Fluxo de energia na natureza. Ciclo de matéria e energia na natureza
Qualquer vida requer um fluxo constante de energia e matéria. A energia é gasta na realização de reações básicas da vida, a matéria é usada para construir os corpos dos organismos. A existência de ecossistemas naturais é acompanhada por processos complexos de troca material e energética entre a natureza viva e a inanimada. Estes processos são muito importantes e dependem não só da composição das comunidades bióticas, mas também do seu ambiente físico.
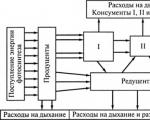
Fluxo de energia na comunidade– esta é a sua transição de organismos de um nível para outro na forma de ligações químicas de compostos orgânicos (alimentos).
O fluxo (ciclo) da matéria é o movimento da matéria na forma de elementos químicos e seus compostos dos produtores para os decompositores e depois (por meio de reações químicas que ocorrem sem a participação de organismos vivos) novamente para os produtores.
A circulação da matéria e o fluxo de energia não são conceitos idênticos, embora vários equivalentes de energia (calorias, quilocalorias, joules) sejam frequentemente utilizados para medir o movimento da matéria. Isso se explica em parte pelo fato de que em todos os níveis tróficos, com exceção do primeiro, a energia necessária à vida dos organismos é transferida na forma da substância dos alimentos consumidos. Somente as plantas (produtoras) podem utilizar diretamente a energia radiante do Sol para sua atividade vital.
Uma medição rigorosa de uma substância que circula em um ecossistema pode ser obtida levando-se em consideração a circulação de elementos químicos individuais, principalmente aqueles que são o principal material de construção do citoplasma das células vegetais e animais.
Ao contrário das substâncias que circulam continuamente através de diferentes blocos do ecossistema e podem sempre reentrar no ciclo, a energia pode ser utilizada no corpo apenas uma vez.
De acordo com as leis da física, a energia pode mudar de uma forma (como a energia luminosa) para outra (como a energia potencial dos alimentos), mas nunca é criada novamente ou destruída. Não pode haver um único processo associado à transformação de energia sem perder parte dela. Em suas transformações, certa quantidade de energia é dissipada em forma de calor e, portanto, perdida. Por isso, não pode haver transformações, por exemplo, de substâncias alimentares nas substâncias que compõem o corpo do organismo, que ocorrem com cem por cento de eficiência.
A existência de todos os ecossistemas depende de um fluxo constante de energia, necessário para que todos os organismos mantenham as suas funções vitais e a sua auto-reprodução.
Apenas cerca de metade do fluxo solar que incide sobre as plantas verdes é absorvido pelos elementos fotossintéticos, e apenas uma pequena fração da energia absorvida (de 1/100 a 1/20 da parte) é armazenada na forma de energia necessária para a atividade. dos tecidos vegetais.
À medida que nos afastamos do produtor primário, a taxa de fluxo de energia (isto é, a quantidade de energia, expressa em unidades de energia, transferida de um nível trófico para outro) diminui acentuadamente.
A queda na quantidade de energia durante a transição de um nível trófico para um superior determina o próprio número desses níveis. Estima-se que qualquer nível trófico receba apenas cerca de 10% (ou um pouco mais) da energia do nível anterior. Portanto, o número total de níveis tróficos raramente excede 3–4.
A proporção de matéria viva em diferentes níveis tróficos geralmente obedece à mesma regra que a proporção de energia recebida: quanto maior o nível, menor a biomassa total e o número de seus organismos constituintes.
A proporção entre o número de diferentes grupos de organismos dá uma ideia da estabilidade da comunidade, porque a biomassa e o número de algumas populações são ao mesmo tempo um indicador do espaço vital para organismos desta e de outras espécies . Por exemplo, o número de árvores numa floresta determina não só o fornecimento total de biomassa e energia nelas contidas, mas também o microclima, bem como o número de abrigos para muitos insetos e pássaros.
Pirâmides de números podem ser invertidas. Isto ocorre quando a taxa de reprodução da população de presas é alta, e mesmo com baixa biomassa, tal população pode ser uma fonte de alimento suficiente para predadores que têm uma biomassa mais elevada, mas uma baixa taxa de reprodução. Por exemplo, muitos insetos podem viver e se alimentar de uma árvore (uma pirâmide populacional invertida). A pirâmide invertida da biomassa é característica dos ecossistemas aquáticos, onde os produtores primários (algas fitoplanctônicas) se dividem e se multiplicam muito rapidamente em número, e seus consumidores (crustáceos zooplanctônicos) são muito maiores, mas têm um longo ciclo de reprodução.
Pastagens e cadeias detríticas
A energia pode fluir através de uma comunidade de muitas maneiras diferentes. Representa a cadeia alimentar de todos os consumidores(sistema consumidor) com a adição de mais dois links: este matéria orgânica morta E cadeia alimentar do decompositor(sistema redutor).
Fluxo de energia das plantas através dos herbívoros(eles são chamados de pastoreio), chamada cadeia alimentar das pastagens.
Os restos dos organismos que consomem e que não são utilizados pelos consumidores reabastecem a matéria orgânica morta. É composto por fezes contendo parte de alimentos não digeridos, além de cadáveres de animais, resíduos vegetais (folhas, galhos, algas) e é denominado detritos.
O fluxo de energia proveniente da matéria orgânica morta e que passa pelo sistema em decomposição é chamado de cadeia alimentar detrítica.
Juntamente com as semelhanças, existem diferenças profundas no funcionamento das pastagens e das cadeias alimentares detríticas. Consiste no fato de que em no sistema de consumo, perdem-se fezes e organismos mortos, e no sistema redutivo– Não.
Mais cedo ou mais tarde, a energia contida na matéria orgânica morta será totalmente utilizada pelos decompositores e dissipada como calor através da respiração, mesmo que isso exija passar várias vezes por um sistema de decompositores. As únicas exceções são aqueles casos em que as condições abióticas locais são muito desfavoráveis ao processo de decomposição (alta umidade, permafrost). Nestes casos, acumulam-se depósitos de substâncias incompletamente processadas e com alto consumo de energia, que se transformam ao longo do tempo e sob condições adequadas em fósseis orgânicos combustíveis - petróleo, carvão, turfa.
Ciclo de substâncias em um ecossistema
A integridade dos ecossistemas naturais se manifesta de maneira especialmente clara quando se consideram os fluxos de matéria que neles circulam. A substância pode ser transmitida em ciclos fechados (circulações), circulando repetidamente entre os organismos e o meio ambiente.
Os movimentos circulares (através da terra, do ar, da água) de elementos químicos (isto é, substâncias) são chamados de ciclos ou circulações biogeoquímicas.
Os elementos e sais dissolvidos necessários à vida são convencionalmente chamados nutrientes(dando vida) ou nutrientes. Entre os elementos biogênicos, distinguem-se dois grupos: substâncias macrotróficas e substâncias microtróficas.
Substâncias macrotróficas abrangem os elementos que constituem a base química dos tecidos dos organismos vivos. Estes incluem: carbono, hidrogênio, oxigênio, nitrogênio, fósforo, potássio, cálcio, magnésio, enxofre.
Substâncias microtróficas incluem elementos e seus compostos, também muito importantes para a existência dos sistemas vivos, mas em quantidades extremamente pequenas. Tais substâncias são frequentemente chamadas microelementos. São eles ferro, manganês, cobre, zinco, boro, sódio, molibdênio, cloro, vanádio e cobalto. Embora os elementos microtróficos sejam essenciais para os organismos em quantidades muito pequenas, a sua deficiência pode limitar severamente a produtividade.
A circulação dos nutrientes costuma ser acompanhada por suas transformações químicas. O nitrogênio nitrato, por exemplo, pode ser convertido em nitrogênio protéico, depois convertido em uréia, convertido em amônia e novamente sintetizado na forma de nitrato sob a influência de microrganismos. Vários mecanismos, tanto biológicos quanto químicos, estão envolvidos nos processos de desnitrificação e fixação de nitrogênio.
As reservas de nutrientes são variáveis. O processo de sequestrar alguns deles como biomassa viva reduz a quantidade restante no ambiente abiótico. E se as plantas e outros organismos não se decompusessem, o fornecimento de nutrientes esgotar-se-ia e a vida na Terra cessaria. Disto podemos concluir que a atividade de heterótrofos, principalmente organismos que funcionam em cadeias detríticas,– fator decisivo na manutenção do ciclo dos nutrientes e na formação dos produtos.
Vejamos alguns dados numéricos que indicam a escala da transferência de substâncias, voltando-nos para o ciclo biogeoquímico do carbono. A fonte natural de carbono utilizada pelas plantas para a síntese da matéria orgânica é o dióxido de carbono, que faz parte da atmosfera ou está dissolvido na água. Durante a fotossíntese, o dióxido de carbono (dióxido de carbono) é convertido em matéria orgânica que serve de alimento para os animais. A respiração, a fermentação e a combustão de combustíveis devolvem dióxido de carbono à atmosfera.
As reservas de carbono na atmosfera do nosso planeta são estimadas em 700 bilhões de toneladas, na hidrosfera - em 50 bilhões de toneladas.De acordo com cálculos anuais, como resultado da fotossíntese, o aumento da massa vegetal na terra e na água é de 30 bilhões de toneladas e 150 bilhões de toneladas, respectivamente. O ciclo do carbono continua por cerca de 300 anos.
Outro exemplo é o ciclo do fósforo. As principais reservas de fósforo contêm diversas rochas, que gradativamente (como resultado da destruição e da erosão) liberam seus fosfatos para os ecossistemas terrestres. Os fosfatos são consumidos pelas plantas e utilizados por elas para sintetizar substâncias orgânicas. Quando as carcaças dos animais são decompostas por microrganismos, os fosfatos são devolvidos ao solo e depois utilizados novamente pelas plantas. Além disso, parte dos fosfatos é transportada por cursos d'água para o mar. Isto garante o desenvolvimento do fitoplâncton e de todas as cadeias alimentares dele dependentes. Parte do fósforo contido na água do mar pode retornar à terra na forma de guano.
ciclo de energia da biosfera do ecossistema
Introdução
Conceito e estrutura dos ecossistemas
1 Conceito de ecossistemas
2 Classificação dos ecossistemas
3 Zoneamento de macroecossistemas
4 Estrutura do ecossistema
Fatores que garantem a integridade dos ecossistemas
1 Ciclos de substâncias
2 Fluxo de energia nos ecossistemas
3 Processos dinâmicos que garantem integridade e sustentabilidade nos ecossistemas
4 A biosfera como ecossistema global que garante a integridade e a sustentabilidade dos ecossistemas
Conclusão
Introdução
Um ecossistema é qualquer conjunto de organismos e componentes inorgânicos nos quais pode ocorrer a circulação de substâncias. De acordo com N.F. Reimers (1990), um ecossistema é qualquer comunidade de seres vivos e seu habitat, unidos em um único todo funcional, surgindo com base na interdependência e nas relações de causa e efeito que existem entre os componentes ambientais individuais. A. Tansley (1935) propôs a seguinte relação:
Existem microecossistemas, mesoecossistemas, macroecossistemas e a biosfera global. Grandes ecossistemas terrestres são chamados de biomas. Os ecossistemas não estão dispersos desordenadamente; pelo contrário, estão agrupados em zonas bastante regulares, tanto horizontalmente (em latitude) como verticalmente (em altura).
Do equador aos pólos, uma certa simetria é visível na distribuição dos biomas dos diferentes hemisférios: florestas tropicais, desertos, estepes, florestas temperadas, florestas de coníferas, taiga.
Todo ecossistema tem dois componentes principais: organismos e fatores de seu ambiente inanimado. A totalidade dos organismos (plantas, animais, micróbios) é chamada de biota de um ecossistema. Do ponto de vista da estrutura trófica (do grego trophe - nutrição), o ecossistema pode ser dividido em duas camadas: superior e inferior.
Os seguintes componentes são diferenciados como parte de um ecossistema: compostos inorgânicos, orgânicos, ambientes de ar, água e substrato, produtores, organismos autotróficos, consumidores ou fagotróficos, decompositores e detritívoros.
A energia solar na Terra causa dois ciclos de substâncias: grandes, ou geológicas, e pequenas, biológicas (bióticas).
Num ecossistema existem ciclos de substâncias. Os mais estudados são: os ciclos do carbono, oxigênio, nitrogênio, fósforo, enxofre, etc.
A manutenção da atividade vital dos organismos e da circulação da matéria nos ecossistemas, ou seja, a existência dos ecossistemas, depende do fluxo constante de energia solar.
Nos ecossistemas, ocorrem constantemente mudanças no estado e na atividade vital dos seus membros e na proporção das populações. As diversas mudanças que ocorrem em qualquer comunidade dividem-se em dois tipos principais: cíclicas e progressivas.
Um ecossistema global é uma biosfera caracterizada pela integridade e estabilidade dos ecossistemas.
1. Conceito e estrutura dos ecossistemas
1 Conceito de ecossistemas
Os organismos vivos e seu ambiente inanimado (abiótico) estão inseparavelmente conectados entre si e em constante interação. Qualquer unidade (biossistema) de um ecossistema é um sistema ecológico. Um sistema ecológico, ou ecossistema, é a unidade funcional básica em ecologia, pois inclui os organismos e o ambiente inanimado - componentes que influenciam mutuamente as propriedades uns dos outros e as condições necessárias para manter a vida na forma que existe na Terra. O termo "ecossistema" foi proposto pela primeira vez em 1935 pelo ecologista inglês A. Tansley. Atualmente, a seguinte definição de ecossistema é amplamente utilizada. Um ecossistema é qualquer conjunto de organismos e componentes inorgânicos nos quais pode ocorrer a circulação de substâncias. De acordo com N.F. Reimers (1990), um ecossistema é qualquer comunidade de seres vivos e seu habitat, unidos em um único todo funcional, surgindo com base na interdependência e nas relações de causa e efeito que existem entre os componentes ambientais individuais. Deve-se enfatizar que a combinação de um ambiente físico-químico específico (biótopo) com uma comunidade de organismos vivos (biocenose) forma um ecossistema. A. Tansley (1935) propôs a seguinte relação:
Ecossistema = Biótopo + Biocenose
De acordo com a definição de V.N. Sukachev, a biogeocenose é “um conjunto de fenômenos naturais homogêneos (atmosfera, rocha, solo e condições hidrológicas) sobre uma certa extensão da superfície terrestre, que possui uma especificidade especial própria de interações desses componentes que o compõem e um certo tipo de troca de matéria e energia entre si e outros fenômenos naturais e representando uma unidade dialética internamente contraditória, em constante movimento e desenvolvimento."
