Energiefluss in der Natur. Kreislauf von Materie und Energie in der Natur
Jedes Leben erfordert einen ständigen Fluss von Energie und Materie. Energie wird für die Durchführung grundlegender Lebensreaktionen aufgewendet, Materie wird für den Aufbau der Körper von Organismen verwendet. Die Existenz natürlicher Ökosysteme geht mit komplexen Prozessen des Stoff- und Energieaustauschs zwischen belebter und unbelebter Natur einher. Diese Prozesse sind sehr wichtig und hängen nicht nur von der Zusammensetzung der Lebensgemeinschaften ab, sondern auch von ihrer physischen Umgebung.
Energiefluss in der Gemeinschaft– Dies ist der Übergang von Organismen einer Ebene zur anderen in Form chemischer Bindungen organischer Verbindungen (Nahrung).
Der Fluss (Kreislauf) von Materie ist die Bewegung von Materie in Form chemischer Elemente und ihrer Verbindungen vom Produzenten zum Zersetzer und dann (durch chemische Reaktionen, die ohne Beteiligung lebender Organismen ablaufen) wieder zum Produzenten.
Die Zirkulation von Materie und der Fluss von Energie sind keine identischen Konzepte, obwohl häufig verschiedene Energieäquivalente (Kalorien, Kilokalorien, Joule) zur Messung der Bewegung von Materie verwendet werden. Dies erklärt sich teilweise dadurch, dass auf allen trophischen Ebenen, mit Ausnahme der ersten, die für das Leben von Organismen notwendige Energie in Form der Substanz der aufgenommenen Nahrung übertragen wird. Nur Pflanzen (Produzenten) können die Strahlungsenergie der Sonne direkt für ihre Lebenstätigkeit nutzen.
Eine genaue Messung einer in einem Ökosystem zirkulierenden Substanz kann durch die Berücksichtigung der Zirkulation einzelner chemischer Elemente erreicht werden, vor allem derjenigen, die das Hauptbaumaterial für das Zytoplasma pflanzlicher und tierischer Zellen darstellen.
Im Gegensatz zu Substanzen, die kontinuierlich durch verschiedene Blöcke des Ökosystems zirkulieren und immer wieder in den Kreislauf eintreten können, kann Energie im Körper nur einmal verwendet werden.
Nach den Gesetzen der Physik kann Energie von einer Form (z. B. Lichtenergie) in eine andere (z. B. die potentielle Energie von Nahrungsmitteln) übergehen, wird jedoch nie wieder erzeugt oder zerstört. Es kann keinen einzigen Prozess geben, der mit der Energieumwandlung verbunden ist, ohne dass ein Teil davon verloren geht. Bei seinen Umwandlungen wird eine gewisse Energiemenge in Form von Wärme abgegeben und geht somit verloren. Aus diesem Grund kann es nicht zu hundertprozentig wirksamen Umwandlungen beispielsweise von Nahrungsstoffen in körpereigene Stoffe kommen.
Die Existenz aller Ökosysteme hängt von einem konstanten Energiefluss ab, der für alle Organismen zur Aufrechterhaltung ihrer lebenswichtigen Funktionen und Selbstreproduktion notwendig ist.
Nur etwa die Hälfte des auf grüne Pflanzen fallenden Sonnenflusses wird von photosynthetischen Elementen absorbiert und nur ein kleiner Teil der absorbierten Energie (von 1/100 bis 1/20) wird in der für die Aktivität notwendigen Energie gespeichert von Pflanzengeweben.
Wenn man sich vom Primärproduzenten entfernt, nimmt die Geschwindigkeit des Energieflusses (d. h. die Energiemenge, ausgedrückt in Energieeinheiten, die von einer trophischen Ebene auf eine andere übertragen wird) stark ab.
Der Abfall der Energiemenge beim Übergang von einer trophischen Ebene zu einer höheren bestimmt die Anzahl dieser Ebenen selbst. Es wird geschätzt, dass jede trophische Ebene nur etwa 10 % (oder etwas mehr) der Energie der vorherigen Ebene erhält. Daher übersteigt die Gesamtzahl der trophischen Ebenen selten 3–4.
Das Verhältnis der lebenden Materie auf verschiedenen trophischen Ebenen folgt im Allgemeinen der gleichen Regel wie das Verhältnis der zugeführten Energie: Je höher die Ebene, desto geringer sind die Gesamtbiomasse und die Anzahl der sie konstituierenden Organismen.
Das Verhältnis der Anzahl verschiedener Organismengruppen gibt einen Eindruck von der Stabilität der Gemeinschaft, denn die Biomasse und die Anzahl einiger Populationen sind gleichzeitig ein Indikator für den Lebensraum für Organismen dieser und anderer Arten . Beispielsweise bestimmt die Anzahl der Bäume in einem Wald nicht nur den Gesamtvorrat an darin enthaltener Biomasse und Energie, sondern auch das Mikroklima sowie die Anzahl der Unterstände für viele Insekten und Vögel.
Zahlenpyramiden können umgekehrt werden. Dies geschieht, wenn die Reproduktionsrate der Beutepopulation hoch ist und selbst bei geringer Biomasse eine solche Population eine ausreichende Nahrungsquelle für Raubtiere sein kann, die über eine höhere Biomasse, aber eine niedrige Reproduktionsrate verfügen. Beispielsweise können viele Insekten auf einem Baum leben und sich davon ernähren (eine umgekehrte Populationspyramide). Die umgekehrte Biomassepyramide ist charakteristisch für aquatische Ökosysteme, in denen sich die Primärproduzenten (Phytoplanktonalgen) sehr schnell teilen und vermehren, während ihre Konsumenten (Zooplanktonkrebse) viel größer sind, aber einen langen Reproduktionszyklus haben.
Weide- und Schuttketten
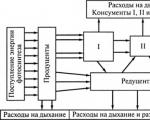
Energie kann auf viele verschiedene Arten durch eine Gemeinschaft fließen. Es repräsentiert die Nahrungskette aller Verbraucher(Consumer-System) mit der Hinzufügung von zwei weiteren Links: this tote organische Substanz Und Zersetzer der Nahrungskette(Reduziersystem).
Energiefluss von Pflanzen durch Pflanzenfresser(sie werden Weiden genannt), wird als Nahrungskette im Grünland bezeichnet.
Die Überreste der von ihnen verzehrten Organismen, die von den Verbrauchern nicht genutzt werden, ergänzen totes organisches Material. Es besteht aus Kot, der einen Teil unverdauter Nahrung enthält, sowie Tierkadavern, Vegetationsresten (Blätter, Zweige, Algen) und wird genannt Detritus.
Der Energiefluss, der aus toten organischen Stoffen entsteht und durch das Zersetzungssystem fließt, wird als schädliche Nahrungskette bezeichnet.
Neben den Ähnlichkeiten gibt es auch tiefgreifende Unterschiede in der Funktionsweise der Weide- und Detrital-Nahrungsketten. Es besteht darin, dass in Im Verbrauchersystem gehen Fäkalien und tote Organismen verloren und im reduktiven System– Nein.
Früher oder später wird die in toter organischer Substanz enthaltene Energie von Zersetzern vollständig genutzt und über die Atmung als Wärme abgeführt, auch wenn dafür ein mehrmaliges Durchlaufen eines Systems von Zersetzern erforderlich ist. Ausnahmen bilden lediglich Fälle, in denen die lokalen abiotischen Bedingungen für den Zersetzungsprozess sehr ungünstig sind (hohe Luftfeuchtigkeit, Permafrost). In diesen Fällen kommt es zu Ablagerungen unvollständig verarbeiteter, sehr energieintensiver Stoffe, die sich im Laufe der Zeit und unter geeigneten Bedingungen in brennbare organische Fossilien – Öl, Kohle, Torf – umwandeln.
Stoffkreislauf in einem Ökosystem
Die Integrität natürlicher Ökosysteme wird besonders deutlich, wenn man die in ihnen zirkulierenden Stoffströme betrachtet. Der Stoff kann in geschlossenen Kreisläufen (Kreisläufen) übertragen werden, wobei er immer wieder zwischen Organismen und der Umwelt zirkuliert.
Kreisförmige Bewegungen (durch Land, Luft, Wasser) chemischer Elemente (also Stoffe) werden biogeochemische Kreisläufe oder Zirkulationen genannt.
Konventionell werden die lebensnotwendigen Elemente und gelösten Salze genannt Nährstoffe(Leben geben) oder Nährstoffe. Bei den biogenen Elementen werden zwei Gruppen unterschieden: makrotrophe Stoffe und mikrotrophe Stoffe.
Makrotrophe Substanzen umfassen die Elemente, die die chemische Grundlage der Gewebe lebender Organismen bilden. Dazu gehören: Kohlenstoff, Wasserstoff, Sauerstoff, Stickstoff, Phosphor, Kalium, Kalzium, Magnesium, Schwefel.
Mikrotrophe Substanzen Dazu gehören Elemente und ihre Verbindungen, die ebenfalls für die Existenz lebender Systeme sehr wichtig sind, jedoch in äußerst geringen Mengen. Solche Substanzen werden oft genannt Mikroelemente. Dies sind Eisen, Mangan, Kupfer, Zink, Bor, Natrium, Molybdän, Chlor, Vanadium und Kobalt. Obwohl mikrotrophe Elemente in sehr geringen Mengen für Organismen lebenswichtig sind, kann ihr Mangel die Produktivität erheblich einschränken.
Die Zirkulation von Nährstoffen geht in der Regel mit deren chemischen Umwandlungen einher. Nitratstickstoff kann beispielsweise in Proteinstickstoff umgewandelt werden, dann in Harnstoff umgewandelt werden, in Ammoniak umgewandelt werden und unter dem Einfluss von Mikroorganismen wieder in die Nitratform synthetisiert werden. An den Prozessen der Denitrifikation und Stickstofffixierung sind verschiedene biologische und chemische Mechanismen beteiligt.
Die Nährstoffreserven sind unterschiedlich. Der Prozess der Sequestrierung einiger davon als lebende Biomasse verringert die Menge, die in der abiotischen Umwelt verbleibt. Und wenn Pflanzen und andere Organismen nicht irgendwann zersetzt würden, wäre der Nährstoffvorrat erschöpft und das Leben auf der Erde würde aufhören. Daraus können wir schließen die Aktivität von Heterotrophen, vor allem Organismen, die in Detritalketten funktionieren,– ein entscheidender Faktor für die Aufrechterhaltung des Nährstoffkreislaufs und der Produktbildung.
Schauen wir uns einige numerische Daten an, die das Ausmaß des Stofftransfers veranschaulichen, und wenden uns dabei dem biogeochemischen Kohlenstoffkreislauf zu. Die natürliche Kohlenstoffquelle, die Pflanzen für die Synthese organischer Stoffe verwenden, ist Kohlendioxid, das Teil der Atmosphäre ist oder in Wasser gelöst ist. Bei der Photosynthese wird Kohlendioxid (Kohlendioxid) in organische Substanz umgewandelt, die den Tieren als Nahrung dient. Atmung, Fermentation und Verbrennung von Kraftstoffen geben Kohlendioxid in die Atmosphäre zurück.
Die Kohlenstoffreserven in der Atmosphäre unseres Planeten werden auf 700 Milliarden Tonnen geschätzt, in der Hydrosphäre auf 50.000 Milliarden Tonnen. Nach jährlichen Berechnungen beträgt die Zunahme der Pflanzenmasse an Land und im Wasser durch Photosynthese 30 Milliarden Tonnen und Jeweils 150 Milliarden Tonnen. Der Kohlenstoffkreislauf dauert etwa 300 Jahre.
