformulavo 1942 m energijos piramidės dėsnis, kuris vadovėliuose dažnai vadinamas „10% įstatymu“. Pagal šį įstatymą iš vieno ekologinės piramidės trofinis lygis Vidutiniškai ne daugiau kaip 10% energijos pereina į kitą lygį.
Vartotojai yra valdymo ir stabilizavimo grandis ekosistemoje. Vartotojai sukuria cenozės įvairovės spektrą, užkertant kelią dominuojančių asmenų monopolijai. Vartotojų vertės kontrolės taisyklė pagrįstai gali būti laikomas gana esminiu. Pagal kibernetinį požiūrį, valdymo sistema turėtų būti sudėtingesnės struktūros nei valdoma, tada paaiškėja vartotojų tipų įvairovės priežastis. Kontroliuojanti vartotojų reikšmė turi ir energetinį pagrindą. Energijos srauto per vieną ar kitą trofinį lygį negali visiškai nulemti maisto prieinamumas pagrindiniame trofiniame lygyje. Kaip žinoma, visada lieka pakankamai „rezervo“, nes visiškas maisto sunaikinimas lemtų vartotojų mirtį. Šie bendrieji modeliai stebimi populiacijos procesų, bendruomenių, ekologinės piramidės lygių ir visos biocenozių kontekste.
Norėdami naudoti pristatymų peržiūras, susikurkite „Google“ paskyrą ir prisijunkite prie jos: https://accounts.google.com
Skaidrių antraštės:
Materijos ir energijos ciklas gamtoje
Medžiagų ciklas – tai pasikartojantys medžiagos virsmo ir judėjimo procesai gamtoje, kurie yra daugiau ar mažiau cikliški. Visos mūsų planetoje esančios medžiagos yra cirkuliacijos procese. Gamtoje yra du pagrindiniai ciklai: didelis (geologinis) mažas (biogeocheminis)
Didysis medžiagų ciklas Didysis ciklas trunka milijonus metų ir jį sukelia saulės energijos sąveika su gilia Žemės energija. Susijęs su geologiniais procesais, uolienų susidarymu ir sunaikinimu bei vėlesniu naikinimo produktų judėjimu.
Mažas medžiagų ciklas Mažas ciklas (biogeocheminis) vyksta biosferoje, biocenozės lygmenyje. Jo esmė – fotosintezės metu iš neorganinių junginių susidaro gyvosios medžiagos ir skaidant organines medžiagas vėl virsta neorganiniais junginiais. Biogeocheminiai ciklai – Vernadskis V.I.
Vandens ciklas Tr nuotėkio inf Vandens garavimas Garų kondensacija Kritulių nuotėkis Transpiracija infiltracija
Transpiracija – tai vandens judėjimas augalu ir jo išgarinimas per išorinius augalo organus, tokius kaip lapai, stiebai ir žiedai. Vanduo yra būtinas augalų gyvybei, tačiau tik nedidelė dalis vandens, tiekiamo per šaknis, yra tiesiogiai naudojama augimui ir medžiagų apykaitai.
Vandens ciklas
Vandens ciklas Didžioji dalis vandens telkiasi vandenynuose. Iš jų paviršiaus garuojantis vanduo aprūpina natūralias ir dirbtines žemės ekosistemas. Kuo vietovė arčiau vandenyno, tuo daugiau kritulių. Žemė nuolat grąžina vandenį į vandenyną: dalis drėgmės išgaruoja, aktyviausiai miškuose, dalį surenka upės: jos sulaukia lietaus ir tirpsmo vandens. Drėgmės mainai tarp vandenyno ir sausumos reikalauja labai didelių energijos sąnaudų: tam išleidžiama apie 30% Žemę pasiekiančios saulės energijos.
Žmogaus įtaka vandens ciklui Vandens ciklas biosferoje iki civilizacijos vystymosi buvo pusiausvyros, t.y. vandenynas iš upių gavo tiek vandens, kiek suvartojo išgaruodamas. Vystantis civilizacijai, šis ciklas ėmė trūkinėti. Visų pirma, miškai išgarina vis mažiau vandens, nes... jų plotas mažėja, o dirvos paviršius, atvirkščiai, didėja, nes Didėja drėkinamos žemės ūkio paskirties žemės plotai. žemes. Pietinių regionų upės tapo seklios. Vanduo blogiau išgaruoja nuo vandenyno paviršiaus, nes... nemaža jo dalis padengta aliejaus plėvele. Visa tai blogina vandens tiekimą į biosferą.