Além dos conceitos de ecossistema A. Tensley e biogeocenose V.N. Sukachev formulou a regra de F. Evans (1956), que propôs usar o termo “ecossistema” de forma absolutamente “adimensional” para designar qualquer sistema vivo supraorganismo interagindo com o meio ambiente. No entanto, muitos autores deram ao termo “ecossistema” o significado de biogeocenose, ou seja, ecossistema elementar e, ao mesmo tempo, superior na hierarquia das formações suprabiogeocenóticas até o ecossistema da biosfera.
2 Classificação dos ecossistemas
Os ecossistemas existentes na Terra são diversos. Existem microecossistemas (por exemplo, o tronco de uma árvore podre), mesoecossistemas (floresta, lagoa, etc.), macroecossistemas (continente, oceano, etc.) e a biosfera global.
Grandes ecossistemas terrestres são chamados de biomas. Cada bioma contém vários ecossistemas menores e interconectados. Existem várias classificações de ecossistemas:
Floresta tropical perene
Deserto: Grama e arbusto Chaparral - áreas com invernos chuvosos e verões secos
Graslenz tropical e savana
Estepe temperada
Floresta decídua temperada
Florestas boreais de coníferas
Tundra: ártica e alpina
Tipos de ecossistemas de água doce:
Fita (águas paradas): lagos, lagoas, etc.
Lótico (águas correntes): rios, riachos, etc.
Zonas úmidas: pântanos e florestas pantanosas
Tipos de ecossistemas marinhos
Oceano aberto (pelágico)
Águas da plataforma continental (águas costeiras)
Áreas de ressurgência (áreas férteis com pesca produtiva)
Estuários (baías costeiras, estreitos, fozes de rios, sapais, etc.). Os biomas terrestres são diferenciados aqui pelas características da vegetação natural ou original, e os tipos de ecossistemas aquáticos são diferenciados pelas características geológicas e físicas. Os principais tipos de ecossistemas listados representam o ambiente em que a civilização humana se desenvolveu e representam as principais comunidades bióticas que sustentam a vida na Terra.
3 Zoneamento de macroecossistemas
O estudo da distribuição geográfica dos ecossistemas só pode ser realizado ao nível das grandes unidades ecológicas - macroecossistemas, que são considerados à escala continental. Os ecossistemas não estão dispersos desordenadamente; pelo contrário, estão agrupados em zonas bastante regulares, tanto horizontalmente (em latitude) como verticalmente (em altura). Isto é confirmado pela lei periódica de zoneamento geográfico de A.A. Grigoriev - M.I. Budyko: com a mudança das zonas físico-geográficas da Terra, zonas de paisagem semelhantes e algumas de suas propriedades gerais se repetem periodicamente. Isso também foi discutido ao considerar o ambiente terrestre da vida. A periodicidade estabelecida por lei manifesta-se no facto de os valores do índice de secura variarem nas diferentes zonas de 0 a 4-5, três vezes entre os pólos e o equador estão próximos da unidade. Estes valores correspondem à maior produtividade biológica das paisagens.
A repetição periódica de propriedades em uma série de sistemas do mesmo nível hierárquico é provavelmente uma lei geral do universo, formulada como a lei da periodicidade na estrutura dos agregados sistêmicos, ou uma lei sistema-periódica - sistemas naturais específicos do mesmo nível ( subnível) de organização constituem uma série periódica ou repetida de estruturas morfologicamente semelhantes dentro dos limites espaço-temporais superiores e inferiores do sistema, além das quais a existência de sistemas em um determinado nível se torna impossível. Eles entram em um estado instável ou se transformam em outra estrutura de sistema, incluindo outro nível de organização.
A temperatura e a precipitação determinam a localização dos principais biomas terrestres na superfície terrestre. O padrão de temperatura e precipitação numa determinada área durante um período de tempo bastante longo é o que chamamos de clima. O clima em diferentes regiões do globo é diferente. A precipitação anual varia de 0 a 2500 mm ou mais. Os regimes de temperatura e precipitação combinam-se de maneiras muito diferentes.
As especificidades das condições climáticas, por sua vez, determinam o desenvolvimento de um determinado bioma.
Do equador aos pólos, uma certa simetria é visível na distribuição dos biomas dos diferentes hemisférios:
Florestas tropicais (norte da América do Sul, América Central, partes ocidentais e centrais da África equatorial, Sudeste Asiático, áreas costeiras do noroeste da Austrália, ilhas dos oceanos Índico e Pacífico). O clima é sem mudança de estação (proximidade do equador), a temperatura média anual é superior a 17°C (normalmente 28°C), a precipitação média anual ultrapassa os 2400 mm.
Vegetação: as florestas dominam. Alguns tipos de árvores chegam a atingir 60 m de altura, em seus troncos e galhos há plantas epífitas, cujas raízes não atingem o solo, e trepadeiras lenhosas que se enraízam no solo e sobem nas árvores até o topo. Tudo isso forma uma copa espessa.
Fauna: a composição de espécies é mais rica do que em todos os outros biomas juntos. Particularmente numerosos são os anfíbios, répteis e aves (rãs, lagartos, cobras, papagaios), macacos e outros pequenos mamíferos, insetos exóticos com cores vivas e peixes de cores vivas nos reservatórios.
Características: os solos são frequentemente finos e pobres, a maioria dos nutrientes está contida na biomassa superficial da vegetação enraizada.
2. Savanas (África subequatorial, América do Sul, uma parte significativa do sul da Índia). O clima é seco e quente na maior parte do ano. Chuvas fortes durante a estação chuvosa. Temperatura: média anual alta. A precipitação é de 750-1650 mm/ano, principalmente durante a estação chuvosa. Vegetação - plantas de bluegrass (cereais) com árvores decíduas raras. Fauna: grandes mamíferos herbívoros, como antílopes, zebras, girafas, rinocerontes, predadores - leões, leopardos, chitas.
Desertos (algumas áreas de África, por exemplo o Saara; o Médio Oriente e a Ásia Central, a Grande Bacia e o sudoeste dos Estados Unidos e o norte do México, etc.). O clima é muito seco. Temperatura – dias quentes e noites frias. A precipitação é inferior a 250 mm/ano. Vegetação: arbustos esparsos, muitas vezes espinhosos, às vezes cactos e gramíneas baixas, cobrindo rapidamente o solo com um tapete florido após chuvas raras. As plantas têm extensos sistemas radiculares superficiais que interceptam a umidade da precipitação rara, bem como raízes principais que penetram no solo até o nível do lençol freático (30 m e mais profundo). Fauna: roedores diversos (rato-canguru, etc.), sapos, lagartos, cobras e outros répteis, corujas, águias, abutres, pequenos pássaros e insetos em grande quantidade.
4. Estepes (centro da América do Norte, Rússia, certas áreas da África e Austrália, sudeste da América do Sul). O clima é sazonal. Temperaturas - as temperaturas no verão variam de moderadamente quentes a quentes, as temperaturas no inverno abaixo de 0°C. Precipitação - 750-2000 mm/ano. Vegetação: dominada por bluegrass (cereais) de até 2 m ou mais de altura em algumas pradarias da América do Norte ou até 50 cm, por exemplo, nas estepes russas, com árvores e arbustos isolados em áreas úmidas. Fauna: grandes mamíferos herbívoros - bisões, antílopes pronghorn (América do Norte), cavalos selvagens (Eurásia), cangurus (Austrália), girafas, zebras, rinocerontes brancos, antílopes (África); Os predadores incluem coiotes, leões, leopardos, chitas, hienas, uma variedade de pássaros e pequenos mamíferos escavadores, como coelhos, esquilos terrestres e porcos-da-terra.
5. Florestas temperadas (Europa Ocidental, Leste Asiático, leste dos EUA). Clima - sazonal com temperaturas de inverno abaixo de 0 0C. Precipitação - 750-2000 mm/ano. Vegetação: dominada por florestas de árvores caducifólias de folhas largas até 35-45 m de altura (carvalho, nogueira, bordo), vegetação rasteira arbustiva, musgos, líquenes. Fauna: mamíferos (veados, porcos-espinhos, guaxinins, gambás, esquilos, coelhos, musaranhos), aves (toutinegras, pica-paus, melros, corujas, falcões), cobras, sapos, salamandras, peixes (trutas, percas, bagres, etc. . ), abundante microfauna do solo (Fig. 12.3). A biota está adaptada ao clima sazonal: hibernação, migração, dormência nos meses de inverno. 6. Florestas de coníferas, taiga (regiões do norte da América do Norte, Europa e Ásia). O clima é de invernos longos e frios, com alguma precipitação caindo em forma de neve. Vegetação: predominam florestas de coníferas perenes, principalmente abetos, pinheiros e abetos. Fauna: grandes ungulados herbívoros (veados-mula, renas), pequenos mamíferos herbívoros (lebre, esquilo, roedores), lobo, lince, raposa, urso preto, urso pardo, carcaju, vison e outros predadores, numerosos insetos sugadores de sangue no curto verão tempo. O clima é muito frio com dias polares e noites polares. A temperatura média anual é inferior a -5°C. Em poucas semanas de verão curto, o solo não descongela mais do que um metro de profundidade. A precipitação é inferior a 250 mm/ano. Vegetação: dominada por líquenes, musgos, gramíneas e ciperáceas de crescimento lento e arbustos anões. Fauna: grandes ungulados herbívoros (renas, bois almiscarados), pequenos mamíferos escavadores (durante todo o ano, por exemplo, lemingues), predadores que adquirem uma cor branca camuflada no inverno (raposa ártica, lince, arminho, coruja das neves). No curto verão, um grande número de aves migratórias nidificam na tundra, entre elas há especialmente muitas aves aquáticas, que se alimentam de insetos e invertebrados de água doce que aqui abundam. A zonação altitudinal vertical dos ecossistemas terrestres, especialmente em locais com relevo pronunciado, também é muito clara. A umidade é o principal fator que determina o tipo de bioma. Com quantidades suficientemente grandes de precipitação, a vegetação florestal se desenvolve. A temperatura determina o tipo de floresta. A situação é exatamente a mesma nos biomas de estepe e deserto. As mudanças nos tipos de vegetação em regiões frias ocorrem com menores quantidades de precipitação anual, uma vez que em baixas temperaturas se perde menos água por evaporação. A temperatura se torna um fator importante apenas em condições muito frias com permafrost. Assim, a composição dos ecossistemas depende em grande parte da sua “finalidade” funcional e vice-versa. De acordo com N.F. Reimers (1994), isto se reflete no princípio da complementaridade ecológica (complementaridade): nenhuma parte funcional do ecossistema (componente ecológico, elemento, etc.) pode existir sem outras partes funcionalmente complementares. Perto dele e ampliando-o está o princípio da congruência ecológica (correspondência):. complementando-se funcionalmente, os componentes vivos do ecossistema desenvolvem adaptações adequadas para isso, coordenadas com as condições do ambiente abiótico, que é em grande parte transformado pelos mesmos organismos (bioclima, etc.), ou seja, há uma dupla série de correspondências - entre os organismos e seu habitat - externa e criada pela cenose. 4 Estrutura do ecossistema Do ponto de vista da estrutura trófica (do grego trophe - nutrição), o ecossistema pode ser dividido em dois níveis: A camada superior - autotrófica (autoalimentada), ou "cinturão verde", incluindo plantas ou suas partes contendo clorofila, onde predomina a fixação da contagem de energia e o uso de compostos inorgânicos simples. A inferior é a camada heterotrófica (alimentada por terceiros), ou “cinturão marrom” de solos e sedimentos, matéria em decomposição, raízes, etc., na qual predomina o uso, transformação e decomposição de compostos complexos. Do ponto de vista biológico, os seguintes componentes são diferenciados como parte de um ecossistema: Num ecossistema, as ligações alimentares e energéticas entre categorias são sempre inequívocas e vão no sentido de: autotróficos - heterótrofos ou de forma mais completa; autotróficos -> consumidores -> decompositores (destruidores). A principal fonte de energia dos ecossistemas é o Sol. O fluxo de energia (de acordo com T.A. Akimova, V.V. Khaskin, 1994) enviado pelo Sol ao planeta Terra excede 20 milhões de EJ por ano. Apenas um quarto deste fluxo se aproxima do limite da atmosfera. Destes, cerca de 70% são refletidos, absorvidos pela atmosfera e emitidos na forma de radiação infravermelha de ondas longas. A radiação solar que atinge a superfície da Terra é de 1,54 milhão de EJ por ano. Esta enorme quantidade de energia é 5.000 vezes a energia total da humanidade no final do século XX e 5,5 vezes a energia de todos os recursos de combustíveis fósseis de origem orgânica disponíveis, acumulados ao longo de pelo menos 100 milhões de anos. A maior parte da energia solar que atinge a superfície do planeta é convertida diretamente em calor, aquecendo a água ou o solo, que por sua vez aquece o ar. Esse calor serve como força motriz do ciclo da água, das correntes de ar e das correntes oceânicas que determinam o clima, e é gradualmente liberado para o espaço sideral, onde se perde. Para determinar o lugar dos ecossistemas neste fluxo natural de energia, é importante perceber que, por mais extensos e complexos que sejam, utilizam apenas uma pequena parte dela. Isto implica um dos princípios básicos do funcionamento dos ecossistemas: eles existem devido à energia solar não poluente e quase eterna, cuja quantidade é relativamente constante e abundante. Vamos detalhar cada uma das características listadas da energia solar: 2. Fatores que garantem a integridade dos ecossistemas 1 Ciclos de substâncias Ambos os ciclos estão mutuamente conectados e representam, por assim dizer, um único processo. Estima-se que todo o oxigênio contido na atmosfera circula pelos organismos (combinado durante a respiração e liberado durante a fotossíntese) em 2.000 anos, o dióxido de carbono na atmosfera circula na direção oposta em 300 anos, e toda a água da Terra é decomposto e recriado através da fotossíntese e da respiração em 2.000.000 de anos. A interação de fatores abióticos e organismos vivos do ecossistema é acompanhada por uma circulação contínua de matéria entre o biótopo e a biocenose na forma de compostos orgânicos e minerais alternados. A troca de elementos químicos entre os organismos vivos e o ambiente inorgânico, cujos vários estágios ocorrem dentro de um ecossistema, é chamada de circulação biogeoquímica, ou ciclo biogeoquímico. O ciclo da água. O ciclo mais significativo da Terra em termos de massas transferidas e consumo de energia é o ciclo hidrológico planetário - o ciclo da água. A cada segundo, 16,5 milhões de m3 de água são envolvidos nele e mais de 40 bilhões de MW de energia solar são gastos nisso (de acordo com T.A. Akimova V.V. Haskin, (1994)). Mas este ciclo não consiste apenas na transferência de massas de água. São transformações de fase, formação de soluções e suspensões, precipitação, cristalização, processos de fotossíntese, bem como diversas reações químicas. A vida surgiu e continua neste ambiente. A água é o elemento básico necessário à vida. Quantitativamente, é o componente inorgânico mais comum da matéria viva. Nos humanos, a água representa 63% do peso corporal, nos cogumelos - 80%, nas plantas - 80-90% e em algumas águas-vivas - 98%. A