Ein weiteres Beispiel ist der Phosphorkreislauf. Die wichtigsten Phosphorreserven sind verschiedene Gesteine, die nach und nach (durch Zerstörung und Erosion) ihre Phosphate an terrestrische Ökosysteme abgeben. Phosphate werden von Pflanzen aufgenommen und von ihnen zur Synthese organischer Substanzen verwendet. Bei der Zersetzung von Tierkadavern durch Mikroorganismen werden Phosphate wieder in den Boden zurückgeführt und von Pflanzen erneut genutzt. Darüber hinaus wird ein Teil der Phosphate über Wasserläufe ins Meer verschleppt. Dadurch wird die Entwicklung des Phytoplanktons und aller davon abhängigen Nahrungsketten sichergestellt. Ein Teil des im Meerwasser enthaltenen Phosphors kann in Form von Guano an Land zurückkehren.
Ökosystem Biosphäre Energiekreislauf
Einführung
Konzept und Struktur von Ökosystemen
1 Konzept von Ökosystemen
2 Klassifizierung von Ökosystemen
3 Zonierung von Makroökosystemen
4 Ökosystemstruktur
Faktoren, die die Integrität von Ökosystemen gewährleisten
1 Stoffkreisläufe
2 Energiefluss in Ökosystemen
3 Dynamische Prozesse, die Integrität und Nachhaltigkeit in Ökosystemen gewährleisten
4 Die Biosphäre als globales Ökosystem, das die Integrität und Nachhaltigkeit von Ökosystemen gewährleistet
Abschluss
Einführung
Ein Ökosystem ist eine Ansammlung von Organismen und anorganischen Bestandteilen, in denen der Stoffkreislauf stattfinden kann. Laut N.F. Reimers (1990) ist ein Ökosystem jede Gemeinschaft von Lebewesen und deren Lebensraum, die zu einem einzigen funktionalen Ganzen vereint sind und auf der Grundlage von Interdependenzen und Ursache-Wirkungs-Beziehungen zwischen einzelnen Umweltkomponenten entstehen. A. Tansley (1935) schlug die folgende Beziehung vor:
Es gibt Mikroökosysteme, Mesoökosysteme, Makroökosysteme und die globale Biosphäre. Große terrestrische Ökosysteme werden Biome genannt. Ökosysteme sind nicht ungeordnet verstreut; im Gegenteil, sie sind in ziemlich regelmäßige Zonen gruppiert, sowohl horizontal (in der Breite) als auch vertikal (in der Höhe).
Vom Äquator bis zu den Polen ist eine gewisse Symmetrie in der Verteilung der Biome verschiedener Hemisphären erkennbar: tropische Regenwälder, Wüsten, Steppen, gemäßigte Wälder, Nadelwälder, Taiga.
Jedes Ökosystem besteht aus zwei Hauptkomponenten: Organismen und Faktoren aus ihrer unbelebten Umgebung. Die Gesamtheit der Organismen (Pflanzen, Tiere, Mikroben) wird als Biota eines Ökosystems bezeichnet. Aus Sicht der trophischen Struktur (von griechisch trophe – Ernährung) lässt sich das Ökosystem in zwei Ebenen einteilen: obere und untere.
Als Teil eines Ökosystems werden folgende Komponenten unterschieden: anorganische, organische Verbindungen, Luft-, Wasser- und Substratumgebungen, Produzenten, autotrophe Organismen, Konsumenten oder Phagotrophen, Zersetzer und Detritivoren.
Sonnenenergie auf der Erde verursacht zwei Stoffkreisläufe: große oder geologische und kleine, biologische (biotische).
In einem Ökosystem gibt es Stoffkreisläufe. Am meisten untersucht sind: die Kreisläufe von Kohlenstoff, Sauerstoff, Stickstoff, Phosphor, Schwefel usw.
Die Aufrechterhaltung der lebenswichtigen Aktivität von Organismen und der Stoffzirkulation in Ökosystemen, also die Existenz von Ökosystemen, hängt vom ständigen Zufluss von Sonnenenergie ab.
In Ökosystemen kommt es ständig zu Veränderungen im Zustand und in der Lebensaktivität ihrer Mitglieder sowie im Verhältnis der Populationen. Die vielfältigen Veränderungen, die in jeder Gemeinschaft stattfinden, lassen sich in zwei Haupttypen einteilen: zyklische und progressive.
Ein globales Ökosystem ist eine Biosphäre, die durch die Integrität und Stabilität der Ökosysteme gekennzeichnet ist.
1. Konzept und Struktur von Ökosystemen
1 Konzept von Ökosystemen
Lebende Organismen und ihre unbelebte (abiotische) Umwelt sind untrennbar miteinander verbunden und stehen in ständiger Wechselwirkung. Jede Einheit (Biosystem) eines Ökosystems ist ein Ökosystem. Ein Ökosystem oder Ökosystem ist die grundlegende Funktionseinheit der Ökologie, da es Organismen und die unbelebte Umwelt umfasst – Komponenten, die sich gegenseitig in ihren Eigenschaften und den notwendigen Bedingungen für die Aufrechterhaltung des Lebens in der auf der Erde existierenden Form beeinflussen. Der Begriff „Ökosystem“ wurde erstmals 1935 vom englischen Ökologen A. Tansley vorgeschlagen. Derzeit wird häufig die folgende Definition eines Ökosystems verwendet. Ein Ökosystem ist eine Ansammlung von Organismen und anorganischen Bestandteilen, in denen der Stoffkreislauf stattfinden kann. Laut N.F. Reimers (1990) ist ein Ökosystem jede Gemeinschaft von Lebewesen und deren Lebensraum, die zu einem einzigen funktionalen Ganzen vereint sind und auf der Grundlage von Interdependenzen und Ursache-Wirkungs-Beziehungen zwischen einzelnen Umweltkomponenten entstehen. Hervorzuheben ist, dass die Kombination einer bestimmten physikalisch-chemischen Umgebung (Biotop) mit einer Gemeinschaft lebender Organismen (Biozönose) ein Ökosystem bildet. A. Tansley (1935) schlug die folgende Beziehung vor:
Ökosystem = Biotop + Biozönose
Nach der Definition von V.N. Sukachev, Biogeozänose ist „eine Reihe homogener natürlicher Phänomene (Atmosphäre, Gestein, Boden und hydrologische Bedingungen) über einen bestimmten Bereich der Erdoberfläche, die ihre eigene besondere Spezifität der Wechselwirkungen dieser Komponenten, aus denen sie besteht, und eine bestimmte Art von.“ Austausch von Materie und Energie untereinander und anderen Naturphänomenen und stellen eine in sich widersprüchliche dialektische Einheit dar, die sich in ständiger Bewegung und Entwicklung befindet.
Neben den Konzepten des Ökosystems A. Tensley und der Biogeozänose V.N. Sukachev formulierte die Regel von F. Evans (1956), der vorschlug, den Begriff „Ökosystem“ absolut „dimensionslos“ zu verwenden, um jedes supraorganismische lebende System zu bezeichnen, das mit der Umwelt interagiert. Viele Autoren gaben dem Begriff „Ökosystem“ jedoch die Bedeutung von Biogeozänose, d.h. elementares Ökosystem und gleichzeitig höher in der Hierarchie der suprabiogeozänotischen Formationen bis hin zum Biosphärenökosystem.
2 Klassifizierung von Ökosystemen
Die auf der Erde existierenden Ökosysteme sind vielfältig. Es gibt Mikroökosysteme (zum Beispiel der Stamm eines verrottenden Baumes), Mesoökosysteme (Wald, Teich usw.), Makroökosysteme (Kontinent, Ozean usw.) und die globale Biosphäre.
Große terrestrische Ökosysteme werden Biome genannt. Jedes Biom enthält eine Reihe kleinerer, miteinander verbundener Ökosysteme. Es gibt verschiedene Klassifizierungen von Ökosystemen:
Immergrüner tropischer Regenwald
Wüste: Gras- und Strauch-Chaparral – Gebiete mit regnerischen Wintern und trockenen Sommern
Tropisches Graslenz und Savannah
Gemäßigte Steppe
Gemäßigter Laubwald
Boreale Nadelwälder
Tundra: arktisch und alpin
Arten von Süßwasserökosystemen:
Band (stille Gewässer): Seen, Teiche usw.
Lotisch (fließende Gewässer): Flüsse, Bäche usw.
Feuchtgebiete: Sümpfe und sumpfige Wälder
Arten mariner Ökosysteme
Offener Ozean (pelagisch)
Kontinentalschelfgewässer (Küstengewässer)
Auftriebsgebiete (fruchtbare Gebiete mit produktiver Fischerei)
Flussmündungen (Küstenbuchten, Meerengen, Flussmündungen, Salzwiesen usw.). Dabei zeichnen sich terrestrische Biome durch natürliche bzw. ursprüngliche Vegetationsmerkmale und aquatische Ökosystemtypen durch geologische und physikalische Merkmale aus. Die aufgeführten Haupttypen von Ökosystemen stellen die Umgebung dar, in der sich die menschliche Zivilisation entwickelt hat, und stellen die wichtigsten Lebensgemeinschaften dar, die das Leben auf der Erde unterstützen.
3 Zonierung von Makroökosystemen
Die Untersuchung der geografischen Verteilung von Ökosystemen kann nur auf der Ebene großer ökologischer Einheiten – Makroökosystemen – durchgeführt werden, die auf kontinentaler Ebene betrachtet werden. Ökosysteme sind nicht ungeordnet verstreut; im Gegenteil, sie sind in ziemlich regelmäßige Zonen gruppiert, sowohl horizontal (in der Breite) als auch vertikal (in der Höhe). Dies wird durch das periodische Gesetz der geografischen Zonierung von A.A. bestätigt. Grigorieva - M.I. Budyko: Mit der Veränderung der physikalisch-geografischen Zonen der Erde wiederholen sich regelmäßig ähnliche Landschaftszonen und einige ihrer allgemeinen Eigenschaften. Dies wurde auch bei der Betrachtung der Boden-Luft-Umgebung des Lebens diskutiert. Die gesetzlich festgelegte Periodizität zeigt sich darin, dass die Werte des Trockenheitsindex in verschiedenen Zonen von 0 bis 4-5 variieren, dreimal zwischen den Polen und dem Äquator liegen sie nahe bei Eins. Diese Werte entsprechen der höchsten biologischen Produktivität von Landschaften.
Die periodische Wiederholung von Eigenschaften in einer Reihe von Systemen einer hierarchischen Ebene ist wahrscheinlich ein allgemeines Gesetz des Universums, formuliert als Gesetz der Periodizität in der Struktur systemischer Aggregate oder als systemperiodisches Gesetz – spezifische natürliche Systeme einer Ebene ( Unterebene) der Organisation stellen eine periodische oder sich wiederholende Reihe morphologisch ähnlicher Strukturen innerhalb der oberen und unteren Raum-Zeit-Grenzen des Systems dar, jenseits derer die Existenz von Systemen auf einer bestimmten Ebene unmöglich wird. Sie geraten in einen instabilen Zustand oder verwandeln sich in eine andere Systemstruktur, einschließlich einer anderen Organisationsebene.
Temperatur und Niederschlag bestimmen die Lage der wichtigsten terrestrischen Biome auf der Erdoberfläche. Das Temperatur- und Niederschlagsmuster in einem bestimmten Gebiet über einen längeren Zeitraum hinweg bezeichnen wir als Klima. Das Klima ist in verschiedenen Regionen der Erde unterschiedlich. Der jährliche Niederschlag variiert zwischen 0 und 2500 mm oder mehr. Temperatur- und Niederschlagsregime wirken auf sehr unterschiedliche Weise zusammen.
Die Besonderheiten der klimatischen Bedingungen bestimmen wiederum die Entwicklung eines bestimmten Bioms.
Vom Äquator bis zu den Polen ist eine gewisse Symmetrie in der Verteilung der Biome verschiedener Hemisphären erkennbar:
Tropische Regenwälder (nördliches Südamerika, Mittelamerika, westliche und zentrale Teile Äquatorialafrikas, Südostasien, Küstengebiete im Nordwesten Australiens, Inseln des Indischen und Pazifischen Ozeans). Das Klima ist ohne Wechsel der Jahreszeiten (Äquatornähe), die durchschnittliche Jahrestemperatur liegt über 17°C (normalerweise 28°C), der durchschnittliche jährliche Niederschlag übersteigt 2400 mm.