Dažnėja sausros, atsiranda ekologinių nelaimių kišenės. Pavyzdžiui, Afrikoje, Sahelio zonoje – pusiau dykumos regione, skiriančiame Sacharą nuo šiaurinių žemyno šalių, katastrofiška sausra tęsiasi daugiau nei 35 metus. Iš sausumos į vandenyną ir kitus vandens telkinius grįžtantis gėlas vanduo dažnai būna užterštas. Daugelio Rusijos upių vanduo tapo praktiškai netinkamas gerti. Gyviems organizmams prieinamo gėlo vandens dalis yra gana maža, todėl jį reikia naudoti taupiai ir neužteršto! Kas ketvirtam planetos gyventojui trūksta švaraus geriamojo vandens. Daugelyje pasaulio vietų nepakanka vandens pramoninei gamybai ir drėkinimui.
Skirtingi hidrosferos komponentai vandens cikle dalyvauja skirtingais būdais ir skirtingu greičiu. Visiškam vandens atsinaujinimui ledynuose reikia 8000 metų, požeminiam – 5000 metų, vandenynui – 3000 metų, dirvožemiui – 1 metai. Atmosferos garai ir upių vandenys visiškai atsinaujina per 10–12 dienų. Vandens ciklas gamtoje trunka apie 1 milijoną metų.
Deguonies ciklas Deguonis yra labiausiai paplitusių biosferos elementų dalis. Deguonies kiekis atmosferoje yra beveik 21%. Deguonis yra vandens molekulių ir gyvų organizmų (baltymų, riebalų, angliavandenių, nukleino rūgščių) dalis. Deguonį gamina gamintojai (žali augalai). Ozonas vaidina svarbų vaidmenį deguonies cikle. Ozono sluoksnis yra 20-30 km aukštyje virš jūros lygio. Deguonies kiekį atmosferoje įtakoja 2 pagrindiniai procesai: 1) fotosintezė 2) organinių medžiagų skilimas, kurio metu jos suvartojamos.
Deguonies ciklas yra lėtas procesas. Kad atmosferoje visiškai atsinaujintų visas deguonis, prireikia maždaug 2000 metų. Palyginimui: visiškas anglies dioksido atsinaujinimas atmosferoje įvyksta maždaug per 3 metus. Deguonį kvėpavimui naudoja dauguma gyvų organizmų. Deguonis naudojamas deginant degalus vidaus degimo varikliuose, šiluminių elektrinių krosnyse, orlaivių ir raketų varikliuose ir kt. Papildomas antropogeninis vartojimas gali sutrikdyti deguonies ciklo pusiausvyrą. Kol kas biosfera kompensuoja žmogaus įsikišimą: nuostolius kompensuoja žali augalai. Toliau mažėjant miškų plotams ir deginant vis daugiau kuro, deguonies kiekis atmosferoje pradės mažėti.
SVARBU!!! Sumažėjus deguonies kiekiui ore iki 16%, pablogėja žmogaus sveikata (ypač kenčia širdis), iki 7%, žmogus netenka sąmonės, o iki 3% miršta.
Anglies ciklas
Anglies ciklas Anglis yra organinių junginių pagrindas, ji yra visų gyvų organizmų dalis baltymų, riebalų ir angliavandenių pavidalu. Anglis į atmosferą patenka anglies dioksido pavidalu. Atmosferoje, kurioje koncentruojasi didžioji anglies dioksido dalis, nuolat vyksta mainai: augalai fotosintezės metu pasisavina anglies dvideginį, o kvėpuodami jį išskiria visi organizmai. Iki 50% anglies CO 2 pavidalu į atmosferą grąžina skaidytojai – dirvožemio mikroorganizmai. Anglis palieka ciklą kaip kalcio karbonatas.