água, como veremos um pouco mais adiante, participa do ciclo biológico e serve como fonte de hidrogênio e oxigênio, constituindo apenas uma pequena parte do seu volume total. Nos estados líquido, sólido e vapor, a água está presente em todos os três componentes principais da biosfera: a atmosfera, a hidrosfera e a litosfera. Todas as águas estão unidas pelo conceito comum de “hidrosfera”. Os componentes da hidrosfera estão interligados por constantes trocas e interações. A água, movendo-se continuamente de um estado para outro, faz ciclos pequenos e grandes. A evaporação da água da superfície do oceano, a condensação do vapor d'água na atmosfera e a precipitação na superfície do oceano formam um pequeno ciclo. Quando o vapor d'água é transportado pelas correntes de ar para a terra, o ciclo se torna muito mais complexo. Nesse caso, parte da precipitação evapora e volta para a atmosfera, a outra alimenta rios e reservatórios, mas acaba retornando ao oceano por meio do escoamento fluvial e subterrâneo, completando assim o grande ciclo. Ciclo biótico (biológico). O ciclo biótico (biológico) refere-se à circulação de substâncias entre solo, plantas, animais e microrganismos. O ciclo biótico (biológico) é o fluxo de elementos químicos do solo, água e atmosfera para os organismos vivos, a transformação dos elementos recebidos em novos compostos complexos e seu retorno no processo de vida com a queda anual de parte do orgânico matéria ou com organismos completamente mortos incluídos na composição do ecossistema. Agora apresentaremos o ciclo biótico de forma cíclica. O ciclo biótico central (de acordo com T.A. Akimova, V.V., Khaskhin) consistia em produtores unicelulares primitivos (P) e decompositores-destruidores (D). Os microrganismos são capazes de se multiplicar e se adaptar rapidamente a diferentes condições, por exemplo, utilizar todos os tipos de substratos - fontes de carbono - em sua dieta. Organismos superiores não possuem tais habilidades. Em ecossistemas completos, eles podem existir na forma de uma estrutura baseada em microrganismos. Primeiro, desenvolvem-se plantas multicelulares (P) - produtores superiores. Juntamente com organismos unicelulares, eles criam matéria orgânica através do processo de fotossíntese, utilizando a energia da radiação solar. Posteriormente, os consumidores primários são conectados - animais herbívoros (T) e depois consumidores carnívoros. Examinamos o ciclo biótico da terra. Isto aplica-se plenamente ao ciclo biótico dos ecossistemas aquáticos, por exemplo, o oceano. Todos os organismos ocupam um determinado lugar no ciclo biótico e desempenham suas funções de transformação dos ramos do fluxo de energia que recebem e de transferência de biomassa. Um sistema de decompositores unicelulares (destruidores) une a todos, despersonaliza suas substâncias e fecha o círculo geral. Eles devolvem ao ambiente abiótico da biosfera todos os elementos necessários para novas e novas revoluções. As características mais importantes do ciclo biótico devem ser enfatizadas. A fotossíntese é um poderoso processo natural que anualmente envolve enormes massas de matéria da biosfera no ciclo e determina seu alto potencial de oxigênio. Devido ao dióxido de carbono e à água, a matéria orgânica é sintetizada e o oxigênio livre é liberado. Os produtos diretos da fotossíntese são vários compostos orgânicos e, em geral, o processo de fotossíntese é bastante complexo. Além da fotossíntese com participação do oxigênio, a chamada fotossíntese oxigenada, devemos focar também na fotossíntese sem oxigênio, ou quimiossíntese. Organismos quimiossintéticos incluem nitrificadores, carboxidobactérias, bactérias sulfurosas, bactérias tiônicas de ferro e bactérias hidrogênicas. Eles recebem esse nome devido aos seus substratos de oxidação, que podem ser NH3, NO2, CO, H2S, S, Fe2+, H2. Algumas espécies são quimiolitoautotróficas obrigatórias, outras são facultativas. Estes últimos incluem carboxidobactérias e bactérias hidrogenadas. A quimiossíntese é característica das fontes hidrotermais de águas profundas. A fotossíntese ocorre, com poucas exceções, em toda a superfície da Terra, cria um enorme efeito geoquímico e pode ser expressa como a quantidade de toda a massa de carbono envolvida anualmente na construção da matéria orgânica - viva de toda a biosfera. O ciclo geral da matéria associado à construção da matéria orgânica por meio da fotossíntese envolve também elementos químicos como N, P, S, além de metais - K, Ca, Mg, Na, Al. Quando um organismo morre, ocorre o processo inverso - a decomposição da matéria orgânica por oxidação, decomposição, etc. com a formação de produtos finais de decomposição. Na biosfera terrestre, esse processo leva ao fato de que a quantidade de biomassa de matéria viva tende a ser um tanto constante. Biomassa da ecosfera (2 10|2t) é sete ordens de grandeza menor que a massa da crosta terrestre (2,10|9t). As plantas da Terra produzem anualmente matéria orgânica igual a 1,6,10"% ou 8% da biomassa da ecosfera. Os destruidores, constituindo menos de 1% da biomassa total dos organismos do planeta, processam uma massa de matéria orgânica que é 10 vezes maior que a sua própria biomassa. Em média, o período de renovação da biomassa é de 12,5 anos. Suponhamos que a massa de matéria viva e a produtividade da biosfera foram as mesmas desde o Cambriano até o presente (530 milhões de anos), então a quantidade total de matéria orgânica que passou pelo ciclo biótico global e foi utilizada pela vida no planeta será de 2,10 " 2-5,ZL08/12,5=8,5L0|9t, o que é 4 vezes a massa da crosta terrestre. Em relação a esses cálculos N.S. Pechurkin (1988) escreveu: “Podemos dizer que os átomos que constituem nossos corpos estavam em bactérias antigas, em dinossauros e em mamutes”. Lei da migração biogênica de átomos V.I. Vernadsky afirma: “A migração de elementos químicos na superfície terrestre e na biosfera como um todo ocorre ou com a participação direta da matéria viva (migração biogênica), ou ocorre em um ambiente cujas características geoquímicas (O2, CO2, H2, etc.) são determinadas matéria viva, tanto aquela que atualmente habita a biosfera quanto aquela que atuou na Terra ao longo da história geológica." DENTRO E. Vernadsky em 1928-1930 em suas profundas generalizações sobre os processos da biosfera, ele deu uma ideia das cinco principais funções biogeoquímicas da matéria viva. A primeira função é o gás. A segunda função é a concentração. A terceira função é redox. A quarta função é bioquímica. A quinta função é a atividade biogeoquímica da humanidade, cobrindo uma quantidade cada vez maior de matéria na crosta terrestre para as necessidades da indústria, dos transportes e da agricultura. O ciclo biológico varia nas diferentes zonas naturais e é classificado de acordo com um conjunto de indicadores: biomassa vegetal, serapilheira, serapilheira, quantidade de elementos fixados na biomassa, etc. A biomassa total é mais elevada na zona florestal e a proporção de órgãos subterrâneos nas florestas é mais baixa. Isso é confirmado pelo índice de intensidade do ciclo biológico - a relação entre a massa da serapilheira e a parte da serapilheira que a forma. Ciclo do carbono. De todos os ciclos biogeoquímicos, o ciclo do carbono é sem dúvida o mais intenso. O carbono circula em altas taxas entre vários meios inorgânicos e através de redes alimentares dentro de comunidades de organismos vivos. CO e CO2 desempenham um certo papel no ciclo do carbono.Muitas vezes, na biosfera da Terra, o carbono é representado pela forma mais móvel de CO2. A fonte de dióxido de carbono primário na biosfera é a atividade vulcânica associada à desgaseificação secular do manto e dos horizontes inferiores da crosta terrestre. A migração de CO2 na biosfera ocorre de duas maneiras. A primeira forma é absorvê-lo durante a fotossíntese com a formação de glicose e outras substâncias orgânicas a partir das quais são construídos todos os tecidos vegetais. Eles são posteriormente transportados através das cadeias alimentares e formam os tecidos de todos os outros seres vivos do ecossistema. Com a morte de plantas e animais na superfície, ocorre a oxidação de substâncias orgânicas com formação de CO2. Os átomos de carbono também retornam à atmosfera quando a matéria orgânica é queimada. Uma característica importante e interessante do ciclo do carbono é que em épocas geológicas distantes, centenas de milhões de anos atrás, uma parte significativa da matéria orgânica criada nos processos de fotossíntese não foi utilizada por consumidores ou decompositores, mas acumulada na litosfera. na forma de combustíveis fósseis: petróleo, carvão, xisto betuminoso, turfa, etc. Estes combustíveis fósseis são extraídos em enormes quantidades para satisfazer as necessidades energéticas da nossa sociedade industrial. Ao queimá-lo, de certa forma completamos o ciclo do carbono. Na segunda forma, a migração do carbono é realizada através da criação de um sistema carbonático em vários reservatórios, onde o CO2 se transforma em H2CO3, HCO, CO3. Com a ajuda do cálcio (ou magnésio) dissolvido em água, os carbonatos (CaCO3) são precipitados através de vias biogênicas e abiogênicas. Formam-se espessas camadas de calcário. De acordo com A. B. Ronov, a proporção entre carbono enterrado em produtos fotossintéticos e carbono em rochas carbonáticas é de 1:4. Juntamente com o grande ciclo do carbono, existem também vários pequenos ciclos do carbono na superfície terrestre e no oceano. Em geral, sem intervenção antrópica, o teor de carbono nos reservatórios biogeoquímicos: biosfera (biomassa + solo e detritos), rochas sedimentares, atmosfera e hidrosfera, é mantido com alto grau de constância (de acordo com T.A. Akimova, V.V. Haskin (1994) ). A constante troca de carbono, por um lado, entre a biosfera, e por outro, entre a atmosfera e a hidrosfera, é determinada pela função gasosa da matéria viva - os processos de fotossíntese, respiração e destruição, e equivale a cerca de 6-1010 toneladas/ano. Há um fluxo de carbono na atmosfera e na hidrosfera e durante a atividade vulcânica uma média de 4,5 106 t/ano. A massa total de carbono nos combustíveis fósseis (petróleo, gás, carvão, etc.) é estimada em 3,2*1015 toneladas, o que corresponde a uma taxa média de acumulação de 7 milhões de toneladas/ano. Essa quantidade é insignificante em comparação com a massa de carbono circulante e, por assim dizer, saiu do ciclo e se perdeu nele. Assim, o grau de abertura (imperfeição) do ciclo é 10"4, ou 0,01%, e, consequentemente, o grau de fechamento é 99,99%. Isso significa, por um lado, que cada átomo de carbono participou do ciclo dezenas de milhares de vezes antes de sair do ciclo, acabaram nas profundezas. Por outro lado, os fluxos de síntese e decadência das substâncias orgânicas na biosfera se ajustam entre si com altíssima precisão. 0,2% do estoque móvel de carbono está em circulação constante. O carbono da biomassa é renovado em 12 anos, na atmosfera - em 8 anos. Ciclo do oxigênio. O oxigênio (O2) desempenha um papel importante na vida da maioria dos organismos vivos em nosso planeta. Em termos quantitativos, este é o principal componente da matéria viva. Por exemplo, se levarmos em conta a água contida nos tecidos, o corpo humano contém 62,8% de oxigênio e 19,4% de carbono. Em geral, na biosfera esse elemento, comparado ao carbono e ao hidrogênio, é o principal entre as substâncias simples. Dentro da biosfera, há uma rápida troca de oxigênio com os organismos vivos ou com seus restos mortais após a morte. As plantas, via de regra, produzem oxigênio livre e os animais o consomem por meio da respiração. Sendo o elemento mais difundido e móvel na Terra, o oxigénio não limita a existência e as funções da ecosfera, embora a disponibilidade de oxigénio para os organismos aquáticos possa ser temporariamente limitada. O ciclo do oxigênio na biosfera é extremamente complexo, pois com ele reage um grande número de substâncias orgânicas e inorgânicas. Como resultado, ocorrem muitos epiciclos, ocorrendo entre a litosfera e a atmosfera ou entre a hidrosfera e estes dois ambientes. O ciclo do oxigênio é, em alguns aspectos, semelhante ao ciclo reverso do dióxido de carbono. O movimento de um ocorre na direção oposta ao movimento do outro. O consumo de oxigênio atmosférico e sua substituição pelos produtores primários ocorre de forma relativamente rápida. Assim, são necessários 2.000 anos para renovar completamente todo o oxigênio atmosférico. Hoje em dia não ocorre acúmulo de oxigênio na atmosfera e seu conteúdo (20,946%) permanece constante. Nas camadas superiores da atmosfera, quando a radiação ultravioleta atua sobre o oxigênio, forma-se o ozônio - O3. Cerca de 5% da energia solar que chega à Terra é gasta na formação de ozônio - cerca de 8,6 * 1015 W. As reações são facilmente reversíveis. Quando o ozônio se decompõe, essa energia é liberada, o que mantém altas temperaturas na alta atmosfera. A concentração média de ozônio na atmosfera é de cerca de 106 vol. %; a concentração máxima de O3 - até 4-10 "* vol.