Vegetation: Wälder dominieren. Einige Baumarten erreichen eine Höhe von bis zu 60 m. An ihren Stämmen und Ästen wachsen epiphytische Pflanzen, deren Wurzeln nicht bis zum Boden reichen, und verholzende Ranken, die im Boden Wurzeln schlagen und die Bäume bis zu ihren Wipfeln erklimmen. All dies bildet ein dichtes Blätterdach.
Fauna: Die Artenzusammensetzung ist reicher als in allen anderen Biomen zusammen. Besonders zahlreich sind Amphibien, Reptilien und Vögel (Frösche, Eidechsen, Schlangen, Papageien), Affen und andere Kleinsäuger, exotische Insekten mit leuchtenden Farben sowie bunte Fische in Stauseen.
Merkmale: Böden sind oft dünn und arm, die meisten Nährstoffe sind in der Oberflächenbiomasse der Wurzelvegetation enthalten.
2. Savannen (subäquatoriales Afrika, Südamerika, ein bedeutender Teil Südindiens). Das Klima ist die meiste Zeit des Jahres trocken und heiß. Starke Regenfälle während der Regenzeit. Temperatur: durchschnittlicher Jahreshöchstwert. Die Niederschlagsmenge beträgt 750–1650 mm/Jahr, hauptsächlich während der Regenzeit. Vegetation - Bluegrass-Pflanzen (Getreidepflanzen) mit seltenen Laubbäumen. Fauna: große pflanzenfressende Säugetiere wie Antilopen, Zebras, Giraffen, Nashörner, Raubtiere – Löwen, Leoparden, Geparden.
Wüsten (einige Gebiete Afrikas, zum Beispiel die Sahara; der Nahe Osten und Zentralasien, das Große Becken und der Südwesten der Vereinigten Staaten und Nordmexiko usw.). Das Klima ist sehr trocken. Temperatur – heiße Tage und kalte Nächte. Die Niederschlagsmenge beträgt weniger als 250 mm/Jahr. Vegetation: spärliche Sträucher, oft dornig, manchmal Kakteen und niedrige Gräser, die nach seltenen Regenfällen den Boden schnell mit einem blühenden Teppich bedecken. Pflanzen verfügen über ausgedehnte Oberflächenwurzelsysteme, die Feuchtigkeit aus seltenen Niederschlägen abfangen, sowie über Pfahlwurzeln, die bis zum Grundwasserspiegel (30 m und tiefer) in den Boden eindringen. Fauna: verschiedene Nagetiere (Känguru-Ratte usw.), Kröten, Eidechsen, Schlangen und andere Reptilien, Eulen, Adler, Geier, kleine Vögel und Insekten in großen Mengen.
4. Steppen (Zentrum Nordamerikas, Russland, bestimmte Gebiete Afrikas und Australiens, südöstlich von Südamerika). Das Klima ist saisonal. Temperaturen: Die Temperaturen im Sommer reichen von mäßig warm bis heiß, im Winter liegen die Temperaturen unter 0 °C. Niederschlag - 750-2000 mm/Jahr. Vegetation: dominiert von Bluegrass (Getreide) mit einer Höhe von bis zu 2 m und mehr in einigen Prärien Nordamerikas oder bis zu 50 cm, beispielsweise in den russischen Steppen, mit vereinzelten Bäumen und Sträuchern in Feuchtgebieten. Fauna: große pflanzenfressende Säugetiere – Bisons, Gabelbockantilopen (Nordamerika), Wildpferde (Eurasien), Kängurus (Australien), Giraffen, Zebras, Breitmaulnashörner, Antilopen (Afrika); Zu den Raubtieren zählen Kojoten, Löwen, Leoparden, Geparden, Hyänen, eine Vielzahl von Vögeln und kleine grabende Säugetiere wie Kaninchen, Erdhörnchen und Erdferkel.
5. Wälder der gemäßigten Zone (Westeuropa, Ostasien, Osten der USA). Klima – saisonal mit Wintertemperaturen unter 0°C 0C. Niederschlag - 750-2000 mm/Jahr. Vegetation: dominiert von Wäldern mit bis zu 35–45 m hohen Laubbäumen (Eiche, Hickory, Ahorn), strauchigem Unterholz, Moosen und Flechten. Fauna: Säugetiere (Weißwedelhirsch, Stachelschwein, Waschbär, Opossum, Eichhörnchen, Kaninchen, Spitzmäuse), Vögel (Trällerer, Spechte, Amseln, Eulen, Falken), Schlangen, Frösche, Salamander, Fische (Forelle, Barsch, Wels usw . ), reichlich vorhandene Bodenmikrofauna (Abb. 12.3). Die Biota ist an das saisonale Klima angepasst: Winterschlaf, Migration, Ruhe in den Wintermonaten. 6. Nadelwälder, Taiga (nördliche Regionen Nordamerikas, Europas und Asiens). Das Klima ist von langen und kalten Wintern geprägt, wobei einige Niederschläge in Form von Schnee fallen. Vegetation: Es überwiegen immergrüne Nadelwälder, hauptsächlich Fichten, Kiefern und Tannen. Fauna: große pflanzenfressende Huftiere (Maultierhirsche, Rentiere), kleine pflanzenfressende Säugetiere (Hasen, Eichhörnchen, Nagetiere), Wölfe, Luchse, Füchse, Schwarzbären, Grizzlybären, Vielfraße, Nerze und andere Raubtiere, im kurzen Sommer zahlreiche blutsaugende Insekten Zeit. Das Klima ist sehr kalt mit Polartagen und Polarnächten. Die durchschnittliche Jahrestemperatur liegt unter - 5°C. In wenigen Wochen des kurzen Sommers taut der Boden höchstens einen Meter tief auf. Die Niederschlagsmenge beträgt weniger als 250 mm/Jahr. Vegetation: dominiert von langsam wachsenden Flechten, Moosen, Gräsern und Seggen sowie Zwergsträuchern. Fauna: große pflanzenfressende Huftiere (Rentiere, Moschusochsen), kleine grabende Säugetiere (ganzjährig zum Beispiel Lemminge), Raubtiere, die im Winter eine weiße Tarnfarbe annehmen (Polarfuchs, Luchs, Hermelin, Schneeeule). Im kurzen Sommer nisten in der Tundra zahlreiche Zugvögel, darunter besonders viele Wasservögel, die sich von den hier reichlich vorkommenden Insekten und wirbellosen Süßwassertieren ernähren. Sehr deutlich ist auch die vertikale Höhenzonierung von Landökosystemen, insbesondere an Orten mit ausgeprägtem Relief. Die Luftfeuchtigkeit ist der Hauptfaktor, der die Art des Bioms bestimmt. Bei ausreichend großen Niederschlagsmengen entwickelt sich Waldvegetation. Die Temperatur bestimmt die Art des Waldes. In den Steppen- und Wüstenbiomen ist die Situation genau die gleiche. Veränderungen der Vegetationstypen in kalten Regionen treten mit geringeren jährlichen Niederschlagsmengen auf, da bei niedrigen Temperaturen weniger Wasser durch Verdunstung verloren geht. Die Temperatur wird nur bei sehr kalten Bedingungen mit Permafrost zu einem wichtigen Faktor. Somit hängt die Zusammensetzung von Ökosystemen maßgeblich von ihrem funktionalen „Zweck“ ab und umgekehrt. Laut N.F. Reimers (1994) spiegelt sich dies im Prinzip der ökologischen Komplementarität (Komplementarität) wider: Kein funktionaler Teil des Ökosystems (ökologische Komponente, Element etc.) kann ohne andere funktional komplementäre Teile existieren. Ihm nahe und erweiternd steht das Prinzip der ökologischen Kongruenz (Korrespondenz): Die lebenden Bestandteile des Ökosystems entwickeln sich funktionell ergänzend und entwickeln dafür entsprechende Anpassungen, abgestimmt auf die Bedingungen der abiotischen Umwelt, die weitgehend von denselben Organismen (Bioklima etc.) verändert wird, d.h. Es gibt eine doppelte Reihe von Korrespondenzen – zwischen Organismen und ihrem Lebensraum – äußerlich und durch die Volkszählung geschaffen. 4 Ökosystemstruktur Aus Sicht der trophischen Struktur (von griech. trophe – Ernährung) lässt sich das Ökosystem in zwei Ebenen einteilen: Die obere - autotrophe (selbsternährende) Stufe oder „grüner Gürtel“, einschließlich Pflanzen oder deren Teile, die Chlorophyll enthalten, wo die Fixierung von Zählenergie und die Verwendung einfacher anorganischer Verbindungen vorherrschen. Die untere ist die heterotrophe (von anderen gespeiste) Schicht oder der „braune Gürtel“ aus Böden und Sedimenten, verrottender Materie, Wurzeln usw., in der die Nutzung, Umwandlung und Zersetzung komplexer Verbindungen vorherrscht. Aus biologischer Sicht werden folgende Komponenten als Teil eines Ökosystems unterschieden: In einem Ökosystem sind Nahrungs- und Energiezusammenhänge zwischen Kategorien immer eindeutig und gehen in die Richtung: Autotrophe - Heterotrophe oder in vollständigerer Form; Autotrophe -> Verbraucher -> Zersetzer (Destruktoren). Die wichtigste Energiequelle für Ökosysteme ist die Sonne. Der Energiefluss (nach T.A. Akimova, V.V. Khaskin, 1994), der von der Sonne zum Planeten Erde gesendet wird, übersteigt 20 Millionen EJ pro Jahr. Nur ein Viertel dieser Strömung erreicht die Grenze der Atmosphäre. Davon werden etwa 70 % reflektiert, von der Atmosphäre absorbiert und in Form langwelliger Infrarotstrahlung abgegeben. Die auf die Erdoberfläche fallende Sonnenstrahlung beträgt 1,54 Millionen EJ pro Jahr. Diese riesige Energiemenge entspricht dem 5000-fachen der gesamten Energie der Menschheit am Ende des 20. Jahrhunderts und dem 5,5-fachen der Energie aller verfügbaren fossilen Brennstoffressourcen organischen Ursprungs, die sich über mindestens 100 Millionen Jahre angesammelt haben. Der Großteil der Sonnenenergie, die die Planetenoberfläche erreicht, wird direkt in Wärme umgewandelt und erwärmt so Wasser oder Boden, was wiederum die Luft erwärmt. Diese Wärme dient als treibende Kraft hinter dem Wasserkreislauf, den Luft- und Meeresströmungen, die das Wetter bestimmen, und wird nach und nach in den Weltraum abgegeben, wo sie verloren geht. Um den Platz von Ökosystemen in diesem natürlichen Energiefluss zu bestimmen, ist es wichtig zu erkennen, dass sie, egal wie umfangreich und komplex sie sind, nur einen kleinen Teil davon nutzen. Dies impliziert eines der Grundprinzipien der Funktionsweise von Ökosystemen: Sie existieren aufgrund der umweltfreundlichen und nahezu ewigen Sonnenenergie, deren Menge relativ konstant und reichlich vorhanden ist. Lassen Sie uns die einzelnen aufgeführten Eigenschaften der Solarenergie genauer erläutern: 2. Faktoren, die die Integrität von Ökosystemen gewährleisten 1 Stoffkreisläufe Beide Kreisläufe sind miteinander verbunden und stellen gewissermaßen einen einzigen Prozess dar. Es wird geschätzt, dass der gesamte in der Atmosphäre enthaltene Sauerstoff in 2000 Jahren durch Organismen zirkuliert (bei der Atmung kombiniert und bei der Photosynthese freigesetzt wird), das Kohlendioxid in der Atmosphäre in 300 Jahren in die entgegengesetzte Richtung zirkuliert und das gesamte Wasser auf der Erde ebenfalls zirkuliert in 2.000.000 Jahren durch Photosynthese und Atmung zersetzt und neu gebildet. Das Zusammenspiel abiotischer Faktoren und lebender Organismen des Ökosystems geht mit einem kontinuierlichen Stoffkreislauf zwischen Biotop und Biozönose in Form abwechselnder organischer und mineralischer Verbindungen einher. Der Austausch chemischer Elemente zwischen lebenden Organismen und der anorganischen Umgebung, der in verschiedenen Phasen innerhalb eines Ökosystems abläuft, wird als biogeochemischer Kreislauf oder biogeochemischer Kreislauf bezeichnet. Der Wasserkreislauf. Der bedeutendste Kreislauf auf der Erde hinsichtlich der übertragenen Massen und des