Žmogaus įtaka anglies ciklui Technogeninė žmogaus veikla sutrikdo natūralią anglies ciklo pusiausvyrą: 1) degant organiniam kurui į atmosferą kasmet išmetama apie 6 milijardus tonų CO 2: a) Elektros gamyba šiluminėse elektrinėse b) ) Automobilių išmetamosios dujos 2) miškų naikinimas. Per pastaruosius 100 metų anglies dioksido lygis atmosferoje nuolat ir sparčiai augo. Anglies dioksidas + metanas + vandens garai + ozonas + azoto oksidai = šiltnamio efektą sukeliančios dujos. Rezultatas yra šiltnamio efektas – visuotinis atšilimas, dėl kurio gali kilti didelio masto stichinės nelaimės.
Azoto ciklas Laisvoje formoje azotas yra oro komponentas – 78%. Azotas yra vienas iš svarbiausių elementų organizmų gyvybei. Azotas yra visų baltymų dalis. Azoto molekulė yra labai stipri, todėl dauguma organizmų nepajėgia absorbuoti atmosferos azoto. Gyvi organizmai azotą pasisavina tik junginių su vandeniliu ir deguonimi pavidalu. Azoto fiksavimas į cheminius junginius vyksta dėl vulkaninės ir perkūnijos veiklos, bet dažniausiai dėl mikroorganizmų – azoto fiksatorių (azotą fiksuojančių bakterijų ir melsvadumblių) veiklos.
Azotas į augalų šaknis patenka nitratų pavidalu, kurie naudojami organinių medžiagų (baltymų) sintezei. Gyvūnai azotą suvartoja per augalinį ar gyvūninį maistą. Azotas grąžinamas į atmosferą skaidant negyvas organines medžiagas. Dirvožemio bakterijos baltymus skaido į neorganines medžiagas – dujas – amoniaką, azoto oksidus, kurie patenka į atmosferą. Azotas, patekęs į vandens telkinius, taip pat pereina augalų-gyvūnų-mikroorganizmų mitybos grandinę ir grįžta į atmosferą.
Žmogaus įtaka azoto ratui Technogeninė žmogaus veikla sutrikdo natūralią azoto ciklo pusiausvyrą. Ariant žemę beveik 5 kartus sumažėja mikroorganizmų – azoto fiksatorių aktyvumas, todėl dirvoje mažėja azoto kiekis, dėl to mažėja dirvožemio derlingumas. Todėl žmonės į dirvą įveda nitratų perteklių, kurie yra mineralinėse trąšose. Didelis azoto oksidų kiekis patenka į atmosferą degant ir apdorojant dujas, naftą ir anglį ir patenka rūgščiojo lietaus pavidalu. Atkurti natūralų azoto ciklą galima sumažinus azoto trąšų gamybą, mažinant pramonines azoto oksidų emisijas į atmosferą ir kt.
Fosforo ciklas
Skirtingai nuo vandens, anglies, azoto ir deguonies ciklų, kurie yra uždari, fosforo ciklas yra atviras, nes fosforas nesudaro lakiųjų junginių, patenkančių į atmosferą. Fosforo yra uolienose, iš kurių jis patenka į ekosistemas natūralaus uolienų naikinimo metu arba įterpiant fosforo trąšas laukuose. Augalai pasisavina neorganinius fosforo junginius, o šiais augalais mintantys gyvūnai fosforą kaupia savo audiniuose. Skilus gyvūnų ir augalų kūnams, ne visas fosforas dalyvauja cikle. Dalis jo išplaunama iš dirvožemio į vandens telkinius (upes, ežerus, jūras) ir nusėda į dugną. Fosforas nedideliais kiekiais grįžta į sausumą su žmonių sugauta žuvimi.
Žmogaus įtaka fosforo ciklui Fosforo pernešimas iš sausumos į vandenyną dėl žmogaus įtakos labai išaugo. Naikinant miškus ir ariant dirvas, padidėja paviršinio vandens tėkmės tūris, be to, į upes ir ežerus iš laukų patenka fosforo trąšos. Kadangi fosforo atsargos sausumoje yra ribotos, o jo grąžinimas iš vandenyno yra sunkus, ateityje žemės ūkyje gali trūkti fosforo, dėl to sumažės derlius (pirmiausia grūdinių kultūrų).