% - é alcançada em altitudes de 20-25 km (T.A. Akimova, V.V. Haskin, 1998). O ozônio serve como uma espécie de filtro UV: bloqueia uma porção significativa dos fortes raios ultravioleta. Provavelmente, a formação da camada de ozônio foi uma das condições para que a vida emergisse do oceano e colonizasse a terra. A maior parte do oxigênio produzido durante as épocas geológicas não permaneceu na atmosfera, mas foi fixado pela litosfera na forma de carbonatos, sulfatos, óxidos de ferro, etc. Essa massa é de 590 x 1.014 toneladas contra 39 x 1.014 toneladas de oxigênio que circula na biosfera na forma de gás ou sulfatos dissolvidos nas águas continentais e oceânicas. Ciclo do nitrogênio. O nitrogênio é um elemento biogênico essencial, pois faz parte de proteínas e ácidos nucléicos. O ciclo do nitrogênio é um dos mais complexos, pois inclui as fases gasosa e mineral e, ao mesmo tempo, os ciclos mais ideais. O ciclo do nitrogênio está intimamente relacionado ao ciclo do carbono. Via de regra, o nitrogênio segue o carbono, com o qual participa da formação de todas as substâncias proteicas. O ar atmosférico, contendo 78% de nitrogênio, é um reservatório inesgotável. No entanto, a maioria dos organismos vivos não pode utilizar directamente este azoto. Para que o nitrogênio seja absorvido pelas plantas, ele deve fazer parte de íons amônio (NH*) ou nitrato (NO3). O gás nitrogênio é continuamente liberado na atmosfera como resultado do trabalho de bactérias desnitrificantes, e as bactérias fixadoras, juntamente com as algas verde-azuladas (cianófitas), o absorvem constantemente, convertendo-o em nitratos. O ciclo do nitrogênio é claramente visível no nível dos destruidores. Proteínas e outras formas de nitrogênio orgânico contidas em plantas e animais após sua morte são expostas a bactérias heterotróficas, actinomicetos, fungos (microrganismos biorredutores), que produzem a energia de que necessitam reduzindo esse nitrogênio orgânico, convertendo-o em amônia. Nos solos ocorre o processo de nitrificação, que consiste em uma cadeia de reações, onde, com a participação de microrganismos, ocorre a oxidação do íon amônio (NH4+) a nitrito (NO~) ou do nitrito a nitrato (N0~). A redução de nitritos e nitratos aos compostos gasosos nitrogênio molecular (N2) ou óxido nitroso (N20) é a essência do processo de desnitrificação. A formação de nitratos inorgânicos em pequenas quantidades ocorre constantemente na atmosfera, ligando o nitrogênio atmosférico ao oxigênio durante descargas elétricas durante tempestades e, em seguida, caindo com a chuva na superfície do solo. Outra fonte de nitrogênio atmosférico são os vulcões, que compensam a perda de nitrogênio excluído do ciclo durante a sedimentação ou deposição no fundo dos oceanos. Em geral, o fornecimento médio de nitrogênio nitrato de origem abiótica durante a deposição da atmosfera no solo não excede 10 kg (ano/ha), as bactérias livres produzem 25 kg (ano/ha), enquanto a simbiose do Rhizobium com plantas leguminosas produz em média 200 kg (ano/ha). A parte predominante do nitrogênio fixado é processada pelas bactérias desnitrificantes em N2 e devolvida à atmosfera. Apenas cerca de 10% do nitrogênio amonificado e nitrificado é absorvido do solo pelas plantas superiores e acaba à disposição dos representantes multicelulares das biocenoses Ciclo do fósforo. O ciclo do fósforo na biosfera está associado a processos metabólicos em plantas e animais. Este importante e necessário elemento do protoplasma, contido nas plantas terrestres e nas algas 0,01-0,1%, nos animais de 0,1% a vários por cento, circula, passando gradativamente de compostos orgânicos em fosfatos, que podem novamente ser utilizados pelas plantas. No entanto, o fósforo, ao contrário de outros elementos biofílicos, não forma gás durante a migração. O reservatório de fósforo não é a atmosfera, como o nitrogênio, mas a parte mineral da litosfera. As principais fontes de fósforo inorgânico são rochas ígneas (apatitas) ou rochas sedimentares (fosforitas). Nas rochas, o fósforo inorgânico está envolvido na circulação por lixiviação e dissolução nas águas continentais. Entrando nos ecossistemas terrestres e no solo, o fósforo é absorvido pelas plantas a partir de uma solução aquosa na forma de íon fosfato inorgânico e está incluído em vários compostos orgânicos, onde aparece na forma de fosfato orgânico. O fósforo se move através das cadeias alimentares das plantas para outros organismos no ecossistema. O fósforo é transportado para os ecossistemas aquáticos pelas águas correntes. Os rios enriquecem continuamente os oceanos com fosfatos. Onde o fósforo passa a fazer parte do fitoplâncton. Alguns compostos de fósforo migram em profundidades rasas, sendo consumidos pelos organismos, enquanto a outra parte é perdida em grandes profundidades. Restos mortos de organismos levam ao acúmulo de fósforo em diferentes profundidades. Ciclo do enxofre. Existem numerosos compostos gasosos de enxofre, como o sulfeto de hidrogênio H2S e o dióxido de enxofre SO2. Porém, a parte predominante do ciclo desse elemento é de natureza sedimentar e ocorre no solo e na água.A disponibilidade de enxofre inorgânico no ecossistema é facilitada pela boa solubilidade de muitos sulfatos na água. As plantas, absorvendo sulfatos, reduzem-nos e produzem aminoácidos contendo enxofre (metionina, cisteína, cistina) que desempenham um papel importante no desenvolvimento da estrutura terciária das proteínas durante a formação de pontes dissulfeto entre diferentes zonas da cadeia polipeptídica. Muitas características básicas do ciclo biogeoquímico são claramente visíveis: S -> S -> SO2 - bactérias sulfurosas incolores, verdes e roxas; - "H2S (redução anaeróbica de sulfato) - Desulfovibno; H2S - "SO2" (oxidação aeróbica de sulfeto) - tiobacilo; S orgânico em SO e H2S. - microrganismos heterotróficos aeróbios e anaeróbios, respectivamente. A produção primária garante a incorporação do sulfato à matéria orgânica, e a excreção pelos animais serve como forma de devolver o sulfato ao ciclo. Em geral, o ecossistema requer menos enxofre em comparação com nitrogênio e fósforo. Conseqüentemente, o enxofre é menos frequentemente um fator limitante para plantas e animais. Ao mesmo tempo, o ciclo do enxofre é fundamental no processo geral de produção e decomposição de biomassa. Por exemplo, quando os sulfetos de ferro se formam nos sedimentos, o fósforo é transferido de uma forma insolúvel para uma forma solúvel e fica disponível para os organismos. Esta é uma confirmação de como um ciclo é regulado por outro. 2 Fluxo de energia nos ecossistemas Ao contrário das substâncias que circulam continuamente pelos diferentes blocos do ecossistema, que podem sempre ser reaproveitadas e entrar no ciclo, a energia pode ser utilizada uma vez, ou seja, uma vez. existe um fluxo linear de energia através do ecossistema. O influxo unilateral de energia como um fenômeno natural universal ocorre como resultado das leis da termodinâmica. A primeira lei afirma que a energia pode ser convertida de uma forma (como a luz) para outra (como a energia potencial dos alimentos), mas não pode ser criada ou destruída. A segunda lei afirma que não pode haver um único processo associado à transformação de energia sem perder parte dela. Uma certa quantidade de energia em tais transformações é dissipada em energia térmica inacessível e, portanto, perdida. Daí: não pode haver transformações, por exemplo, de substâncias alimentares na substância que compõe o corpo do organismo, ocorrendo com 100% de eficiência. Assim, os organismos vivos são conversores de energia. E cada vez que a energia é convertida, parte dela é perdida na forma de calor. Em última análise, toda a energia que entra no ciclo biótico de um ecossistema é dissipada na forma de calor. Na verdade, os organismos vivos não usam o calor como fonte de energia para realizar trabalho - eles usam luz e energia química. Cadeias e redes alimentares, níveis tróficos. Dentro de um ecossistema, substâncias que contêm energia são criadas por organismos autotróficos e servem de alimento para heterótrofos. As conexões alimentares são mecanismos de transferência de energia de um organismo para outro. Um exemplo típico: um animal come plantas. Este animal, por sua vez, pode ser comido por outro animal. Dessa forma, a energia pode ser transferida através de vários organismos - cada um subsequente se alimenta do anterior, que o fornece matéria-prima e energia. Essa sequência de transferência de energia é chamada de cadeia alimentar (trófica) ou cadeia alimentar. A localização de cada elo da cadeia alimentar é um nível trófico. O primeiro nível trófico, conforme observado anteriormente, é ocupado pelos autotróficos, ou os chamados produtores primários. Os organismos do segundo nível trófico são chamados de consumidores primários, o terceiro - consumidores secundários, etc. Geralmente existem três tipos de cadeias alimentares: A cadeia alimentar dos carnívoros começa com as plantas e passa de organismos pequenos para organismos cada vez maiores. Em terra, as cadeias alimentares consistem em três a quatro elos. Uma das cadeias alimentares mais simples se parece com: planta -> lebre -> lobo produtor -" herbívoro -> -> carnívoro As seguintes cadeias alimentares também são comuns: Material vegetal (por exemplo, néctar) - "mosca -" -" aranha -> musaranho -> coruja. Seiva da roseira -> pulgão -> -> joaninha -> -> aranha - “ave insetívora -> ave de rapina. Nos ecossistemas aquáticos e, em particular, nos marinhos, as cadeias alimentares dos predadores tendem a ser mais longas do que nos terrestres. O terceiro tipo de cadeias alimentares, começando com restos de plantas mortas, carcaças e excrementos de animais, são referidos como cadeias alimentares detríticas (saprófitas) ou cadeias de decomposição detrítica. As florestas decíduas desempenham um papel importante nas cadeias alimentares detríticas dos ecossistemas terrestres, cuja maior parte da folhagem não é consumida pelos herbívoros e faz parte da serapilheira das folhas caídas. As folhas são esmagadas por numerosos detritívoros - fungos, bactérias, insetos (por exemplo, colêmbolos), etc., e depois ingeridas por minhocas, que distribuem uniformemente o húmus na camada superficial da terra, formando o chamado mull. Neste nível, os cogumelos desenvolvem micélio. Os microrganismos em decomposição que completam a cadeia produzem a mineralização final da matéria orgânica morta. Em geral, as cadeias alimentares detríticas típicas das nossas florestas podem ser representadas da seguinte forma: Serapilheira -> minhoca -> melro - gavião; Animal morto - "larvas de moscas carniceiras -" sapo-grama -> cobra-grama comum. Nos diagramas da cadeia alimentar discutidos, cada organismo é representado alimentando-se de outros organismos de um tipo. As próprias conexões alimentares em um ecossistema são muito mais complexas, uma vez que um animal pode se alimentar de organismos de diferentes tipos da mesma cadeia alimentar ou de cadeias alimentares diferentes, por exemplo, predadores dos níveis tróficos superiores. Os animais geralmente se alimentam de plantas e de outros animais. Eles são chamados de onívoros. As teias alimentares nos ecossistemas são muito complexas e podemos concluir que a energia que entra nelas leva muito tempo para migrar de um organismo para outro. Pirâmides ecológicas. Dentro de cada ecossistema, as teias alimentares apresentam uma estrutura bem definida, que se caracteriza pela natureza e pelo número de organismos representados em cada nível das diversas cadeias alimentares. Para estudar as relações entre os organismos em um ecossistema e representá-las graficamente, eles geralmente usam pirâmides ecológicas em vez de diagramas de teias alimentares. As pirâmides ecológicas expressam a estrutura trófica de um ecossistema em forma geométrica. Eles são construídos na forma de retângulos de mesma largura, mas o comprimento dos retângulos deve ser proporcional ao valor do objeto que está sendo medido. A partir daqui você pode obter pirâmides de números, biomassa e energia. As pirâmides ecológicas refletem as características fundamentais de qualquer biocenose quando mostram sua estrutura trófica. Pirâmides de números. Eles representam a abordagem mais simples para o estudo da estrutura trófica de um ecossistema. Pirâmide de biomassa. Reflete de forma mais completa as relações nutricionais do ecossistema, pois leva em consideração a massa total dos organismos (biomassa) de cada nível trófico. Pirâmide de energia. A maneira mais fundamental de exibir conexões entre organismos em diferentes níveis tróficos é através de pirâmides energéticas. Eles representam a eficiência de conversão de energia e a produtividade das cadeias alimentares e são construídos contando a quantidade de energia (kcal) acumulada por unidade de área de superfície por unidade de tempo e utilizada pelos organismos em cada nível trófico. A energia solar recebida pela planta é apenas parcialmente utilizada no processo de fotossíntese. A energia fixada nos carboidratos representa a produção bruta do ecossistema.Os carboidratos são usados para construir o protoplasma e o crescimento das plantas. Parte de sua energia é gasta na respiração. Os consumidores de segunda ordem (predadores) não destroem toda a biomassa das suas vítimas. Além disso, da quantidade que destroem, apenas uma parte é utilizada para criar biomassa do seu próprio nível trófico. O restante é gasto principalmente na energia respiratória e é excretado com excrementos e excrementos. Em 1942, R. Lindeman formulou pela primeira vez a lei da pirâmide das energias, que nos livros didáticos é frequentemente chamada de “lei dos 10%”. De acordo com esta lei, em média, não mais do que 10% da energia passa de um nível trófico da pirâmide ecológica para outro nível. Apenas 10-20% da energia inicial é transferida para heterótrofos subsequentes. Usando a lei da pirâmide energética, é fácil calcular que a quantidade de energia que chega aos carnívoros terciários (nível trófico V) é cerca de 0,0001 da energia absorvida pelos produtores. Conclui-se que a transferência de energia de um nível para outro ocorre com eficiência muito baixa. Isto explica o número limitado de elos da cadeia alimentar, independentemente de uma biocenose específica. E. Odum (1959) numa cadeia alimentar extremamente simplificada - alfafa -> bezerro - “a criança avaliou a transformação da energia, ilustrou a magnitude das suas perdas. 4 hectares. Os bezerros se alimentam neste campo (presume-se que comem apenas alfafa), e um menino de 12 anos come exclusivamente vitela. Os resultados dos cálculos, apresentados na forma de três pirâmides: números, biomassa e energia, indicam que a alfafa utiliza apenas 0,24% de toda a energia solar que cai no campo; o bezerro absorve 8% deste produto e apenas 0,7% da biomassa do bezerro garante o desenvolvimento da criança durante o ano. E. Odum mostrou, assim, que apenas um milionésimo da energia solar incidente é convertida em biomassa carnívora, neste caso contribuindo para o aumento do peso da criança, e o restante é perdido e dissipado de forma degradada no ambiente. O exemplo acima ilustra claramente a muito baixa eficiência ecológica dos ecossistemas e a baixa eficiência de transformação nas cadeias alimentares. Podemos afirmar o seguinte: se 1000 kcal (dia m 2) registrado pelos produtores, então 10 kcal (dia m 2) vai para a biomassa dos herbívoros e apenas 1 kcal (dia m 2) - na biomassa dos carnívoros. Como uma certa quantidade de uma substância pode ser usada repetidamente por cada biocenose, e uma porção de energia uma vez, é mais conveniente dizer que ocorre uma transferência de energia em cascata no ecossistema. Os consumidores servem como um elo de gestão e estabilização do ecossistema. Os consumidores geram um espectro de diversidade na cenose, evitando o monopólio dos dominantes. A regra do valor controlador dos consumidores pode, com razão, ser considerada bastante fundamental. De acordo com as visões cibernéticas, o sistema de controle deveria ser mais complexo em estrutura do que o controlado, então fica clara a razão da multiplicidade de tipos de ativos. A importância controladora dos consumidores também tem uma base energética. O fluxo de energia através de um ou outro nível trófico não pode ser absolutamente determinado pela disponibilidade de alimento no nível trófico subjacente. Como se sabe, sobra sempre uma “reserva” suficiente, pois a destruição total dos alimentos levaria à morte dos consumidores. Esses padrões gerais são observados no âmbito dos processos populacionais, comunidades, níveis da pirâmide ecológica e biocenoses como um todo. 3 Processos dinâmicos que garantem integridade e sustentabilidade nos ecossistemas As mudanças cíclicas nas comunidades refletem a periodicidade diária, sazonal e de longo prazo das condições externas e a manifestação dos ritmos endógenos dos organismos. A dinâmica diária dos ecossistemas está associada principalmente ao ritmo dos fenômenos naturais e é de natureza estritamente periódica. Já consideramos que em cada biocenose existem grupos de organismos cuja atividade vital ocorre em diferentes horários do dia. Alguns são ativos durante o dia, outros à noite. Conseqüentemente, ocorrem mudanças periódicas na composição e proporção dos tipos individuais de biocenose