Energieverbrauchs ist der planetarische Wasserkreislauf – der Wasserkreislauf. Jede Sekunde sind daran 16,5 Millionen m3 Wasser beteiligt und dafür werden mehr als 40 Milliarden MW Sonnenenergie aufgewendet (laut T.A. Akimova V.V. Haskin, (1994)). Bei diesem Kreislauf handelt es sich jedoch nicht nur um die Übertragung von Wassermassen. Dies sind Phasenumwandlungen, die Bildung von Lösungen und Suspensionen, Fällung, Kristallisation, Photosyntheseprozesse sowie verschiedene chemische Reaktionen. Das Leben ist in dieser Umgebung entstanden und lebt weiter. Wasser ist das Grundelement, das zum Leben notwendig ist. Quantitativ ist es der häufigste anorganische Bestandteil lebender Materie. Beim Menschen macht Wasser 63 % des Körpergewichts aus, bei Pilzen 80 %, bei Pflanzen 80–90 % und bei einigen Quallen 98 % Wie wir etwas später sehen werden, ist Wasser am biologischen Kreislauf beteiligt und dient als Quelle für Wasserstoff und Sauerstoff, wobei es nur einen kleinen Teil seines Gesamtvolumens ausmacht. Im flüssigen, festen und dampfförmigen Zustand ist Wasser in allen drei Hauptbestandteilen der Biosphäre vorhanden: der Atmosphäre, der Hydrosphäre und der Lithosphäre. Alle Gewässer verbindet das gemeinsame Konzept der „Hydrosphäre“. Die Bestandteile der Hydrosphäre sind durch ständigen Austausch und Wechselwirkung miteinander verbunden. Wasser, das sich ständig von einem Zustand in einen anderen bewegt, vollzieht kleine und große Kreisläufe. Die Verdunstung von Wasser von der Meeresoberfläche, die Kondensation von Wasserdampf in der Atmosphäre und der Niederschlag an der Meeresoberfläche bilden einen kleinen Kreislauf. Wenn Wasserdampf durch Luftströmungen an Land transportiert wird, wird der Kreislauf viel komplexer. In diesem Fall verdunstet ein Teil des Niederschlags und gelangt wieder in die Atmosphäre, der andere Teil speist Flüsse und Stauseen, kehrt aber letztendlich über Fluss- und Untergrundabflüsse in den Ozean zurück, wodurch der große Kreislauf geschlossen wird. Biotischer (biologischer) Zyklus. Der biotische (biologische) Kreislauf bezeichnet den Stoffkreislauf zwischen Boden, Pflanzen, Tieren und Mikroorganismen. Der biotische (biologische) Kreislauf ist der Fluss chemischer Elemente aus Boden, Wasser und Atmosphäre in lebende Organismen, die Umwandlung einströmender Elemente in neue komplexe Verbindungen und deren Rückkehr in den Lebensprozess mit dem jährlichen Rückgang eines Teils der organischen Substanz Materie oder mit vollständig toten Organismen, die in der Zusammensetzung des Ökosystems enthalten sind. Nun stellen wir den biotischen Zyklus in zyklischer Form dar. Der zentrale biotische Zyklus (nach T.A. Akimova, V.V., Khaskhin) bestand aus primitiven einzelligen Produzenten (P) und Zersetzer-Destruktoren (D). Mikroorganismen sind in der Lage, sich schnell zu vermehren und sich an unterschiedliche Bedingungen anzupassen, indem sie beispielsweise alle Arten von Substraten – Kohlenstoffquellen – in ihrer Ernährung nutzen. Höhere Organismen verfügen nicht über solche Fähigkeiten. In vollständigen Ökosystemen können sie in Form einer Struktur auf der Grundlage von Mikroorganismen existieren. Zunächst entwickeln sich mehrzellige Pflanzen (P) – höhere Produzenten. Zusammen mit einzelligen Organismen erzeugen sie durch den Prozess der Photosynthese organisches Material und nutzen dabei die Energie der Sonnenstrahlung. Anschließend werden Primärkonsumenten verbunden – pflanzenfressende Tiere (T) und dann fleischfressende Konsumenten. Wir haben den biotischen Kreislauf von Land untersucht. Dies gilt uneingeschränkt für den biotischen Kreislauf aquatischer Ökosysteme, beispielsweise des Ozeans. Alle Organismen nehmen einen bestimmten Platz im biotischen Kreislauf ein und erfüllen ihre Aufgabe, die Zweige des Energieflusses, die sie empfangen, umzuwandeln und Biomasse zu übertragen. Ein System einzelliger Zersetzer (Destruktoren) vereint alle, entpersonalisiert ihre Substanzen und schließt den allgemeinen Kreis. Sie geben der abiotischen Umgebung der Biosphäre alle Elemente zurück, die für immer neue Revolutionen notwendig sind. Die wichtigsten Merkmale des biotischen Zyklus sollten hervorgehoben werden. Die Photosynthese ist ein leistungsstarker natürlicher Prozess, der jedes Jahr große Mengen an Biosphärenmaterial in den Kreislauf einbezieht und deren hohes Sauerstoffpotenzial bestimmt. Durch Kohlendioxid und Wasser wird organisches Material synthetisiert und freier Sauerstoff freigesetzt. Die direkten Produkte der Photosynthese sind verschiedene organische Verbindungen, und im Allgemeinen ist der Prozess der Photosynthese recht komplex. Neben der Photosynthese unter Beteiligung von Sauerstoff, der sogenannten sauerstoffhaltigen Photosynthese, sollten wir uns auch auf die sauerstofffreie Photosynthese bzw. Chemosynthese konzentrieren. Zu den chemosynthetischen Organismen gehören Nitrifikatoren, Carboxydobakterien, Schwefelbakterien, thionische Eisenbakterien und Wasserstoffbakterien. Sie sind nach ihren Oxidationssubstraten benannt, bei denen es sich um NH3, NO2, CO, H2S, S, Fe2+, H2 handeln kann. Einige Arten sind obligat chemolithoautotroph, andere sind fakultativ. Zu letzteren zählen Carboxydobakterien und Wasserstoffbakterien. Chemosynthese ist charakteristisch für hydrothermale Quellen in der Tiefsee. Die Photosynthese findet bis auf wenige Ausnahmen auf der gesamten Erdoberfläche statt, erzeugt einen enormen geochemischen Effekt und kann als Menge der gesamten Kohlenstoffmasse ausgedrückt werden, die jährlich am Aufbau organischer – lebender Materie der gesamten Biosphäre beteiligt ist. Der allgemeine Stoffkreislauf, der mit dem Aufbau organischer Materie durch Photosynthese verbunden ist, umfasst auch chemische Elemente wie N, P, S sowie Metalle – K, Ca, Mg, Na, Al. Wenn ein Organismus stirbt, findet der umgekehrte Prozess statt – die Zersetzung organischer Stoffe durch Oxidation, Zerfall usw. unter Bildung endgültiger Zersetzungsprodukte. In der Biosphäre der Erde führt dieser Prozess dazu, dass die Menge der Biomasse lebender Materie tendenziell einigermaßen konstant ist. Biomasse der Ökosphäre (2 10|2t) ist sieben Größenordnungen kleiner als die Masse der Erdkruste (2,10|9t). Pflanzen der Erde produzieren jährlich organisches Material, das 1,6,10 % oder 8 % der Biomasse der Ökosphäre entspricht. Destruktoren, die weniger als 1 % der gesamten Biomasse der Organismen des Planeten ausmachen, verarbeiten eine Masse organischer Substanz, die 10 % entspricht Mal größer als ihre eigene Biomasse. Im Durchschnitt beträgt der Zeitraum der Biomasseerneuerung 12,5 Jahre. Nehmen wir also an, dass die Masse der lebenden Materie und die Produktivität der Biosphäre vom Kambrium bis zur Gegenwart (530 Millionen Jahre) gleich waren Die Gesamtmenge an organischer Substanz, die den globalen Biozyklus durchlaufen hat und vom Leben auf dem Planeten genutzt wurde, beträgt 2,10 " 2-5,ZL08/12,5=8,5L0|9t, was dem Vierfachen der Masse der Erdkruste entspricht. Zu diesen Berechnungen N.S. Pechurkin (1988) schrieb: „Wir können sagen, dass die Atome, aus denen unser Körper besteht, in alten Bakterien, in Dinosauriern und in Mammuts waren.“ Gesetz der biogenen Migration von Atomen V.I. Wernadskij stellt fest: „Die Migration chemischer Elemente auf der Erdoberfläche und in der Biosphäre insgesamt erfolgt entweder unter direkter Beteiligung lebender Materie (biogene Migration) oder sie findet in einer Umgebung statt, deren geochemische Eigenschaften (O2, CO2, H2, usw.) sind determinierte lebende Materie, sowohl das, was derzeit in der Biosphäre lebt, als auch das, was im Laufe der Erdgeschichte auf die Erde eingewirkt hat.“ IN UND. Wernadskij 1928-1930 In seinen tiefgreifenden Verallgemeinerungen zu Prozessen in der Biosphäre gab er eine Vorstellung von den fünf wichtigsten biogeochemischen Funktionen lebender Materie. Die erste Funktion ist Gas. Die zweite Funktion ist die Konzentration. Die dritte Funktion ist Redox. Die vierte Funktion ist biochemischer Natur. Die fünfte Funktion ist die biogeochemische Aktivität des Menschen, die eine immer größere Menge an Materie in der Erdkruste für den Bedarf von Industrie, Verkehr und Landwirtschaft abdeckt. Der biologische Kreislauf variiert in verschiedenen Naturzonen und wird anhand einer Reihe von Indikatoren klassifiziert: Pflanzenbiomasse, Einstreu, Einstreu, Menge der in der Biomasse fixierten Elemente usw. In der Waldzone ist die Gesamtbiomasse am höchsten und der Anteil unterirdischer Organe im Wald am niedrigsten. Dies wird durch den Intensitätsindex des biologischen Zyklus bestätigt – das Verhältnis der Masse der Einstreu zu dem Teil der Einstreu, aus dem sie besteht. Kohlenstoffzyklus. Von allen biogeochemischen Kreisläufen ist der Kohlenstoffkreislauf zweifellos der intensivste. Kohlenstoff zirkuliert mit hoher Geschwindigkeit zwischen verschiedenen anorganischen Medien und durch Nahrungsnetze innerhalb von Gemeinschaften lebender Organismen. CO und CO2 spielen eine gewisse Rolle im Kohlenstoffkreislauf. In der Biosphäre der Erde wird Kohlenstoff häufig durch die mobilste Form von CO2 repräsentiert. Die Quelle des primären Kohlendioxids in der Biosphäre ist vulkanische Aktivität, die mit der säkularen Entgasung des Erdmantels und der unteren Horizonte der Erdkruste verbunden ist. Die Migration von CO2 in der Biosphäre erfolgt auf zwei Arten. Der erste Weg besteht darin, es während der Photosynthese unter Bildung von Glukose und anderen organischen Substanzen aufzunehmen, aus denen alle Pflanzengewebe aufgebaut sind. Anschließend werden sie über Nahrungsketten transportiert und bilden das Gewebe aller anderen Lebewesen im Ökosystem. Beim Absterben von Pflanzen und Tieren an der Oberfläche kommt es zur Oxidation organischer Stoffe unter Bildung von CO2. Auch bei der Verbrennung organischer Stoffe gelangen Kohlenstoffatome wieder in die Atmosphäre. Ein wichtiges und interessantes Merkmal des Kohlenstoffkreislaufs besteht darin, dass in fernen geologischen Epochen, vor Hunderten von Millionen Jahren, ein erheblicher Teil der bei den Prozessen der Photosynthese entstandenen organischen Substanz weder von Verbrauchern noch von Zersetzern genutzt wurde, sondern sich in der Lithosphäre ansammelte in Form von fossilen Brennstoffen: Öl, Kohle, Ölschiefer, Torf usw. Diese fossilen Brennstoffe werden in großen Mengen abgebaut, um den Energiebedarf unserer Industriegesellschaft zu decken. Indem wir es verbrennen, schließen wir gewissermaßen den Kohlenstoffkreislauf. Beim zweiten Weg erfolgt die Kohlenstoffmigration durch die Schaffung eines Karbonatsystems in verschiedenen Reservoirs, in dem sich CO2 in H2CO3, HCO, CO3 umwandelt. Mit Hilfe von in Wasser gelöstem Calcium (oder Magnesium) werden Carbonate (CaCO3) auf biogenen und abiogenen Wegen ausgefällt. Es bilden sich dicke Kalksteinschichten. Laut A.B. Laut Ronov beträgt das Verhältnis von vergrabenem Kohlenstoff in Photosyntheseprodukten zu Kohlenstoff in Karbonatgesteinen 1:4. Neben dem großen Kohlenstoffkreislauf gibt es auch eine Reihe kleiner Kohlenstoffkreisläufe auf der Landoberfläche und im Ozean. Im Allgemeinen bleibt der Kohlenstoffgehalt in biogeochemischen Reservoirs: Biosphäre (Biomasse + Boden und Detritus), Sedimentgestein, Atmosphäre und Hydrosphäre ohne anthropogene Eingriffe mit einem hohen Maß an Konstanz erhalten (nach T.A. Akimova, V.V. Haskin (1994)). ). Der ständige Kohlenstoffaustausch einerseits zwischen der Biosphäre und andererseits zwischen Atmosphäre und Hydrosphäre wird durch die Gasfunktion der lebenden Materie – die Prozesse der Photosynthese, Atmung und Zerstörung – bestimmt und beträgt etwa 6-1010 Tonnen/Jahr. Es gibt einen Kohlenstofffluss in die Atmosphäre und Hydrosphäre und bei vulkanischer Aktivität durchschnittlich 4,5 106 t/Jahr. Die Gesamtmasse an Kohlenstoff in fossilen Brennstoffen (Öl, Gas, Kohle usw.) wird auf 3,2*1015 Tonnen geschätzt, was einer durchschnittlichen Akkumulationsrate von 7 Millionen Tonnen/Jahr entspricht. Diese Menge ist im Vergleich zur Masse des zirkulierenden Kohlenstoffs unbedeutend und fiel sozusagen aus dem Kreislauf heraus und ging in diesem verloren. Daher beträgt der Grad der Offenheit (Unvollkommenheit) des Zyklus 10,4 oder 0,01 %, und dementsprechend beträgt der Grad der Schließung 99,99 %. Dies bedeutet einerseits, dass jedes Kohlenstoffatom am Zyklus teilgenommen hat Zehntausende Male landeten die Stoffe, bevor sie aus dem Kreislauf fielen, in der Tiefe. Andererseits sind die Ströme der Synthese und des Abbaus organischer Stoffe in der Biosphäre sehr präzise aufeinander abgestimmt. 0,2 % des mobilen Kohlenstoffbestands sind ständig im Umlauf. Biomasse-Kohlenstoff erneuert sich in 12 Jahren, in der Atmosphäre in 8 Jahren. Sauerstoffkreislauf. Sauerstoff (O2) spielt eine wichtige Rolle im Leben der meisten Lebewesen auf unserem Planeten. Quantitativ gesehen ist dies der Hauptbestandteil der lebenden Materie. Berücksichtigt man beispielsweise das im Gewebe enthaltene Wasser, besteht der menschliche Körper aus 62,8 % Sauerstoff und 19,4 % Kohlenstoff. Im Allgemeinen ist dieses Element in der Biosphäre im Vergleich zu Kohlenstoff und Wasserstoff das wichtigste unter den einfachen Substanzen. Innerhalb der Biosphäre findet ein schneller Sauerstoffaustausch mit lebenden Organismen oder deren Überresten nach dem Tod statt. Pflanzen produzieren in der Regel freien Sauerstoff, Tiere verbrauchen ihn durch Atmung. Als am weitesten verbreitetes und mobilstes Element auf der Erde schränkt Sauerstoff die Existenz und Funktionen der Ökosphäre nicht ein, obwohl die Verfügbarkeit von Sauerstoff für Wasserorganismen vorübergehend eingeschränkt sein kann. Der Sauerstoffkreislauf in der Biosphäre ist äußerst komplex, da eine Vielzahl organischer und anorganischer Stoffe mit ihm reagieren. Infolgedessen treten viele Epizyklen auf, die zwischen der Lithosphäre und der Atmosphäre oder zwischen der Hydrosphäre und diesen beiden Umgebungen auftreten. Der Sauerstoffkreislauf ähnelt in mancher Hinsicht dem umgekehrten Kohlendioxidkreislauf. Die Bewegung des einen erfolgt in entgegengesetzter Richtung zur Bewegung des anderen. Der Verbrauch von Luftsauerstoff und dessen Ersatz durch Primärproduzenten erfolgt relativ schnell. Somit dauert es 2000 Jahre, um den gesamten Luftsauerstoff vollständig zu erneuern. Heutzutage findet keine Sauerstoffanreicherung in der Atmosphäre statt und sein Gehalt (20,946 %) bleibt konstant. In den oberen Schichten der Atmosphäre entsteht Ozon – O3 – wenn ultraviolette Strahlung auf Sauerstoff einwirkt. Etwa 5 % der die Erde erreichenden Sonnenenergie werden für die Ozonbildung aufgewendet – etwa 8,6 * 1015 W. Die Reaktionen sind leicht reversibel. Beim Zerfall von Ozon wird diese Energie freigesetzt, die in der oberen Atmosphäre hohe Temperaturen aufrechterhält. Die durchschnittliche Ozonkonzentration in der Atmosphäre beträgt etwa 106 Vol.-%. %; die maximale O3-Konzentration – bis zu 4-10 „* Vol.-% – wird in Höhen von 20-25 km erreicht (T.A. Akimova, V.V. Haskin, 1998). Ozon dient als eine Art UV-Filter: Es blockiert einen erheblichen Teil der harten ultravioletten Strahlen. Wahrscheinlich war die Bildung der Ozonschicht eine der Voraussetzungen dafür, dass Leben aus dem Ozean auftauchte und Land besiedelte. Der größte Teil des in geologischen Epochen erzeugten Sauerstoffs verblieb nicht in der Atmosphäre, sondern wurde von der Lithosphäre in Form von Karbonaten, Sulfaten, Eisenoxiden usw. fixiert. Diese Masse beträgt 590 * 1014 Tonnen gegenüber 39 * 1014 Tonnen Sauerstoff, der in der Biosphäre in Form von Gas oder Sulfaten zirkuliert, die in kontinentalen und ozeanischen Gewässern gelöst sind. Stickstoffkreislauf. Stickstoff ist ein essentielles biogenes Element, da er Bestandteil von Proteinen und Nukleinsäuren ist. Der Stickstoffkreislauf ist einer der komplexesten, da er sowohl Gas- als auch Mineralphasen umfasst und gleichzeitig die idealsten Kreisläufe darstellt. Der Stickstoffkreislauf ist eng mit dem Kohlenstoffkreislauf verbunden. Stickstoff folgt in der Regel dem Kohlenstoff, mit dem er an der Bildung aller Eiweißstoffe beteiligt ist. Atmosphärische Luft enthält 78 % Stickstoff und ist ein unerschöpflicher Speicher. Die meisten Lebewesen können diesen Stickstoff jedoch nicht direkt nutzen. Damit Stickstoff von Pflanzen aufgenommen werden kann, muss er Bestandteil von Ammonium- (NH*) oder Nitrat- (NO3) Ionen sein. Durch die Arbeit denitrifizierender Bakterien wird kontinuierlich Stickstoffgas in die Atmosphäre freigesetzt, und fixierende Bakterien nehmen es zusammen mit Blaualgen (Cyanophyten) ständig auf und wandeln es in Nitrate um. Der Stickstoffkreislauf ist auf der Ebene der Destruktoren deutlich sichtbar. Proteine und andere Formen von organischem Stickstoff, die nach ihrem Tod in Pflanzen und Tieren enthalten sind, werden heterotrophen Bakterien, Aktinomyceten und Pilzen (bioreduzierenden Mikroorganismen) ausgesetzt, die die benötigte Energie erzeugen, indem sie diesen organischen Stickstoff reduzieren und in Ammoniak umwandeln. In Böden findet der Prozess der Nitrifikation statt, der aus einer Reaktionskette besteht, bei der unter Beteiligung von Mikroorganismen die Oxidation von Ammoniumionen (NH4+) zu Nitrit (NO~) oder Nitrit zu Nitrat (N0~) erfolgt. Die Reduktion von Nitriten und Nitraten zu den gasförmigen Verbindungen molekularer Stickstoff (N2) oder Lachgas (N20) ist der Kern des Denitrifikationsprozesses. Die anorganische Bildung von Nitraten in geringen Mengen erfolgt ständig in der Atmosphäre, indem bei elektrischen Entladungen bei Gewittern Luftstickstoff mit Sauerstoff gebunden wird und dieser dann mit Regen auf die Bodenoberfläche fällt. Eine weitere Quelle für atmosphärischen Stickstoff sind Vulkane, die den Verlust von aus dem Kreislauf ausgeschlossenem Stickstoff während der Sedimentation oder Ablagerung auf dem Meeresboden ausgleichen. Im Allgemeinen beträgt die durchschnittliche Zufuhr von Nitratstickstoff abiotischen Ursprungs bei der Ablagerung aus der Atmosphäre in den Boden nicht mehr als 10 kg (Jahr/ha), freie Bakterien produzieren 25 kg (Jahr/ha), während die Symbiose von Rhizobien mit Hülsenfrüchten erfolgt produziert durchschnittlich 200 kg (Jahr/ha). Der überwiegende Teil des fixierten Stickstoffs wird durch denitrifizierende Bakterien zu N2 verarbeitet und wieder in die Atmosphäre abgegeben. Nur etwa 10 % des ammonisierten und nitrifizierten Stickstoffs werden von höheren Pflanzen aus dem Boden aufgenommen und stehen vielzelligen Vertretern von Biozönosen zur Verfügung Phosphorkreislauf. Der Phosphorkreislauf in der Biosphäre ist mit Stoffwechselprozessen bei Pflanzen und Tieren verbunden. Dieses wichtige und notwendige Element des Protoplasmas, das in Landpflanzen und Algen zu 0,01–0,1 %, bei Tieren zu 0,1 % bis zu mehreren Prozent enthalten ist, zirkuliert und wandelt sich allmählich von organischen Verbindungen in Phosphate um, die von Pflanzen wieder genutzt werden können. Allerdings bildet Phosphor im Gegensatz zu anderen biophilen Elementen bei der Migration keine Gasform. Das Phosphorreservoir ist nicht wie Stickstoff die Atmosphäre, sondern der mineralische Teil der Lithosphäre. Die Hauptquellen für anorganischen Phosphor sind magmatische Gesteine (Apatite) oder Sedimentgesteine (Phosphorite). Aus Gesteinen zirkuliert anorganischer Phosphor durch Auswaschung und Auflösung in kontinentalen Gewässern. Wenn Phosphor in terrestrische Ökosysteme und Böden gelangt, wird er von Pflanzen aus einer wässrigen Lösung in Form von anorganischen Phosphationen aufgenommen und ist in verschiedenen organischen Verbindungen enthalten, wo er in Form von organischem Phosphat vorkommt. Phosphor gelangt über Nahrungsketten von Pflanzen zu anderen Organismen im Ökosystem. Phosphor wird durch Fließgewässer in aquatische Ökosysteme transportiert. Flüsse reichern die Ozeane kontinuierlich mit Phosphaten an. Wo Phosphor Teil des Phytoplanktons wird. Einige Phosphorverbindungen wandern in geringe Tiefen und werden von Organismen aufgenommen, während der andere Teil in größeren Tiefen verloren geht. Abgestorbene Organismenreste führen zur Anreicherung von Phosphor in unterschiedlichen Tiefen. Schwefelkreislauf. Es gibt zahlreiche gasförmige Schwefelverbindungen, wie zum Beispiel Schwefelwasserstoff H2S und Schwefeldioxid SO2. Der überwiegende Teil des Kreislaufs dieses Elements ist jedoch sedimentärer Natur und findet im Boden und im Wasser statt. Die Verfügbarkeit von anorganischem Schwefel im Ökosystem wird durch die gute Löslichkeit vieler Sulfate im Wasser