de um determinado ecossistema, à medida que organismos individuais são desligados dele por um certo tempo. A dinâmica diária da biocenose é fornecida tanto por animais quanto por plantas. Como se sabe, a intensidade e a natureza dos processos fisiológicos nas plantas mudam durante o dia - a fotossíntese não ocorre à noite, muitas vezes as flores nas plantas abrem apenas à noite e são polinizadas por animais noturnos, outras estão adaptadas à polinização durante o dia. A dinâmica diária nas biocenoses, via de regra, é mais pronunciada quanto maior for a diferença de temperatura, umidade e outros fatores ambientais entre o dia e a noite. Desvios mais significativos nas biocenoses são observados com a dinâmica sazonal. Isto se deve aos ciclos biológicos dos organismos, que dependem da ciclicidade sazonal dos fenômenos naturais. Assim, a mudança de estação tem um impacto significativo na atividade vital dos animais e das plantas (hibernação, sono de inverno, diapausa e migração nos animais; períodos de floração, frutificação, crescimento ativo, queda das folhas e dormência de inverno nas plantas). A estrutura em camadas da biocenose está frequentemente sujeita à variabilidade sazonal. Camadas individuais de plantas podem desaparecer completamente nas estações apropriadas do ano, por exemplo, uma camada herbácea composta por plantas anuais. A duração das estações biológicas varia em diferentes latitudes. A este respeito, a dinâmica sazonal das biocenoses nas zonas ártica, temperada e tropical é diferente. É mais claramente expresso em ecossistemas de clima temperado e latitudes setentrionais. A variabilidade de longo prazo é normal na vida de qualquer biocenose. Assim, a quantidade de precipitação que cai na estepe florestal de Barabinsk flutua acentuadamente de ano para ano: vários anos secos alternam-se com períodos de longo prazo de precipitação abundante. Isso tem um impacto significativo em plantas e animais. Nesse caso, ocorre o desenvolvimento de nichos ecológicos – demarcação funcional no conjunto emergente ou sua adição com pouca diversidade. Mudanças de longo prazo na composição das biocenoses também se repetem em conexão com mudanças periódicas na circulação geral da atmosfera, que por sua vez são causadas por um aumento ou diminuição da atividade solar. No processo de dinâmica diária e sazonal, a integridade das biocenoses geralmente não é violada. A biocenose experimenta apenas flutuações periódicas nas características qualitativas e quantitativas. Mudanças progressivas no ecossistema levam, em última análise, à substituição de uma biocenose por outra, com um conjunto diferente de espécies dominantes. As razões para tais mudanças podem ser fatores externos à biocenose que atuam por muito tempo em uma direção, por exemplo, aumento da poluição dos corpos d'água, aumento do ressecamento dos solos pantanosos em decorrência da recuperação, aumento do pastoreio, etc. Essas mudanças de uma biocenose para outra são chamadas de exogenéticas. No caso em que a influência crescente de um fator leva a uma simplificação gradual da estrutura da biocenose, ao esgotamento de sua composição e à diminuição da produtividade, tais mudanças são chamadas de digressivas ou digressivas. As mudanças endogenéticas surgem como resultado de processos que ocorrem dentro da própria biocenose. A substituição sucessiva de uma biocenose por outra é chamada de sucessão ecológica (do Lat. - sucessão - sequência, mudança). A sucessão é um processo de autodesenvolvimento dos ecossistemas. A sucessão baseia-se na incompletude do ciclo biológico em uma determinada biocenose. Sabe-se que os organismos vivos, como resultado de sua atividade vital, alteram o ambiente ao seu redor, retirando dele algumas substâncias e saturando-o com produtos metabólicos. Quando as populações existem durante um período de tempo relativamente longo, alteram o seu ambiente numa direcção desfavorável e, como resultado, são deslocadas por populações de outras espécies, para as quais as transformações ambientais resultantes revelam-se ecologicamente benéficas. Em uma biocenose, ocorre assim uma mudança de espécie dominante. A regra (princípio) da duplicação ecológica é claramente visível aqui. A existência de uma biocenose a longo prazo só é possível se as mudanças no ambiente causadas pela atividade de alguns organismos vivos forem favoráveis para outros com necessidades opostas. Com base nas interações competitivas das espécies durante a sucessão, formam-se gradualmente combinações mais estáveis que correspondem a condições ambientais abióticas específicas. Um exemplo de sucessão que leva à substituição de uma comunidade por outra é o crescimento excessivo de um pequeno lago seguido pelo aparecimento de um pântano e depois de uma floresta em seu lugar. Primeiro, um tapete flutuante é formado ao longo das margens do lago - um tapete flutuante de juncos, musgos e outras plantas. O lago está constantemente cheio de restos de plantas mortas - turfa. Forma-se um pântano, gradualmente coberto de floresta. Uma série sucessiva de comunidades que se substituem gradual e naturalmente em sucessão é chamada de série de sucessão. As sucessões na natureza são extremamente variadas em escala. Podem ser observados em bancos com culturas, que são comunidades planctônicas - vários tipos de algas flutuantes e seus consumidores - rotíferos, flagelados em poças e lagoas, pântanos, prados, florestas, terras aráveis abandonadas, rochas desgastadas, etc. na organização dos ecossistemas Também se manifesta em processos de sucessão - transformações maiores de biocenoses são constituídas por transformações menores. Em ecossistemas estáveis com um ciclo regulado de substâncias, também ocorrem constantemente mudanças sucessivas locais, apoiando a complexa estrutura interna das comunidades. Tipos de mudanças sucessivas. Existem dois tipos principais de mudanças sucessionais: 1 - com participação de populações autotróficas e heterotróficas; 2 - com participação apenas de heterótrofos. A sucessão do segundo tipo ocorre apenas em condições em que se cria um abastecimento preliminar ou constante de compostos orgânicos, devido aos quais a comunidade existe: em montes ou pilhas de esterco, em matéria vegetal em decomposição, em reservatórios poluídos com substâncias orgânicas, etc. Processo de sucessão. Segundo F. Clements (1916), o processo de sucessão consiste nas seguintes etapas: A sucessão com mudança de vegetação pode ser primária e secundária. A sucessão primária é o processo de desenvolvimento e mudança de ecossistemas em áreas anteriormente desabitadas, começando com a sua colonização. Um exemplo clássico é a constante incrustação de rochas nuas com o eventual desenvolvimento de florestas sobre elas. Assim, nas sucessões primárias que ocorrem nas rochas dos Montes Urais, distinguem-se as seguintes etapas: A sucessão secundária é a restauração de um ecossistema que já existiu em uma determinada área. Começa quando as relações estabelecidas entre os organismos em uma biocenose estabelecida são interrompidas como resultado de uma erupção vulcânica, incêndio, derrubada, aragem, etc. As mudanças que levam à restauração da biocenose anterior são chamadas de demutacionais na geobotânica. Um exemplo é a dinâmica da diversidade de espécies na ilha de Krakatoa após a completa destruição da flora e da fauna nativas por um vulcão. Outro exemplo é a sucessão secundária da floresta escura de coníferas da Sibéria (taiga de cedro-abeto) após um incêndio florestal devastador. Em áreas mais queimadas, musgos pioneiros aparecem a partir de esporos soprados pelo vento: 3-5 anos após um incêndio, o “musgo de fogo” mais abundante é Funaria hygrometrica, Geratodon purpureus, etc. coloniza áreas queimadas), que já após 2-3 meses floresce profusamente no fogo, bem como junco moído (Calamagrostis epigeios) e outras espécies. Observam-se outras fases de sucessão: o prado de junco dá lugar a arbustos, depois a floresta de bétulas ou álamos, floresta mista de pinheiros, floresta de pinheiros, floresta de pinheiros e cedros e, finalmente, após 250 anos, ocorre a restauração da floresta de cedros e abetos. . As sucessões secundárias, via de regra, ocorrem de forma mais rápida e fácil que as primárias, pois no habitat perturbado são preservados o perfil do solo, sementes, primórdios e parte da população anterior e conexões anteriores. A demutação não é uma repetição de qualquer estágio da sucessão primária. Ecossistema clímax. A sucessão termina com um estágio em que todas as espécies do ecossistema, enquanto se reproduzem, mantêm um número relativamente constante e não ocorre mais nenhuma alteração em sua composição. Este estado de equilíbrio é denominado clímax, e o ecossistema é denominado clímax. Sob diferentes condições abióticas, diferentes ecossistemas clímax são formados. Num clima quente e úmido será uma floresta tropical, num clima seco e quente será um deserto. Os principais biomas da Terra são os ecossistemas clímax de suas respectivas áreas geográficas. Mudanças no ecossistema durante a sucessão. Produtividade e biomassa. Como já foi observado, a sucessão é um processo natural e direcionado e as mudanças que ocorrem em um ou outro estágio são características de qualquer comunidade e não dependem de sua composição de espécies ou localização geográfica. Existem quatro tipos principais de mudanças sucessivas: De referir ainda que a mudança das fases sucessórias ocorre de acordo com determinadas regras. Cada fase prepara o ambiente para o surgimento da próxima. A lei da sequência de passagem das fases de desenvolvimento opera aqui; as fases de desenvolvimento de um sistema natural só podem seguir uma ordem evolutivamente fixa (historicamente, ambientalmente determinada), geralmente de relativamente simples a complexa, como regra, sem a perda de estágios intermediários, mas possivelmente com sua passagem muito rápida ou ausência evolutivamente fixa. Quando um ecossistema se aproxima do estado de menopausa, nele, como em todos os sistemas de equilíbrio, todos os processos de desenvolvimento ficam mais lentos. Esta situação reflete-se na lei da desaceleração sucessória: os processos que ocorrem em ecossistemas maduros de equilíbrio e que se encontram num estado estável, em regra, tendem a abrandar. Nesse caso, o tipo restaurativo de sucessão muda para seu curso secular, ou seja, o autodesenvolvimento ocorre dentro dos limites da menopausa ou do desenvolvimento nodal. A lei empírica da desaceleração sucessional é uma consequência da regra de G. Odum e R. Pinkerton, ou a regra da energia máxima para manter um sistema maduro: a sucessão prossegue na direção de uma mudança fundamental no fluxo de energia no sentido de aumentar seu quantidade, visando a manutenção do sistema. A regra de G. Odum e R. Pinkerton, por sua vez, baseia-se na regra de energia máxima em sistemas biológicos, formulada por A. Lotka. Esta questão foi posteriormente bem desenvolvida por R. Margalef, Y. Odum e é conhecida como prova do princípio do “máximo zero”, ou minimização do crescimento num ecossistema maduro: um ecossistema em desenvolvimento sucessional tende a formar a maior biomassa com o menor produtividade biológica. Lindeman (1942) provou experimentalmente que a sucessão é acompanhada por um aumento na produtividade até a comunidade clímax, na qual a conversão de energia ocorre de forma mais eficiente. Dados de estudos sobre a sucessão de florestas de carvalhos e freixos mostram que nas fases posteriores a sua produtividade aumenta efectivamente. No entanto, durante a transição para uma comunidade clímax, normalmente ocorre uma diminuição na produtividade global. Assim, a produtividade nas florestas antigas é menor do que nas florestas jovens, que por sua vez podem ser menos produtivas do que as camadas herbáceas mais ricas em espécies que as precedem. Quedas semelhantes na produtividade foram observadas em alguns sistemas aquáticos. Há várias razões para isso. Uma delas é que o acúmulo de nutrientes no crescimento da biomassa florestal pode levar à diminuição da sua ciclagem. O declínio na produtividade global pode ser simplesmente o resultado da diminuição da vitalidade dos indivíduos à medida que aumenta a sua idade média na comunidade. À medida que a sucessão avança, uma proporção crescente de nutrientes disponíveis acumula-se na biomassa da comunidade e, consequentemente, o seu conteúdo na componente abiótica do ecossistema (no solo ou na água) diminui. A quantidade de detritos produzidos também aumenta. Os principais consumidores primários não são herbívoros, mas organismos detritívoros. Mudanças correspondentes também ocorrem nas redes tróficas. Os detritos se tornam a principal fonte de nutrientes. Durante a sucessão, o fechamento dos ciclos biogeoquímicos das substâncias aumenta. Aproximadamente 10 anos a partir do início da restauração da cobertura vegetal, a abertura dos ciclos diminui de 100 para 10%, e depois diminui ainda mais, atingindo um mínimo na fase de clímax. A regra do crescente fechamento do ciclo biogeoquímico das substâncias durante a sucessão, pode-se afirmar com toda a segurança, é violada pela transformação antrópica da vegetação e dos ecossistemas naturais em geral. Sem dúvida, isto leva a uma longa série de anomalias na biosfera e nas suas divisões. Uma diminuição na diversidade de espécies durante a menopausa não significa a sua baixa importância ecológica. A diversidade de espécies molda a sucessão, a sua direção e garante que o espaço real seja preenchido com vida. Um número insuficiente de espécies que compõem o complexo não poderia formar uma série de sucessão e, gradativamente, com a destruição dos ecossistemas clímax, ocorreria a desertificação completa do planeta. O valor da diversidade é funcional tanto estática quanto dinamicamente. Deve-se notar que onde a diversidade de espécies não é suficiente para formar a biosfera, que serve de base para o curso natural normal do processo de sucessão, e o próprio ambiente é fortemente perturbado, a sucessão não atinge a fase de clímax, mas termina com uma comunidade nodal - um paraclímax, uma comunidade derivada de longo ou curto prazo. Quanto mais profunda a perturbação do ambiente de um determinado espaço, mais cedo termina a sucessão de fases. Quando uma ou mais espécies são perdidas como resultado de sua destruição (desaparecimento antropogênico de habitats, menos frequentemente extinção), a chegada da menopausa não é uma restauração completa do ambiente natural. Na verdade, este é um novo ecossistema, porque nele surgiram novas conexões, muitas das antigas foram perdidas e um diferente “desgaste” de espécies se desenvolveu. O ecossistema não pode regressar ao seu antigo estado, uma vez que uma espécie perdida não pode ser restaurada. Quando qualquer factor abiótico ou biótico muda, por exemplo, com o arrefecimento sustentado, ou a introdução de uma nova espécie, uma espécie que está mal adaptada às novas condições enfrentará um de três caminhos. A lei da irreversibilidade evolutivo-ecológica afirma que um ecossistema que perdeu parte de seus elementos ou foi substituído por outro em decorrência de um desequilíbrio de componentes ecológicos não pode retornar ao seu estado original durante a sucessão se, durante as mudanças, evolutivos (microevolutivos ) ocorreram mudanças em elementos ecológicos (preservados ou perdidos temporariamente). No caso de algumas espécies serem perdidas em fases intermediárias de sucessão, essa perda pode ser compensada funcionalmente, mas não completamente. Quando a diversidade diminui para além de um nível crítico, o curso da sucessão é distorcido e, de facto, não é possível alcançar um clímax idêntico ao passado. Para avaliar a natureza dos ecossistemas restaurados, a lei da irreversibilidade ecológica evolutiva é importante. Com a perda de elementos, estas são, na verdade, formações naturais ecológicas completamente novas, com padrões e conexões recém-formados. Assim, a transferência no passado de uma espécie que saiu do ecossistema durante a sua reaclimatação não significa o seu retorno mecânico. Na verdade, trata-se da introdução de uma nova espécie em um ecossistema renovado.A lei da irreversibilidade evolutivo-ecológica enfatiza a direção da evolução não apenas no nível dos biossistemas, mas também em todos os outros níveis hierárquicos da biota. 