erleichtert. Pflanzen absorbieren Sulfate, reduzieren sie und produzieren schwefelhaltige Aminosäuren (Methionin, Cystein, Cystin), die eine wichtige Rolle bei der Entwicklung der Tertiärstruktur von Proteinen bei der Bildung von Disulfidbrücken zwischen verschiedenen Zonen der Polypeptidkette spielen. Viele grundlegende Merkmale des biogeochemischen Kreislaufs sind deutlich sichtbar: S -> S -> SO2 - farblose, grüne und violette Schwefelbakterien; - "H2S (anaerobe Reduktion von Sulfat) - Desulfovibno; H2S - "SO2" (aerobe Oxidation von Sulfid) - Thiobacillus; organisches S in SO und H2S. - aerobe bzw. anaerobe heterotrophe Mikroorganismen. Die Primärproduktion sorgt für den Einbau von Sulfat in die organische Substanz und die Ausscheidung durch Tiere dient der Rückführung von Sulfat in den Kreislauf. Generell benötigt das Ökosystem weniger Schwefel als Stickstoff und Phosphor. Daher ist Schwefel seltener ein limitierender Faktor für Pflanzen und Tiere. Gleichzeitig ist der Schwefelkreislauf von zentraler Bedeutung im Gesamtprozess der Biomasseproduktion und -zersetzung. Wenn sich beispielsweise Eisensulfide in Sedimenten bilden, wird Phosphor von einer unlöslichen Form in eine lösliche Form überführt und steht Organismen zur Verfügung. Dies ist eine Bestätigung dafür, wie ein Zyklus durch einen anderen reguliert wird. 2 Energiefluss in Ökosystemen Im Gegensatz zu Stoffen, die kontinuierlich durch verschiedene Blöcke des Ökosystems zirkulieren, jederzeit wiederverwendet werden können und in den Kreislauf gelangen, kann Energie nur einmal verwendet werden, d. h. Es gibt einen linearen Energiefluss durch das Ökosystem. Der einseitige Energiezufluss als universelles Naturphänomen erfolgt aufgrund der Gesetze der Thermodynamik. Das erste Gesetz besagt, dass Energie von einer Form (z. B. Licht) in eine andere (z. B. die potenzielle Energie von Nahrungsmitteln) umgewandelt, aber nicht erzeugt oder zerstört werden kann. Der zweite Hauptsatz besagt, dass es keinen einzigen Prozess geben kann, der mit der Energieumwandlung verbunden ist, ohne dass ein Teil davon verloren geht. Bei solchen Umwandlungen wird ein Teil der Energie in unzugängliche Wärmeenergie umgewandelt und geht somit verloren. Daher kann es beispielsweise nicht zu einer Umwandlung von Nahrungsstoffen in die Substanz, aus der der Körper des Organismus besteht, mit 100-prozentiger Effizienz kommen. Lebewesen sind somit Energiewandler. Und bei jeder Energieumwandlung geht ein Teil davon in Form von Wärme verloren. Letztendlich wird die gesamte Energie, die in den biotischen Kreislauf eines Ökosystems gelangt, als Wärme abgegeben. Lebende Organismen nutzen für ihre Arbeit nicht unbedingt Wärme als Energiequelle – sie nutzen Licht und chemische Energie. Nahrungsketten und -netzwerke, trophische Ebenen. Innerhalb eines Ökosystems werden energiehaltige Substanzen von autotrophen Organismen erzeugt und dienen Heterotrophen als Nahrung. Nahrungsverbindungen sind Mechanismen zur Energieübertragung von einem Organismus auf einen anderen. Ein typisches Beispiel: Ein Tier frisst Pflanzen. Dieses Tier wiederum kann von einem anderen Tier gefressen werden. Auf diese Weise kann Energie durch mehrere Organismen übertragen werden – jeder nachfolgende ernährt sich vom vorherigen, der ihn mit Rohstoffen und Energie versorgt. Diese Abfolge der Energieübertragung wird als Nahrungskette (trophische Kette) oder Nahrungskette bezeichnet. Die Position jedes Glieds in der Nahrungskette ist eine trophische Ebene. Die erste trophische Ebene wird, wie bereits erwähnt, von Autotrophen oder sogenannten Primärproduzenten besetzt. Organismen der zweiten trophischen Ebene werden als Primärkonsumenten bezeichnet, die dritte als Sekundärkonsumenten usw. Im Allgemeinen gibt es drei Arten von Nahrungsketten: Die Nahrungskette der Fleischfresser beginnt bei Pflanzen und verläuft von kleinen Organismen zu immer größeren Organismen. An Land bestehen Nahrungsketten aus drei bis vier Gliedern. Eine der einfachsten Nahrungsketten sieht so aus: Pflanze -> Hase -> Wolf Produzent -" Pflanzenfresser -> -> Fleischfresser Weiter verbreitet sind folgende Nahrungsketten: Pflanzenmaterial (z. B. Nektar) – „Fliege“ – „Spinne -> Spitzmaus -> Eule“. Rosenstrauchsaft -> Blattlaus -> -> Marienkäfer -> -> Spinne - „insektenfressender Vogel -> Raubvogel“. In aquatischen und insbesondere marinen Ökosystemen sind die Nahrungsketten von Raubtieren tendenziell länger als in terrestrischen. Die dritte Art von Nahrungsketten, beginnend mit abgestorbenen Pflanzenresten, Kadavern und tierischen Exkrementen, werden als detritale (saprophytische) Nahrungsketten oder detritale Zersetzungsketten bezeichnet. Laubwälder spielen eine wichtige Rolle in den schädlichen Nahrungsketten terrestrischer Ökosysteme, deren Laub größtenteils nicht von Pflanzenfressern gefressen wird und Teil der Abfälle aus abgefallenem Laub ist. Die Blätter werden von zahlreichen Detritivoren – Pilzen, Bakterien, Insekten (z. B. Collembola) usw. – zerkleinert und dann von Regenwürmern aufgenommen, die den Humus gleichmäßig in der Oberflächenschicht der Erde verteilen und den sogenannten Mull bilden. Auf dieser Ebene entwickeln Pilze Myzel. Die zersetzenden Mikroorganismen, die die Kette vervollständigen, bewirken die endgültige Mineralisierung abgestorbener organischer Substanz. Im Allgemeinen lassen sich die typischen schädlichen Nahrungsketten unserer Wälder wie folgt darstellen: Laubstreu -> Regenwurm -> Amsel - Sperber; Totes Tier - „Larven der Aasfliege“ – Grasfrosch -> Grasnatter. In den besprochenen Nahrungskettendiagrammen wird jeder Organismus so dargestellt, dass er sich von anderen Organismen einer Art ernährt. Die tatsächlichen Nahrungszusammenhänge in einem Ökosystem sind viel komplexer, da sich ein Tier von Organismen unterschiedlicher Art aus derselben Nahrungskette oder aus unterschiedlichen Nahrungsketten ernähren kann, beispielsweise von Raubtieren der oberen trophischen Ebenen. Tiere ernähren sich oft sowohl von Pflanzen als auch von anderen Tieren. Sie werden Allesfresser genannt. Nahrungsnetze in Ökosystemen sind sehr komplex, und wir können daraus schließen, dass die in sie eintretende Energie lange braucht, um von einem Organismus zum anderen zu gelangen. Ökologische Pyramiden. Innerhalb jedes Ökosystems haben Nahrungsnetze eine genau definierte Struktur, die durch die Art und Anzahl der Organismen gekennzeichnet ist, die auf jeder Ebene der verschiedenen Nahrungsketten vertreten sind. Um die Beziehungen zwischen Organismen in einem Ökosystem zu untersuchen und grafisch darzustellen, verwenden sie in der Regel ökologische Pyramiden anstelle von Nahrungsnetzdiagrammen. Ökologische Pyramiden drücken die trophische Struktur eines Ökosystems in geometrischer Form aus. Sie sind in Form von Rechtecken gleicher Breite aufgebaut, die Länge der Rechtecke muss jedoch proportional zum Wert des Messobjekts sein. Von hier aus können Sie Pyramiden aus Zahlen, Biomasse und Energie erhalten. Ökologische Pyramiden spiegeln die grundlegenden Merkmale jeder Biozönose wider, indem sie ihre trophische Struktur zeigen. Zahlenpyramiden. Sie stellen die einfachste Annäherung an die Untersuchung der trophischen Struktur eines Ökosystems dar. Biomassepyramide. Spiegelt die Ernährungsverhältnisse im Ökosystem besser wider, da die Gesamtmasse der Organismen (Biomasse) jeder trophischen Ebene berücksichtigt wird. Energiepyramide. Die grundlegendste Möglichkeit, Verbindungen zwischen Organismen auf verschiedenen trophischen Ebenen darzustellen, sind Energiepyramiden. Sie stellen die Energieumwandlungseffizienz und Produktivität von Nahrungsketten dar und werden durch Zählen der Energiemenge (kcal) ermittelt, die pro Flächeneinheit und Zeiteinheit angesammelt und von Organismen auf jeder trophischen Ebene verbraucht wird. Die von der Pflanze aufgenommene Sonnenenergie wird bei der Photosynthese nur teilweise genutzt. Die in Kohlenhydraten fixierte Energie stellt die Bruttoproduktion des Ökosystems dar. Kohlenhydrate werden zum Aufbau von Protoplasma und zum Pflanzenwachstum verwendet. Ein Teil ihrer Energie wird für die Atmung aufgewendet. Konsumenten zweiter Ordnung (Raubtiere) zerstören nicht die gesamte Biomasse ihrer Opfer. Darüber hinaus wird von der Menge, die sie zerstören, nur ein Teil zur Bildung von Biomasse ihres eigenen trophischen Niveaus verwendet. Der Rest wird hauptsächlich für die Atemenergie aufgewendet und mit Kot und Exkrementen ausgeschieden. Im Jahr 1942 formulierte R. Lindeman erstmals das Gesetz der Energiepyramide, das in Lehrbüchern oft als „Gesetz von 10 %“ bezeichnet wird. Nach diesem Gesetz bewegen sich im Durchschnitt nicht mehr als 10 % der Energie von einer trophischen Ebene der ökologischen Pyramide auf eine andere. Nur 10–20 % der ursprünglichen Energie werden an nachfolgende Heterotrophe übertragen. Mit dem Gesetz der Energiepyramide lässt sich leicht berechnen, dass die Energiemenge, die tertiäre Fleischfresser (trophische Ebene V) erreicht, etwa 0,0001 der von den Produzenten absorbierten Energie beträgt. Daraus folgt, dass die Energieübertragung von einer Ebene auf eine andere mit sehr geringem Wirkungsgrad erfolgt. Dies erklärt die begrenzte Anzahl von Gliedern in der Nahrungskette, unabhängig von einer bestimmten Biozönose. E. Odum (1959) in einer extrem vereinfachten Nahrungskette - Luzerne -> Kalb - „Das Kind beurteilte die Energieumwandlung und veranschaulichte das Ausmaß seiner Verluste. Nehmen wir an, überlegte er, auf einer Fläche von 4 Hektar. Auf diesem Feld ernähren sich Kälber (man geht davon aus, dass sie nur Luzerne fressen), und ein 12-jähriger Junge isst ausschließlich Kalbfleisch. Die Ergebnisse der Berechnungen, dargestellt in Form von drei Pyramiden: Zahlen, Biomasse und Energie, weisen darauf hin, dass Luzerne nur 0,24 % der gesamten auf das Feld fallenden Sonnenenergie verbraucht; das Kalb absorbiert 8 % dieses Produkts und nur 0,7 % der Biomasse des Kalbes gewährleisten die Entwicklung des Kindes im Laufe des Jahres. E. Odum zeigte somit, dass nur ein Millionstel der einfallenden Sonnenenergie in Fleischfresser-Biomasse umgewandelt wird, was in diesem Fall zu einer Gewichtszunahme des Kindes beiträgt, und der Rest geht verloren und wird in abgebauter Form in der Umwelt abgegeben. Das obige Beispiel verdeutlicht die sehr geringe ökologische Effizienz von Ökosystemen und die geringe Effizienz der Transformation in Nahrungsketten. Wir können Folgendes feststellen: Wenn 1000 kcal (Tag m 2) von den Herstellern aufgezeichnet, dann 10 kcal (Tag m 2) geht in die Biomasse von Pflanzenfressern ein und nur 1 kcal (Tag m 2) - in die Biomasse von Fleischfressern. Da eine bestimmte Menge eines Stoffes von jeder Biozönose wiederholt und ein Teil der Energie einmalig genutzt werden kann, ist es sinnvoller zu sagen, dass es im Ökosystem zu einer Kaskadenübertragung von Energie kommt. Verbraucher fungieren als steuerndes und stabilisierendes Glied im Ökosystem. Verbraucher erzeugen in der Volkszählung ein Spektrum an Vielfalt und verhindern so das Monopol der Dominanten. Die Regel der Wertkontrolle der Verbraucher kann zu Recht als ganz grundlegend angesehen werden. Nach kybernetischer Auffassung sollte das Kontrollsystem komplexer aufgebaut sein als das gesteuerte, dann wird der Grund für die Vielfalt der Instrumententypen klar. Die steuernde Bedeutung der Verbraucher hat auch eine energetische Grundlage. Der Energiefluss durch die eine oder andere trophische Ebene kann nicht absolut durch die Verfügbarkeit von Nahrungsmitteln in der zugrunde liegenden trophischen Ebene bestimmt werden. Es bleibt bekanntlich immer eine ausreichende „Reserve“ übrig, da die völlige Vernichtung von Lebensmitteln zum Tod der Verbraucher führen würde. Diese allgemeinen Muster werden im Rahmen von Populationsprozessen, Gemeinschaften, Ebenen der ökologischen Pyramide und Biozönosen insgesamt beobachtet. 3 Dynamische Prozesse, die Integrität und Nachhaltigkeit in Ökosystemen gewährleisten Zyklische Veränderungen in Gemeinschaften spiegeln die tägliche, saisonale und langfristige Periodizität äußerer Bedingungen und die Manifestation endogener Rhythmen von Organismen wider. Die tägliche Dynamik von Ökosystemen hängt hauptsächlich mit dem Rhythmus natürlicher Phänomene zusammen und ist streng periodischer Natur. Wir haben bereits berücksichtigt, dass es in jeder Biozönose Gruppen von Organismen gibt, deren Lebensaktivität zu unterschiedlichen Tageszeiten stattfindet. Einige sind tagsüber aktiv, andere nachts. Daher kommt es zu periodischen Veränderungen in der Zusammensetzung und dem Verhältnis einzelner Arten der Biozönose eines bestimmten Ökosystems, da einzelne Organismen für eine bestimmte Zeit von diesem abgeschaltet werden. Die tägliche Dynamik der Biozönose wird sowohl von Tieren als auch von Pflanzen bereitgestellt. Wie Sie wissen, ändern sich Intensität und Art physiologischer Prozesse in Pflanzen im Laufe des Tages – die Photosynthese findet nachts nicht statt, oft öffnen sich Blüten in Pflanzen nur nachts und werden von nachtaktiven Tieren bestäubt, andere sind tagsüber an die Bestäubung angepasst. Die Tagesdynamik in Biozönosen ist in der Regel umso ausgeprägter, je größer der Unterschied in Temperatur, Luftfeuchtigkeit und anderen Umweltfaktoren zwischen Tag und Nacht ist. Bei der saisonalen Dynamik werden größere Abweichungen in den Biozönosen beobachtet. Dies liegt an den biologischen Kreisläufen von Organismen, die von der saisonalen Zyklizität natürlicher Phänomene abhängen. Somit hat der Wechsel der Jahreszeiten einen erheblichen Einfluss auf die Lebensaktivität von Tieren und Pflanzen (Winterschlaf, Winterschlaf, Diapause und Migration bei Tieren; Perioden der Blüte, Fruchtbildung, aktiven Wachstums, Laubfall und Winterruhe bei Pflanzen). Die abgestufte Struktur der Biozönose unterliegt häufig saisonalen Schwankungen. Einzelne Pflanzenschichten können in den entsprechenden Jahreszeiten vollständig verschwinden, beispielsweise eine krautige Schicht aus einjährigen Pflanzen. Die Dauer der biologischen Jahreszeiten variiert je nach Breitengrad. In dieser Hinsicht ist die saisonale Dynamik von Biozönosen in der Arktis, den gemäßigten und tropischen Zonen unterschiedlich. Es kommt am deutlichsten in Ökosystemen gemäßigten Klimas und nördlichen Breiten zum Ausdruck. Langfristige Variabilität ist im Leben jeder Biozönose normal. So schwankt die Niederschlagsmenge in der Waldsteppe von Barabinsk von Jahr zu Jahr stark; mehrere trockene Jahre wechseln sich mit längerfristigen Perioden reichlicher Niederschläge ab. Dies hat erhebliche Auswirkungen auf Pflanzen und Tiere. In diesem Fall kommt es zur Entwicklung ökologischer Nischen – funktionale Abgrenzung in der entstehenden Menge oder deren Ergänzung mit geringer Diversität. Langfristige Veränderungen in der Zusammensetzung von Biozönosen wiederholen sich auch im Zusammenhang mit periodischen Veränderungen der allgemeinen Zirkulation der Atmosphäre, die wiederum durch eine Zunahme oder Abnahme der Sonnenaktivität verursacht werden. Im Zuge der Tages- und Saisondynamik wird die Integrität von Biozönosen in der Regel nicht verletzt. Die Biozönose unterliegt nur periodischen Schwankungen qualitativer und quantitativer Merkmale. Fortschreitende Veränderungen im Ökosystem führen letztendlich dazu, dass eine Biozönose durch eine andere mit einer anderen Gruppe dominanter Arten ersetzt wird. Ursachen für solche Veränderungen können biozönoseexterne Faktoren sein, die über einen längeren Zeitraum in eine Richtung wirken, beispielsweise eine zunehmende Verschmutzung von Gewässern, eine zunehmende Austrocknung von Sumpfböden durch Rekultivierung, eine verstärkte Beweidung usw. Diese Veränderungen von einer Biozönose zur anderen werden als exogenetisch bezeichnet. Wenn der zunehmende Einfluss eines Faktors zu einer allmählichen Vereinfachung der Struktur der Biozönose, einer Erschöpfung ihrer Zusammensetzung und einem Rückgang der Produktivität führt, werden solche Verschiebungen als degressiv oder degressiv bezeichnet. Endogene Veränderungen entstehen durch Prozesse, die innerhalb der Biozönose selbst ablaufen. Die sukzessive Ersetzung einer Biozönose durch eine andere nennt man ökologische Sukzession (von lat. – Sukzession – Abfolge, Veränderung). Nachfolge ist ein Prozess der Selbstentwicklung von Ökosystemen. Die Nachfolge basiert auf der Unvollständigkeit des biologischen Zyklus in einer bestimmten Biozönose. Es ist bekannt, dass lebende Organismen aufgrund ihrer lebenswichtigen Aktivität die Umgebung um sie herum verändern, indem sie ihr einige Substanzen entziehen und sie mit Stoffwechselprodukten sättigen. Wenn Populationen über einen längeren Zeitraum bestehen, verändern sie ihre Umwelt in eine ungünstige Richtung und werden dadurch durch Populationen anderer Arten verdrängt, für die sich die daraus resultierenden Umweltveränderungen als ökologisch vorteilhaft erweisen. Bei einer Biozönose kommt es also zu einem Wechsel dominanter Arten. Die Regel (das Prinzip) der ökologischen Vervielfältigung ist hier deutlich sichtbar. Das langfristige Bestehen einer Biozönose ist nur möglich, wenn Veränderungen in der Umwelt, die durch die Aktivität einiger lebender Organismen verursacht werden, für andere mit entgegengesetzten Anforderungen günstig sind. Basierend auf konkurrierenden Interaktionen der Arten während der Sukzession werden nach und nach stabilere Kombinationen gebildet, die spezifischen abiotischen Umweltbedingungen entsprechen. Ein Beispiel für Sukzession, die zur Ersetzung einer Gemeinde durch eine andere führt, ist die Überwucherung eines kleinen Sees, gefolgt von der Entstehung eines Sumpfes und dann eines Waldes an seiner Stelle. Zunächst bildet sich entlang der Seeränder ein schwimmender Teppich – ein schwimmender Teppich aus Seggen, Moosen und anderen Pflanzen. Der See ist ständig mit toten Pflanzenresten gefüllt – Torf. Es entsteht ein Sumpf, der nach und nach mit Wald bewachsen wird. Eine aufeinanderfolgende Reihe von Gemeinschaften, die sich nach und nach auf natürliche Weise gegenseitig ersetzen, wird als Nachfolgereihe bezeichnet. Sukzessionen in der Natur sind in ihrem Ausmaß äußerst unterschiedlich. Sie können in Ufern mit Kulturen beobachtet werden, bei denen es sich um planktonische Gemeinschaften handelt – verschiedene Arten von Schwebealgen und deren Konsumenten – Rädertierchen, Flagellaten in Pfützen und Teichen, Sümpfen, Wiesen, Wäldern, verlassenen Ackerflächen, verwitterten Felsen usw. Es gibt eine Hierarchie in der Organisation von Ökosystemen äußert es sich auch in Sukzessionsprozessen – größere Transformationen von Biozönosen setzen sich aus kleineren zusammen. In stabilen Ökosystemen mit einem regulierten Stoffkreislauf finden zudem ständig lokale Sukzessionsveränderungen statt, die die komplexe innere Struktur von Lebensgemeinschaften unterstützen. Arten sukzessiver Veränderungen. Es gibt zwei Haupttypen sukzessiver Veränderungen: 1 – unter Beteiligung autotropher und heterotropher Populationen; 2 - nur unter Beteiligung von Heterotrophen. Die Sukzession der zweiten Art findet nur unter Bedingungen statt, unter denen eine Vorversorgung oder ständige Versorgung mit organischen Verbindungen entsteht, aufgrund derer die Gemeinschaft existiert: in Misthaufen oder Misthaufen, in zersetzendem Pflanzenmaterial, in mit organischen Substanzen verunreinigten Reservoirs usw. Nachfolgeprozess. Nach F. Clements (1916) besteht der Nachfolgeprozess aus folgenden Phasen: Die Sukzession mit Vegetationswechsel kann primär und sekundär sein. Primäre Sukzession ist der Prozess der Entwicklung und Veränderung von Ökosystemen in zuvor unbewohnten Gebieten, beginnend mit ihrer Besiedlung. Ein klassisches Beispiel ist die ständige Verschmutzung kahler Felsen, auf denen sich schließlich Wälder entwickeln. So werden bei den Primärsukzessionen auf den Felsen des Uralgebirges folgende Stadien unterschieden: Sekundäre Sukzession ist die Wiederherstellung eines Ökosystems, das einst in einem bestimmten Gebiet existierte. Es beginnt, wenn die etablierten Beziehungen der Organismen in einer etablierten Biozönose durch einen Vulkanausbruch, Feuer, Holzeinschlag, Pflügen usw. gestört werden. Verschiebungen, die zur Wiederherstellung der vorherigen Biozönose führen, werden in der Geobotanik als Demutation bezeichnet. Ein Beispiel ist die Dynamik der Artenvielfalt auf der Insel Krakatau nach der vollständigen Zerstörung der einheimischen Flora und Fauna durch einen Vulkan. Ein weiteres Beispiel ist die sekundäre Sukzession des