4 A biosfera como ecossistema global que garante a integridade e a sustentabilidade dos ecossistemas A biosfera é um ecossistema global. Conforme observado anteriormente, a biosfera é dividida em geobiosfera, hidrobiosfera e aerobiosfera (Fig. 2.4). A geobiosfera possui divisões de acordo com os principais fatores formadores do ambiente: terrabiosfera e litobiosfera - dentro da geobiosfera, marinobiosfera (oceanobiosfera) e aquabiosfera - dentro da hidrobiosfera. Essas formações são chamadas de subesferas. O principal fator formador do ambiente em sua formação é a fase física do ambiente vivo: ar-água na aerobiosfera, água - água doce e salgada na hidrobiosfera, ar sólido na terrabiosfera e água sólida na litobiosfera. Por sua vez, todos eles se dividem em camadas: a aerobiosfera - na tropobiosfera e na altobiosfera; hidrobiosfera - em fotosfera, disfotosfera e afotosfera. Os fatores formadores de estrutura aqui, além do ambiente físico, energia (luz e calor), condições especiais para a formação e evolução da vida - as direções evolutivas de penetração da biota na terra, em suas profundezas, nos espaços acima do a terra, o abismo do oceano, são sem dúvida diferentes. Juntamente com a apobiosfera, a parabiosfera e outras camadas sub e supra-biosfera, constituem o chamado “bolo de camadas da vida” e a geosfera (ecosfera) da sua existência dentro dos limites da megabiosfera. Num sentido sistêmico, as formações listadas são grandes partes funcionais de dimensões virtualmente universais ou subplanetárias. Os cientistas acreditam que na biosfera existem pelo menos 8-9 níveis de ciclos de substâncias relativamente independentes dentro da interligação de 7 componentes ecológicos principais de material-energia e o 8º - informativo. Os ciclos globais, regionais e locais de substâncias não são fechados e “intersectam-se” parcialmente dentro da hierarquia do ecossistema. Este “acoplamento” material-energético e parcialmente informativo garante a integridade dos supersistemas ecológicos até à biosfera como um todo. Integridade e sustentabilidade dos ecossistemas. A biosfera é formada em maior medida não por fatores externos, mas por padrões internos. A propriedade mais importante da biosfera é a interação de seres vivos e inanimados, que se reflete na lei da migração biogênica de átomos de V. I. Vernadsky. A lei da migração biogênica dos átomos permite que a humanidade controle conscientemente os processos biogeoquímicos tanto na Terra como um todo quanto em suas regiões. A quantidade de matéria viva na biosfera, como se sabe, não está sujeita a mudanças perceptíveis. Este padrão foi formulado na forma da lei da constância da quantidade de matéria viva por V.I. Vernadsky: a quantidade de matéria viva na biosfera durante um determinado período geológico é uma constante. Na prática, esta lei é uma consequência quantitativa da lei do equilíbrio dinâmico interno do ecossistema global - a biosfera. Como a matéria viva, de acordo com a lei da migração biogênica dos átomos, é um intermediário energético entre o Sol e a Terra, sua quantidade deve ser constante ou suas características energéticas devem mudar. A lei da unidade física e química da matéria viva (toda a matéria viva da Terra é física e quimicamente unida e exclui mudanças significativas nesta última propriedade. Conseqüentemente, a estabilidade quantitativa é inevitável para a matéria viva do planeta. É totalmente característico de o número de espécies. A matéria viva, como acumulador de energia solar, deve responder simultaneamente às influências externas (cósmicas) e às mudanças internas. Uma diminuição ou aumento na quantidade de matéria viva em um local da biosfera deve levar a um processo exatamente oposto em outro local, pois os nutrientes liberados podem ser assimilados pelo restante da matéria viva ou sua deficiência será observada. Aqui devemos levar em conta a velocidade do processo, que no caso da mudança antropogénica é muito menor do que a perturbação direta da natureza pelo homem. Além da constância e constância da quantidade de matéria viva, que se reflete na lei da unidade física e química da matéria viva, na natureza viva há uma preservação constante da estrutura informacional e somática, apesar de ela mudar um pouco com o curso da evolução. Esta propriedade foi observada por Yu Goldsmith (1981) e foi chamada de lei da conservação da estrutura da biosfera - informacional e somática, ou a primeira lei da ecodinâmica. Para preservar a estrutura da biosfera, os seres vivos esforçam-se por atingir um estado de maturidade ou equilíbrio ecológico. A lei do desejo da menopausa - a segunda lei da ecodinâmica de Yu. Goldsmith, aplica-se à biosfera e outros níveis de sistemas ecológicos, embora haja especificidades - a biosfera é um sistema mais fechado do que suas subdivisões. A unidade da matéria viva da biosfera e a homologia da estrutura dos seus subsistemas levam ao facto de os elementos vivos de diferentes idades geológicas e origens geográficas originais que nela surgiram estarem intrinsecamente entrelaçados evolutivamente. O entrelaçamento de elementos de diferentes gêneses espaço-temporais em todos os níveis ecológicos da biosfera reflete a regra ou princípio da heterogênese da matéria viva. Esta adição não é caótica, mas está sujeita aos princípios da complementaridade ecológica, conformidade ecológica (congruência) e outras leis. No quadro da ecodinâmica de Yu Goldsmith, esta é a sua terceira lei - o princípio da ordem ecológica, ou mutualismo ecológico, indicando uma propriedade global devido à influência do todo sobre as suas partes, a influência inversa das partes diferenciadas sobre o desenvolvimento do todo, etc., o que no total leva à estabilidade de conservação da biosfera como um todo. A assistência mútua no âmbito da ordem ecológica, ou mutualismo sistêmico, é afirmada pela lei da ordem de preenchimento do espaço e da certeza espaço-temporal: o preenchimento do espaço dentro de um sistema natural, devido à interação entre seus subsistemas, é ordenado em de tal forma que permita que as propriedades homeostáticas do sistema sejam realizadas com contradições mínimas entre as partes dentro dele. Desta lei resulta que a existência a longo prazo de acidentes “desnecessários” à natureza, incluindo criações humanas que lhe são estranhas, é impossível. As regras da ordem sistêmica mutualística na biosfera também incluem o princípio da complementaridade do sistema, que afirma que os subsistemas de um sistema natural em seu desenvolvimento fornecem um pré-requisito para o desenvolvimento bem-sucedido e a autorregulação de outros subsistemas incluídos no mesmo sistema. A quarta lei da ecodinâmica de Yu. Goldsmith inclui a lei do autocontrole e autorregulação dos seres vivos: sistemas vivos e sistemas sob a influência controladora dos seres vivos são capazes de autocontrole e autorregulação no processo de sua adaptação às mudanças no ambiente. Na biosfera, o autocontrole e a autorregulação ocorrem durante processos em cascata e em cadeia de interação geral - durante a luta pela existência da seleção natural (no sentido mais amplo deste conceito), adaptação de sistemas e subsistemas, ampla co-evolução , etc. Além disso, todos estes processos conduzem a resultados positivos “do ponto de vista” da natureza - a preservação e o desenvolvimento dos ecossistemas da biosfera e dela como um todo. O elo de ligação entre generalizações de natureza estrutural e evolutiva é a regra de manutenção automática do habitat global: a matéria viva, no curso da autorregulação e interação com fatores abióticos, mantém autodinamicamente um ambiente de vida adequado ao seu desenvolvimento. Este processo é limitado por mudanças na escala cósmica e global da ecosfera e ocorre em todos os ecossistemas e biossistemas do planeta, como uma cascata de auto-regulação que atinge uma escala global. A regra de manutenção automática do habitat global segue os princípios biogeoquímicos de V.I. Vernadsky, as regras para a preservação dos habitats das espécies, relativa consistência interna e serve como uma constante para a presença de mecanismos conservadores na biosfera e ao mesmo tempo confirma a regra da complementaridade dinâmica do sistema. O impacto cósmico na biosfera é evidenciado pela lei de refração dos impactos cósmicos: fatores cósmicos, tendo impacto na biosfera e especialmente em suas subdivisões, estão sujeitos a alterações pela ecosfera do planeta e, portanto, em termos de força e tempo , as manifestações podem ser enfraquecidas e alteradas ou até mesmo perder completamente o seu efeito. A generalização aqui é importante devido ao facto de existir frequentemente um fluxo de efeitos sincronizados da actividade solar e de outros factores cósmicos nos ecossistemas da Terra e nos organismos que a habitam. Deve-se notar que muitos processos na Terra e em sua biosfera, embora sujeitos à influência do espaço, são assumidos ciclos de atividade solar com intervalos de 1850, 600, 400, 178, 169, 88, 83, 33, 22, 16 , 11, 5 (11,1), 6,5 e 4,3 anos, a própria biosfera e suas divisões não precisam necessariamente reagir com a mesma ciclicidade em todos os casos. As influências cósmicas do sistema da biosfera podem ser bloqueadas total ou parcialmente. Conclusão Os ecossistemas são a unidade funcional básica da ecologia, pois incluem os organismos e o ambiente inanimado - componentes que influenciam mutuamente as propriedades uns dos outros e as condições necessárias para manter a vida na forma que existe na Terra. A combinação de um ambiente físico-químico específico (biótopo) com uma comunidade de organismos vivos (biocenose) forma um ecossistema. Existem microecossistemas, mesoecossistemas e o ecossistema global - a biosfera. Os ecossistemas não estão dispersos desordenadamente; pelo contrário, estão agrupados em zonas bastante regulares, tanto horizontalmente (em latitude) como verticalmente (em altura). A principal fonte de energia dos ecossistemas é o Sol. A radiação solar que atinge a superfície da Terra é de 1,54 milhão de EJ por ano. A maior parte da energia solar que atinge a superfície do planeta é convertida diretamente em calor, aquecendo a água ou o solo, que por sua vez aquece o ar. Os ecossistemas existem devido à energia solar não poluente e quase eterna, cuja quantidade é relativamente constante e abundante. A energia solar na Terra causa dois ciclos de substâncias: grandes, ou geológicas, e pequenas, biológicas (bióticas). Ambos os ciclos estão mutuamente conectados e representam, por assim dizer, um único processo. A existência de ciclos biogeoquímicos, ou ciclos biogeoquímicos, cria a oportunidade de autorregulação (homeostase) do sistema, o que confere estabilidade ao ecossistema: uma incrível constância do conteúdo percentual de vários elementos. Existem muitos ciclos de substâncias. O ciclo mais significativo na Terra em termos de massas transferidas e consumo de energia é o ciclo hidrológico planetário ou ciclo da água. O ciclo biótico (biológico) refere-se à circulação de substâncias entre solo, plantas, animais e microrganismos. De todos os ciclos biogeoquímicos, o ciclo do carbono é sem dúvida o mais intenso. A maior parte do oxigênio produzido durante as épocas geológicas não permaneceu na atmosfera, mas foi fixado pela litosfera na forma de carbonatos, sulfatos, óxidos de ferro, etc. Essa massa é de 590x1014 toneladas contra 39x1014 toneladas de oxigênio que circula na biosfera na forma de gás ou sulfatos dissolvidos em águas continentais e oceânicas. O nitrogênio é um elemento biogênico essencial, pois faz parte de proteínas e ácidos nucléicos. O ciclo do nitrogênio é um dos mais complexos, pois inclui as fases gasosa e mineral e, ao mesmo tempo, os ciclos mais ideais. O ciclo do fósforo na biosfera está associado a processos metabólicos em plantas e animais. Este importante e necessário elemento do protoplasma, contido nas plantas terrestres e nas algas 0,01-0,1%, nos animais de 0,1% a vários por cento, circula, passando gradativamente de compostos orgânicos em fosfatos, que podem novamente ser utilizados pelas plantas. Existem numerosos compostos gasosos de enxofre, como o sulfeto de hidrogênio H2S e o dióxido de enxofre SO2. A manutenção da atividade vital dos organismos e da circulação da matéria nos ecossistemas, ou seja, a existência dos ecossistemas, depende do fluxo constante de energia necessária a todos os organismos para as suas funções vitais e auto-reprodução. Ao contrário das substâncias que circulam continuamente pelos diferentes blocos do ecossistema, que podem sempre ser reutilizadas e entrar no ciclo, a energia pode ser utilizada uma vez, ou seja, existe um fluxo linear de energia através do ecossistema. Dentro de um ecossistema, substâncias que contêm energia são criadas por organismos autotróficos e servem de alimento para heterótrofos. As conexões alimentares são mecanismos de transferência de energia de um organismo para outro. Dentro de cada ecossistema, as teias alimentares apresentam uma estrutura bem definida, que se caracteriza pela natureza e pelo número de organismos representados em cada nível das diversas cadeias alimentares. A formação de ecossistemas é um processo dinâmico. Nos ecossistemas, ocorrem constantemente mudanças no estado e na atividade vital dos seus membros e na proporção das populações. As diversas mudanças que ocorrem em qualquer comunidade dividem-se em dois tipos principais: cíclicas e progressivas. A sucessão é um processo natural e direcionado e as mudanças que ocorrem em um ou outro estágio são características de qualquer comunidade e não dependem de sua composição de espécies ou localização geográfica. O ecossistema global é a biosfera. Conforme observado anteriormente, a biosfera é dividida em geobiosfera, hidrobiosfera e aerobiosfera. A biosfera é formada em maior medida não por fatores externos, mas por padrões internos. A propriedade mais importante da biosfera é a interação de seres vivos e inanimados, que se reflete na lei da migração biogênica de átomos de V.I. Vernadsky. Os ecossistemas existentes na Terra são diversos. Existem microecossistemas (por exemplo, o tronco de uma árvore podre), mesoecossistemas (floresta, lagoa, etc.), macroecossistemas (continente, oceano, etc.) e a biosfera global. Grandes ecossistemas terrestres são chamados de biomas. Cada bioma contém vários ecossistemas menores e interconectados. Existem várias classificações de ecossistemas: floresta tropical perene; deserto: gramado e arbustivo; pastagens tropicais e savanas; estepe temperada; floresta decídua temperada: florestas boreais de coníferas. Tundra: ártica e alpina. Tipos de ecossistemas de água doce: fita (água parada): lagos, lagoas, etc.; lótica (águas correntes): rios, riachos, etc.; zonas húmidas: pântanos e florestas pantanosas; Tipos de ecossistemas marinhos: oceano aberto (pelágico); águas da plataforma continental (águas costeiras); áreas de ressurgência (áreas férteis com pesca produtiva); estuários (baías costeiras, estreitos, fozes de rios, sapais, etc.). Todo ecossistema tem dois componentes principais: organismos e fatores de seu ambiente inanimado. A totalidade dos organismos (plantas, animais, micróbios) é chamada de biota de um ecossistema. As formas de interação entre diferentes categorias de organismos são a sua estrutura biótica. A energia solar na Terra causa dois ciclos de substâncias: grande, ou geológico, mais claramente manifestado no ciclo da água e na circulação atmosférica, e pequeno, biológico (biótico), desenvolvendo-se com base no grande e consistindo em um ciclo contínuo, cíclico, mas desigual no tempo e no espaço, e acompanhado por perdas mais ou menos significativas na redistribuição natural da matéria, energia e informação dentro dos sistemas ecológicos de vários níveis de organização. Ambos os ciclos estão mutuamente conectados e representam, por assim dizer, um único processo. A interação de fatores abióticos e organismos vivos do ecossistema é acompanhada por uma circulação contínua de matéria entre o biótopo e a biocenose na forma de compostos orgânicos e minerais alternados. A troca de elementos químicos entre os organismos vivos e o ambiente inorgânico, cujos vários estágios ocorrem dentro de um ecossistema, é chamada de ciclo biogeoquímico, ou ciclo biogeoquímico. A existência de tais ciclos cria a oportunidade de autorregulação (homeostase) do sistema, o que confere estabilidade ao ecossistema: uma incrível constância da porcentagem de vários elementos. Aqui se aplica o princípio de funcionamento dos ecossistemas: a aquisição de recursos e a eliminação de resíduos ocorrem no quadro do ciclo de todos os elementos. Lista de literatura usada 1.Bigon M.I. Ecologia, indivíduos, populações e comunidades. Moscou: 1989. - 290 p. 2.Beysenova A.S., Shildebaev Zh.B., Sautbaeva G.Z. Ecologia. Almaty: "Gylym, 2001. - 201 p. 3.Akimova T.A., Khaskin V.V. Ecologia. M.: Editora UNIDADE, 1998. - 233 p. .Gorelov A.A. Ecologia. Curso de palestras. M.: "Centro" 1997. - 280 p. .Stadnitsky G.V., Rodionov A.I. Ecologia. M.: Editora UNIDADE, 1996. - 272 p. .Sadanov A.K., Svanbaeva Z.S., Ecologia. Almaty: "Agrouniversidade", 1999. - 197 p. .Shilov I.A. Ecologia. M.: Ensino Superior, 2000. - 348 p. .Reimers N.F. Ecologia (Teoria, leis, regras, princípios e hipóteses). M.: "Jovem Rússia", 1994. - 260 p. .Tsvetkova L.I., Alekseev M.I., Usenov B.P. e outros Ecologia. Livro didático para universidades técnicas. M.; ASV, São Petersburgo: Khimizdat, 1999, 185 p. .Girusov E.V. Ecologia e economia da gestão ambiental. M.: Direito e Direito, UNIDADE. 1998 - 232 p. .Vronsky V.A. Ecologia aplicada: livro didático. Beneficiar. Rostov do Don: 1995. - 197 p. .Budyko M.I. Ecologia global. M.: Mysl, 1977.-248 p. .Alekseenko V.A. Geoquímica ambiental. Livro didático. M.: Logos, 2000 - 410 p. .Petrov K.M. Ecologia geral de S-P. Tutorial. "Química", 1997. - 218 p. .Andersen J.M. Ecologia e ciência ambiental. Biosfera, ecossistema, homem. L., 1985. 376 p. .Girenok F.I. Ecologia. Civilização. Noosfera.M.: 191987. - 281 p. .Em busca do equilíbrio: a ecologia no sistema de prioridades políticas sociais M.: 1992. - 427 p. .História da interação entre sociedade e natureza: Fatores e conceito. M.: ANSSSR 1990. .Stepanovskikh A.S. Ecologia aplicada: Proteção ambiental. Livro didático para universidades. M.: UNIDADE DANA. 2003. 751 pág. Trabalhos semelhantes a - Ciclo de substâncias e fluxos de energia nos ecossistemas
Os termos “fluxo de matéria” e “fluxo de energia” devem ser claramente definidos. O fluxo de matéria é o movimento na forma de elementos químicos e seus compostos, dos produtores aos decompositores (através ou sem consumidores). O fluxo de energia é a transferência de energia na forma de ligações químicas de compostos orgânicos (alimentos) ao longo das cadeias alimentares de um nível trófico para outro (superior).
Deve-se notar que, ao contrário das substâncias que circulam constantemente pelos diferentes blocos do ecossistema e podem sempre reentrar no ciclo, a energia que entra só pode ser utilizada uma vez.
Como fenômeno natural universal, o fluxo unidirecional de energia é determinado pelas leis da termodinâmica. De acordo com a primeira lei, a energia pode passar de uma forma (a energia da luz) para outra (a energia potencial dos alimentos), mas nunca é criada novamente e não desaparece sem deixar vestígios.
A segunda lei da termodinâmica afirma que não pode haver um único processo associado à transformação de energia sem perder parte dela. Por isso, não pode haver transformação com 100% de eficiência, por exemplo, do alimento na substância que constitui o corpo do organismo.
Assim, o funcionamento de todos os ecossistemas é determinado por um fluxo constante de energia, necessário para que todos os organismos mantenham a sua existência e auto-reprodução.
Existem também relações competitivas nos ecossistemas. Nesse aspecto, a lei da maximização energética (G. Odum - E. Odum) é de grande interesse: na competição com outros ecossistemas, sobrevive aquele que melhor contribui para o fornecimento de energia e utiliza sua quantidade máxima da forma mais eficiente. (conservas). De acordo com a lei, para isso o sistema: 1) cria acumuladores (armazenamentos) de energia de alta qualidade (por exemplo, reservas de gordura); 2) gasta uma certa quantidade de energia acumulada para garantir o fornecimento de nova energia; 3) garante a circulação de diversas substâncias; 4) cria mecanismos regulatórios que apoiam a estabilidade do sistema e a sua capacidade de adaptação às condições em mudança; 5) estabelece trocas com outros sistemas necessários para atender a necessidade de tipos especiais de energia.
É necessário enfatizar uma circunstância importante: a lei da maximização da energia também é válida em relação à informação, portanto (segundo N. F. Reimers) também pode ser considerada como a lei da maximização da energia e da informação: o sistema que mais contribui para o recebimento, produção e uso eficaz de energia e informação.
Notemos que o fornecimento máximo de uma substância, enquanto tal, ainda não garante o sucesso do sistema num grupo competitivo de outros sistemas semelhantes.
Foi observado anteriormente que fortes relações alimentares, ou uma cadeia alimentar, surgem e se estabelecem entre os organismos da biocenose. Este último consiste em três elos principais: produtores, consumidores e decompositores.
As cadeias alimentares que começam com organismos fotossintéticos são chamadas de cadeias de pastoreio (ou pastoreio), e as cadeias que começam com matéria vegetal morta, carcaças e excrementos de animais são chamadas de cadeias detríticas.
A localização de cada elo da cadeia alimentar é chamada de nível trófico; é caracterizado por diferentes intensidades de fluxo de substâncias e energia. O primeiro nível trófico é sempre o dos produtores; o segundo são consumidores herbívoros; o terceiro são carnívoros que vivem de formas herbívoras; quarto nível – consumir outros carnívoros, etc.
Existem consumidores de primeira, segunda, terceira e quarta ordens, ocupando diferentes níveis da cadeia alimentar (Fig. 9).
Arroz. 9.
É óbvio que a especialização alimentar dos consumidores desempenha um papel importante neste contexto. Espécies com uma ampla variedade de nutrição podem ser incluídas em cadeias alimentares em diferentes níveis tróficos. A dieta de, por exemplo, uma pessoa inclui alimentos vegetais e carne de herbívoros e carnívoros. Portanto, atua em diferentes cadeias alimentares como consumidor das ordens I, II ou III.
Como a energia é perdida ao transferir energia de um nível para outro, a cadeia de energia não pode ser longa: geralmente consiste em 4...6 elos (Tabela 1).
1. Diagramas típicos de cadeias alimentares (de acordo com V. M. Ivonin, 1996)
Porém, tais cadeias em sua forma pura geralmente não são encontradas na natureza, uma vez que a mesma espécie pode estar presente simultaneamente em diferentes elos. Isso se deve ao fato de haver poucos monófagos na natureza; oligófagos e polífagos são muito mais comuns. Por exemplo, predadores que se alimentam de vários herbívoros e carnívoros são elos de muitas cadeias. Como resultado, em cada biocenose, complexos de cadeias alimentares são formados evolutivamente, representando um todo único. É assim que se criam redes de poder, que são muito complexas.
Assim, podemos concluir que a cadeia alimentar é o principal canal de transferência de energia na comunidade (entre plantas - produtores, animais - consumidores e microrganismos - decompositores) (Fig. 10).Já no diagrama fica claro que a ideia de cadeias alimentares e níveis tróficos é uma abstração. Uma cadeia linear com níveis claramente separados pode ser criada em laboratório. Mas na natureza existem realmente redes tróficas nas quais muitas populações pertencem a vários níveis tróficos ao mesmo tempo. O mesmo organismo consome animais e plantas; o predador pode se alimentar de consumidores de primeira e segunda ordem; muitos animais comem plantas vivas e mortas.
Devido à complexidade das relações tróficas, a perda de uma espécie muitas vezes quase não tem efeito sobre a comunidade. Outros “usuários” passam a consumir os alimentos das espécies extintas,

Arroz. 10.
as espécies que dele se alimentam encontram novas fontes de alimento: em geral, o equilíbrio é mantido na comunidade.
A energia absorvida pelos produtores flui através das cadeias alimentares e é gradualmente consumida. No final de uma cadeia alimentar a quantidade de energia é sempre menor do que no início. Durante a fotossíntese, as plantas retêm, em média, apenas cerca de 1% da energia solar que as atinge. Um animal que comeu uma planta não digere parte do alimento e o excreta na forma de excremento. Normalmente 20...60% dos alimentos vegetais são digeridos; A energia absorvida é utilizada para manter a vida do animal. O funcionamento das células e órgãos é acompanhado pela liberação de calor, ou seja, com isso uma proporção significativa da energia alimentar é logo dissipada no ambiente. Uma porção relativamente pequena de comida é usada para construir novos tecidos e criar reservas de gordura. A seguir, um predador que comeu um herbívoro e representa o terceiro nível trófico recebe apenas aquela energia da energia acumulada pela planta que fica retida no corpo de sua presa (segundo nível) na forma de aumento de biomassa.
Sabe-se que em cada etapa da transferência de matéria e energia na cadeia alimentar, aproximadamente 90% da energia é perdida e apenas cerca de um décimo dela passa para o próximo consumidor, ou seja, a transferência de energia nas ligações alimentares dos organismos obedece à “regra dos dez por cento” (princípio de Lindemann). Por exemplo, a quantidade de energia que atinge os carnívoros terciários (quinto nível trófico) é apenas cerca de 10 -4 da energia absorvida pelos produtores. Isto explica o número limitado (5...6) de elos (níveis) na cadeia alimentar, independentemente da complexidade da composição de espécies da biocenose.

Arroz. onze.
Considerando o fluxo de energia nos ecossistemas, também é fácil compreender porque é que a biomassa diminui à medida que o nível trófico aumenta. Aqui se manifesta o terceiro princípio básico do funcionamento dos ecossistemas: quanto maior a biomassa de uma população, menor deve ser o nível trófico que ela ocupa, ou caso contrário: no final de longas cadeias alimentares não pode haver uma grande biomassa.
Os três princípios básicos do funcionamento do ecossistema listados acima - o ciclo dos nutrientes, o fluxo da energia solar e a diminuição da biomassa com o aumento do nível trófico - podem ser apresentados na forma de um diagrama geral (Fig. 11). Se organizarmos os organismos de acordo com as suas relações nutricionais, indicando para cada um deles a “entrada” e a “saída” de energia e nutrientes, torna-se óbvio que os nutrientes são continuamente reciclados dentro do ecossistema, e o fluxo de energia passa através dele.
A manutenção da atividade vital dos organismos e da circulação da matéria nos ecossistemas, ou seja, a existência dos ecossistemas, depende do fluxo constante de energia necessária a todos os organismos para as suas funções vitais e auto-reprodução.
Ao contrário das substâncias que circulam continuamente pelos diferentes blocos do ecossistema, que podem sempre ser reutilizadas e entrar no ciclo, a energia pode ser utilizada apenas uma vez, ou seja, existe um fluxo linear de energia através do ecossistema.
O influxo unilateral de energia como um fenômeno natural universal ocorre como resultado das leis da termodinâmica. Primeira Lei afirma que a energia pode ser convertida de uma forma (como a luz) para outra (como a energia potencial dos alimentos), mas não pode ser criada ou destruída. Segunda Lei afirma que não pode haver um único processo associado à transformação de energia sem perder parte dela. Uma certa quantidade de energia em tais transformações é dissipada em energia térmica inacessível e, portanto, perdida. Assim, não pode haver transformações, por exemplo, de substâncias alimentares na substância que compõe o corpo do organismo, que ocorrem com 100 por cento de eficiência.
Assim, os organismos vivos são conversores de energia. E cada vez que a energia é convertida, parte dela é perdida na forma de calor. Em última análise, toda a energia que entra no ciclo biótico de um ecossistema é dissipada na forma de calor. Na verdade, os organismos vivos não usam o calor como fonte de energia para realizar trabalho - eles usam luz e energia química.
Pirâmides ecológicas. Dentro de cada ecossistema, as teias alimentares apresentam uma estrutura bem definida, que se caracteriza pela natureza e pelo número de organismos representados em cada nível das diversas cadeias alimentares. Para estudar as relações entre os organismos em um ecossistema e representá-las graficamente, eles geralmente usam pirâmides ecológicas em vez de diagramas de teias alimentares. As pirâmides ecológicas expressam a estrutura trófica de um ecossistema em forma geométrica. Eles são construídos na forma de retângulos de mesma largura, mas o comprimento dos retângulos deve ser proporcional ao valor do objeto que está sendo medido. A partir daqui você pode obter pirâmides de números, biomassa e energia.
As pirâmides ecológicas refletem as características fundamentais de qualquer biocenose quando mostram sua estrutura trófica:
A sua altura é proporcional ao comprimento da cadeia alimentar em questão, ou seja, ao número de níveis tróficos que contém;
A sua forma reflete mais ou menos a eficiência das transformações de energia durante a transição de um nível para outro.
Pirâmides de números. Eles representam a abordagem mais simples para o estudo da estrutura trófica de um ecossistema. Nesse caso, primeiro é contado o número de organismos em um determinado território, agrupados por níveis tróficos e apresentados na forma de um retângulo, cujo comprimento (ou área) é proporcional ao número de organismos que vivem em uma determinada área ( ou em determinado volume, se for um ecossistema aquático). Foi estabelecida uma regra básica de que em qualquer ambiente existem mais plantas que animais, mais herbívoros que carnívoros, mais insetos que pássaros, etc.
As pirâmides populacionais refletem a densidade dos organismos em cada nível trófico. Existe uma grande diversidade na construção de diversas pirâmides populacionais. Muitas vezes eles estão de cabeça para baixo.
Por exemplo, numa floresta há significativamente menos árvores (produtores primários) do que insetos (herbívoros).
Pirâmide de biomassa. Reflete de forma mais completa as relações alimentares no ecossistema, uma vez que leva em conta a massa total dos organismos (biomassa) cada nível trófico. Os retângulos nas pirâmides de biomassa representam a massa de organismos em cada nível trófico por unidade de área ou volume. A forma da pirâmide de biomassa é muitas vezes semelhante à forma da pirâmide populacional. Uma diminuição na biomassa em cada nível trófico sucessivo é característica.
As pirâmides de biomassa, assim como os números, podem ser não apenas retas, mas também invertidas. As pirâmides invertidas de biomassa são características de ecossistemas aquáticos, nos quais os produtores primários, como as algas fitoplanctônicas, se dividem muito rapidamente, e seus consumidores - os crustáceos zooplanctônicos - são muito maiores, mas possuem um longo ciclo de reprodução. Em particular, isto aplica-se a ambientes de água doce, onde a produtividade primária é fornecida por organismos microscópicos cujas taxas metabólicas são aumentadas, isto é, a biomassa é baixa, a produtividade é elevada.
Pirâmide de energia. A maneira mais fundamental de exibir conexões entre organismos em diferentes níveis tróficos é através de pirâmides energéticas. Eles representam a eficiência de conversão de energia e a produtividade das cadeias alimentares e são construídos contando a quantidade de energia (kcal) acumulada por unidade de área de superfície por unidade de tempo e utilizada pelos organismos em cada nível trófico. Assim, é relativamente fácil determinar a quantidade de energia armazenada na biomassa, mas é mais difícil estimar a quantidade total de energia absorvida em cada nível trófico. Tendo construído um gráfico (Fig. 12.28), podemos afirmar que destruidores, cuja importância parece pequena na pirâmide de biomassa, e vice-versa na pirâmide populacional; recebem uma parcela significativa da energia que passa pelo ecossistema. Além disso, apenas parte de toda esta energia permanece nos organismos em cada nível trófico do ecossistema e é armazenada na biomassa; o resto é utilizado para satisfazer as necessidades metabólicas dos seres vivos: manutenção da existência, crescimento, reprodução. Os animais também gastam uma quantidade significativa de energia no trabalho muscular.
R. Lindeman formulou pela primeira vez em 1942 lei da pirâmide energética, que nos livros didáticos é frequentemente chamada de “lei dos 10%”. De acordo com esta lei, de um nível trófico da pirâmide ecológica Em média, não mais que 10% da energia passa para outro nível.
Os consumidores servem como um elo de gestão e estabilização do ecossistema. Os consumidores geram um espectro de diversidade na cenose, evitando o monopólio dos dominantes. Regra de controle do valor dos consumidores pode ser legitimamente considerado bastante fundamental. De acordo com as visões cibernéticas, o sistema de controle deveria ser mais complexo em estrutura do que o controlado, então fica clara a razão da multiplicidade de tipos de consumidores. A importância controladora dos consumidores também tem uma base energética. O fluxo de energia através de um ou outro nível trófico não pode ser absolutamente determinado pela disponibilidade de alimento no nível trófico subjacente. Como se sabe, sobra sempre uma “reserva” suficiente, pois a destruição total dos alimentos levaria à morte dos consumidores. Esses padrões gerais são observados no âmbito dos processos populacionais, comunidades, níveis da pirâmide ecológica e biocenoses como um todo.
Para usar visualizações de apresentações, crie uma conta do Google e faça login nela: https://accounts.google.com
Legendas dos slides:
Ciclo de matéria e energia na natureza
O ciclo das substâncias são os processos repetidos de transformação e movimento da matéria na natureza, que são mais ou menos cíclicos. Todas as substâncias do nosso planeta estão em processo de circulação. Existem dois ciclos principais na natureza: Grande (geológico) Pequeno (biogeoquímico)
O Grande Ciclo das Substâncias O Grande Ciclo dura milhões de anos e é causado pela interação da energia solar com a energia profunda da Terra. Associado a processos geológicos, a formação e destruição de rochas e a subsequente movimentação de produtos de destruição.
Pequeno ciclo de substâncias O pequeno ciclo (biogeoquímico) ocorre dentro da biosfera, ao nível da biocenose. Sua essência é a formação de matéria viva a partir de compostos inorgânicos durante o processo de fotossíntese e a transformação da matéria orgânica durante a decomposição em compostos inorgânicos. Ciclos biogeoquímicos – Vernadsky V.I.
Ciclo da água Tr escoamento inf Evaporação de água Condensação de vapor Escoamento de precipitação Transpiração infiltração
A transpiração é o processo de movimentação da água através de uma planta e sua evaporação através dos órgãos externos da planta, como folhas, caules e flores. A água é essencial para a vida das plantas, mas apenas uma pequena porção da água fornecida pelas raízes é usada diretamente para o crescimento e o metabolismo.
O ciclo da água
Ciclo da água A maior parte da água está concentrada nos oceanos. A água que evapora de sua superfície abastece ecossistemas terrestres naturais e artificiais. Quanto mais próxima uma área estiver do oceano, mais precipitação haverá. A terra devolve constantemente água ao oceano: parte da umidade evapora, mais ativamente nas florestas, parte é coletada pelos rios: eles recebem chuva e derretem a água. A troca de umidade entre o oceano e a terra exige custos energéticos muito elevados: aproximadamente 30% da energia solar que chega à Terra é gasta nisso.
Influência humana no ciclo da água O ciclo da água na biosfera antes do desenvolvimento da civilização estava em equilíbrio, ou seja, o oceano recebeu tanta água dos rios quanto consumiu por evaporação. Com o desenvolvimento da civilização, este ciclo começou a ser interrompido. Em particular, as florestas evaporam cada vez menos água porque... sua área está diminuindo, e a superfície do solo, ao contrário, está aumentando, porque A área de terras agrícolas irrigadas está aumentando. terras. Os rios das regiões do sul tornaram-se rasos. A água evapora pior da superfície do oceano, porque... uma parte significativa dele é coberta por uma película de óleo. Tudo isso piora o abastecimento de água à biosfera.
As secas estão a tornar-se mais frequentes e estão a surgir bolsas de desastres ambientais. Por exemplo, uma seca catastrófica já dura mais de 35 anos na África, na zona do Sahel - uma região semidesértica que separa o Saara dos países do norte do continente. A água doce que retorna da terra para o oceano e outras massas de água é frequentemente poluída. A água de muitos rios russos tornou-se praticamente imprópria para beber. A proporção de água doce disponível para os organismos vivos é bastante pequena, por isso deve ser usada com moderação e não poluída! Cada quarta pessoa no planeta carece de água potável. Em muitas áreas do mundo não há água suficiente para a produção industrial e irrigação.
Diferentes componentes da hidrosfera participam do ciclo da água de diferentes maneiras e em velocidades diferentes. Para a renovação completa da água nas geleiras são necessários 8.000 anos, águas subterrâneas – 5.000 anos, oceano – 3.000 anos, solo – 1 ano. O vapor atmosférico e as águas dos rios são completamente renovados em 10–12 dias. O ciclo da água na natureza leva cerca de 1 milhão de anos.
Ciclo do oxigênio O oxigênio faz parte dos elementos mais comuns da biosfera. O teor de oxigênio na atmosfera é de quase 21%. O oxigênio faz parte das moléculas de água e dos organismos vivos (proteínas, gorduras, carboidratos, ácidos nucléicos). O oxigênio é produzido pelos produtores (plantas verdes). O ozônio desempenha um papel importante no ciclo do oxigênio. A camada de ozônio está localizada a uma altitude de 20 a 30 km acima do nível do mar. O teor de oxigênio na atmosfera é afetado por 2 processos principais: 1) fotossíntese 2) decomposição da matéria orgânica, durante a qual é consumida.
O ciclo do oxigênio é um processo lento. Demora cerca de 2.000 anos para que todo o oxigênio da atmosfera seja completamente renovado. Para efeito de comparação: a renovação completa do dióxido de carbono na atmosfera ocorre em cerca de 3 anos. O oxigênio é usado para respiração pela maioria dos organismos vivos. O oxigênio é utilizado na queima de combustível em motores de combustão interna, nos fornos de usinas termelétricas, em motores de aeronaves e foguetes, etc. O consumo antropogénico adicional pode perturbar o equilíbrio do ciclo do oxigénio. Até agora, a biosfera está a compensar a intervenção humana: as perdas são compensadas por plantas verdes. Com a redução adicional da área florestal e a queima de cada vez mais combustível, o teor de oxigênio na atmosfera começará a diminuir.
É IMPORTANTE!!! Quando o teor de oxigênio no ar diminui para 16%, a saúde da pessoa piora (o coração sofre principalmente), para 7% a pessoa perde a consciência e para 3% ocorre a morte.
Ciclo do carbono
Ciclo do carbono O carbono é a base dos compostos orgânicos; faz parte de todos os organismos vivos na forma de proteínas, gorduras e carboidratos. O carbono entra na atmosfera na forma de dióxido de carbono. Na atmosfera, onde se concentra a maior parte do dióxido de carbono, ocorre uma troca constante: as plantas absorvem o dióxido de carbono durante a fotossíntese e todos os organismos o liberam durante a respiração. Até 50% do carbono na forma de CO 2 é devolvido à atmosfera pelos decompositores - microrganismos do solo. O carbono sai do ciclo como carbonato de cálcio.
Influência humana no ciclo do carbono A atividade humana tecnogênica perturba o equilíbrio natural do ciclo do carbono: 1) durante a combustão de combustível orgânico, cerca de 6 bilhões de toneladas de CO 2 são emitidas anualmente na atmosfera: a) Produção de eletricidade em usinas termelétricas b ) Gases de escapamento de automóveis 2) destruição de florestas. Nos últimos 100 anos, o nível de dióxido de carbono na atmosfera tem crescido de forma constante e rápida. Dióxido de carbono + metano + vapor de água + ozônio + óxidos de nitrogênio = gás de efeito estufa. O resultado é o efeito estufa – aquecimento global, que pode levar a desastres naturais em grande escala.
Ciclo do nitrogênio Na forma livre, o nitrogênio é um componente do ar - 78%. O nitrogênio é um dos elementos mais importantes para a vida dos organismos. O nitrogênio faz parte de todas as proteínas. A molécula de nitrogênio é muito forte, razão pela qual a maioria dos organismos não consegue absorver o nitrogênio atmosférico. O nitrogênio é absorvido pelos organismos vivos apenas na forma de compostos com hidrogênio e oxigênio. A fixação de nitrogênio em compostos químicos ocorre como resultado da atividade vulcânica e de trovoadas, mas principalmente como resultado da atividade de microrganismos - fixadores de nitrogênio (bactérias fixadoras de nitrogênio e algas verde-azuladas).
O nitrogênio entra nas raízes das plantas na forma de nitratos, que são utilizados para a síntese de matéria orgânica (proteínas). Os animais consomem nitrogênio através de alimentos vegetais ou animais. O nitrogênio é devolvido à atmosfera através da decomposição de matéria orgânica morta. As bactérias do solo decompõem as proteínas em substâncias inorgânicas - gases - amônia, óxidos de nitrogênio, que entram na atmosfera. O nitrogênio que entra nos corpos d'água também passa pela cadeia alimentar dos microrganismos vegetais-animais e retorna à atmosfera.
Impacto humano no ciclo do nitrogênio A atividade humana tecnogênica perturba o equilíbrio natural do ciclo do nitrogênio. Ao arar a terra, a atividade dos microrganismos - fixadores de nitrogênio diminui quase 5 vezes, portanto o teor de nitrogênio no solo diminui, o que leva a uma diminuição da fertilidade do solo. Portanto, as pessoas introduzem no solo excesso de nitratos, que fazem parte dos fertilizantes minerais. Uma grande quantidade de óxidos de nitrogênio entra na atmosfera durante a combustão e processamento de gás, petróleo e carvão e cai na forma de chuva ácida. A restauração do ciclo natural do nitrogênio é possível reduzindo a produção de fertilizantes nitrogenados, reduzindo as emissões industriais de óxidos de nitrogênio na atmosfera, etc.
Ciclo do fósforo
Ao contrário dos ciclos da água, do carbono, do nitrogênio e do oxigênio, que são fechados, o ciclo do fósforo é aberto, porque o fósforo não forma compostos voláteis liberados na atmosfera. O fósforo está contido nas rochas, de onde entra nos ecossistemas durante a destruição natural das rochas ou quando fertilizantes fosfatados são aplicados nos campos. As plantas absorvem compostos inorgânicos de fósforo e os animais que se alimentam dessas plantas acumulam fósforo em seus tecidos. Após a decomposição dos cadáveres de animais e plantas, nem todo o fósforo está envolvido no ciclo. Parte dela é arrastada do solo para corpos d’água (rios, lagos, mares) e se deposita no fundo. O fósforo retorna à terra em pequenas quantidades com os peixes capturados pelo homem.
Impacto Humano no Ciclo do Fósforo A transferência de fósforo da terra para o oceano aumentou acentuadamente sob a influência humana. Quando as florestas são destruídas e os solos são arados, o volume do fluxo de água superficial aumenta e, além disso, entram fertilizantes fosfatados aplicados em rios e lagos a partir dos campos. Como as reservas de fósforo na terra são limitadas e o seu retorno do oceano é difícil, no futuro poderá haver falta de fósforo na agricultura, o que causará uma diminuição nos rendimentos (principalmente nas culturas de cereais).