sibirischen dunklen Nadelwaldes (Tannen-Zedern-Taiga) nach einem verheerenden Waldbrand. In stärker verbrannten Gebieten entstehen Pioniermoose aus vom Wind verwehten Sporen: 3-5 Jahre nach einem Brand ist Funaria hygrometrica, Geratodon purpureus usw. das am häufigsten vorkommende „Feuermoos“. Unter höheren Pflanzen wächst sehr schnell Weidenröschen (Chamaenerion angustifolium). besiedelt verbrannte Flächen ), die bereits nach 2-3 Monaten im Feuer üppig blüht, sowie gemahlenes Schilfgras (Calamagrostis epigeios) und andere Arten. Weitere Sukzessionsphasen sind zu beobachten: Die Schilfwiese weicht Sträuchern, dann Birken- oder Espenwald, Kiefernmischwald, Kiefernwald, Kiefern-Zedern-Wald und schließlich, nach 250 Jahren, erfolgt die Wiederherstellung des Zedern-Tannen-Waldes . Sekundäre Sukzessionen erfolgen in der Regel schneller und einfacher als primäre, da im gestörten Lebensraum das Bodenprofil, Samen, Primordien und ein Teil der vorherigen Population und früheren Verbindungen erhalten bleiben. Bei der Demutation handelt es sich nicht um eine Wiederholung irgendeines Stadiums der primären Sukzession. Höhepunkt-Ökosystem. Die Sukzession endet mit einem Stadium, in dem alle Arten des Ökosystems bei der Fortpflanzung eine relativ konstante Anzahl beibehalten und keine weiteren Änderungen in ihrer Zusammensetzung auftreten. Dieser Gleichgewichtszustand wird als Höhepunkt bezeichnet, und das Ökosystem wird als Höhepunkt bezeichnet. Unter unterschiedlichen abiotischen Bedingungen bilden sich unterschiedliche Klimax-Ökosysteme aus. In einem heißen und feuchten Klima wird es ein tropischer Regenwald sein, in einem trockenen und heißen Klima wird es eine Wüste sein. Die Hauptbiome der Erde sind die Höhepunktökosysteme ihrer jeweiligen geografischen Gebiete. Veränderungen im Ökosystem während der Sukzession. Produktivität und Biomasse. Wie bereits erwähnt, ist die Sukzession ein natürlicher, gerichteter Prozess und die Veränderungen, die in der einen oder anderen Phase auftreten, sind charakteristisch für jede Gemeinschaft und hängen nicht von ihrer Artenzusammensetzung oder ihrem geografischen Standort ab. Es gibt vier Haupttypen von Sukzessionswechseln: Zu beachten ist auch, dass der Wechsel der Nachfolgephasen nach bestimmten Regeln erfolgt. Jede Phase bereitet die Umgebung auf die Entstehung der nächsten vor. Hier gilt das Gesetz der Abfolge der Entwicklungsphasen; die Entwicklungsphasen eines natürlichen Systems können nur in einer evolutionär festgelegten (historisch, umweltbedingten) Reihenfolge folgen, normalerweise von relativ einfach zu komplex, in der Regel ohne Verlust von Zwischenstadien, aber möglicherweise mit ihrem sehr schnellen Ablauf oder ihrer evolutionär festgelegten Abwesenheit. Wenn sich ein Ökosystem dem Zustand der Menopause nähert, verlangsamen sich darin, wie in allen Gleichgewichtssystemen, alle Entwicklungsprozesse. Diese Situation spiegelt sich im Gesetz der sukzessiven Verlangsamung wider: Prozesse in ausgereiften Gleichgewichtsökosystemen, die sich in einem stabilen Zustand befinden, neigen in der Regel dazu, sich zu verlangsamen. In diesem Fall ändert sich der restaurative Typ der Nachfolge in ihren säkularen Verlauf, d. h. Die Selbstentwicklung erfolgt innerhalb der Grenzen der Menopause oder der Knotenentwicklung. Das empirische Gesetz der sukzessiven Verlangsamung ist eine Folge der Regel von G. Odum und R. Pinkerton bzw. der Regel der maximalen Energie zur Aufrechterhaltung eines ausgereiften Systems: Die Sukzession schreitet in Richtung einer grundlegenden Verschiebung des Energieflusses hin zu seiner Steigerung voran Menge, die auf die Aufrechterhaltung des Systems abzielt. Die Regel von G. Odum und R. Pinkerton wiederum basiert auf der von A. Lotka formulierten Regel der maximalen Energie in biologischen Systemen. Diese Frage wurde später von R. Margalef und Y. Odum gut entwickelt und gilt als Beweis für das Prinzip des „Nullmaximums“ oder der Minimierung des Wachstums in einem ausgereiften Ökosystem: Ein Ökosystem in sukzessiver Entwicklung neigt dazu, die größte Biomasse zu bilden geringste biologische Produktivität. Lindeman (1942) hat experimentell nachgewiesen, dass die Sukzession mit einer Steigerung der Produktivität bis zur Höhepunktgemeinschaft einhergeht, in der die Energieumwandlung am effizientesten erfolgt. Daten aus Studien zur Sukzession von Eichen- und Eichen-Eschenwäldern zeigen, dass ihre Produktivität in späteren Stadien tatsächlich zunimmt. Während des Übergangs zu einer Höhepunktgemeinschaft kommt es jedoch normalerweise zu einem Rückgang der Gesamtproduktivität. Daher ist die Produktivität in alten Wäldern geringer als in jungen Wäldern, die wiederum möglicherweise weniger produktiv sind als die ihnen vorausgehenden artenreicheren Krautschichten. Ähnliche Produktivitätsrückgänge wurden in einigen aquatischen Systemen beobachtet. Dafür gibt es mehrere Gründe. Einer davon ist, dass die Anreicherung von Nährstoffen in der wachsenden Waldbiomasse zu einer Verringerung ihres Kreislaufs führen kann. Der Rückgang der Gesamtproduktivität könnte einfach das Ergebnis einer verminderten Vitalität einzelner Personen sein, wenn ihr Durchschnittsalter in der Gemeinschaft steigt. Mit fortschreitender Sukzession reichert sich ein zunehmender Anteil der verfügbaren Nährstoffe in der Biomasse der Lebensgemeinschaft an und dementsprechend nimmt ihr Gehalt in der abiotischen Komponente des Ökosystems (im Boden oder im Wasser) ab. Auch die Menge an produziertem Detritus nimmt zu. Die Hauptkonsumenten sind keine Pflanzenfresser, sondern schädliche Organismen. Entsprechende Veränderungen treten auch in trophischen Netzwerken auf. Detritus wird zur Hauptnährstoffquelle. Mit der Sukzession nimmt die Geschlossenheit biogeochemischer Stoffkreisläufe zu. Ungefähr 10 Jahre nach Beginn der Wiederherstellung der Vegetationsdecke nimmt die Offenheit der Zyklen von 100 auf 10 % ab und nimmt dann noch mehr ab, bis sie in der Höhepunktphase ein Minimum erreicht. Die Regel der zunehmenden Geschlossenheit des biogeochemischen Stoffkreislaufs im Laufe der Sukzession wird mit Sicherheit durch die anthropogene Transformation der Vegetation und natürlicher Ökosysteme im Allgemeinen verletzt. Zweifellos führt dies zu einer langen Reihe von Anomalien in der Biosphäre und ihren Teilungen. Ein Rückgang der Artenvielfalt in den Wechseljahren bedeutet nicht, dass sie von geringerer ökologischer Bedeutung ist. Die Artenvielfalt prägt die Sukzession, ihre Richtung und sorgt dafür, dass der reale Raum mit Leben gefüllt ist. Eine unzureichende Anzahl von Arten, aus denen der Komplex besteht, konnte keine Sukzessionsreihe bilden, und mit der Zerstörung der Höhepunktökosysteme würde es allmählich zu einer vollständigen Wüstenbildung des Planeten kommen. Der Wert der Vielfalt ist sowohl statisch als auch dynamisch funktional. Es ist zu beachten, dass dort, wo die Artenvielfalt nicht ausreicht, um die Biosphäre zu bilden, die als Grundlage für den normalen natürlichen Ablauf des Sukzessionsprozesses dient, und die Umwelt selbst stark gestört ist, die Sukzession nicht die Höhepunktphase erreicht, sondern endet mit einer Knotengemeinschaft – einem Paraklimax, einer langfristigen oder kurzfristigen abgeleiteten Gemeinschaft. Je tiefer die Störung der Umgebung eines bestimmten Raums ist, desto früher endet die Phasenfolge. Wenn eine Art oder eine Gruppe von Arten infolge ihrer Zerstörung verloren geht (anthropogenes Verschwinden von Lebensräumen, seltener Aussterben), ist das Erreichen der Wechseljahre keine vollständige Wiederherstellung der natürlichen Umwelt. Tatsächlich handelt es sich um ein neues Ökosystem, da in ihm neue Verbindungen entstanden sind, viele alte verloren gegangen sind und sich eine andere „Ausfransung“ der Arten entwickelt hat. Das Ökosystem kann nicht in seinen alten Zustand zurückkehren, da eine verlorene Art nicht wiederhergestellt werden kann. Wenn sich ein abiotischer oder biotischer Faktor ändert, zum Beispiel durch anhaltende Abkühlung oder die Einführung einer neuen Art, wird eine Art, die schlecht an neue Bedingungen angepasst ist, einen von drei Wegen einschlagen. Das Gesetz der evolutionär-ökologischen Irreversibilität besagt, dass ein Ökosystem, das aufgrund eines Ungleichgewichts ökologischer Komponenten einen Teil seiner Elemente verloren hat oder durch ein anderes ersetzt wurde, im Laufe der Sukzession nicht in seinen ursprünglichen Zustand zurückkehren kann, wenn während der Veränderungen evolutionäre (mikroevolutionäre) Veränderungen auftreten ) Es sind Veränderungen in ökologischen Elementen aufgetreten (konserviert oder vorübergehend verloren). Für den Fall, dass einige Arten in Zwischenphasen der Sukzession verloren gehen, kann dieser Verlust zwar funktionell kompensiert werden, jedoch nicht vollständig. Wenn die Diversität über ein kritisches Maß hinaus abnimmt, wird der Ablauf der Nachfolge verzerrt und es kann tatsächlich kein identischer Höhepunkt wie in der Vergangenheit erreicht werden. Um die Natur wiederhergestellter Ökosysteme beurteilen zu können, ist das Gesetz der evolutionär-ökologischen Irreversibilität wichtig. Mit dem Verlust von Elementen handelt es sich tatsächlich um völlig neue ökologische Naturformationen mit neu gebildeten Mustern und Verbindungen. Somit bedeutet der Transfer einer Art, die während ihrer Reakklimatisierung aus dem Ökosystem ausgeschieden ist, in die Vergangenheit nicht ihre mechanische Rückkehr. Dabei handelt es sich eigentlich um die Einführung einer neuen Art in ein erneuertes Ökosystem. Das Gesetz der evolutionär-ökologischen Irreversibilität betont die Richtung der Evolution nicht nur auf der Ebene von Biosystemen, sondern auch auf allen anderen hierarchischen Ebenen der Biota. 4 Die Biosphäre als globales Ökosystem, das die Integrität und Nachhaltigkeit von Ökosystemen gewährleistet Die Biosphäre ist ein globales Ökosystem. Wie bereits erwähnt, wird die Biosphäre in Geobiosphäre, Hydrobiosphäre und Aerobiosphäre unterteilt (Abb. 2.4). Die Geobiosphäre ist entsprechend den wichtigsten umweltbildenden Faktoren unterteilt: Terrabiosphäre und Lithobiosphäre – innerhalb der Geobiosphäre, Marinobiosphäre (Ozeanobiosphäre) und Aquabiosphäre – innerhalb der Hydrobiosphäre. Diese Formationen werden Subsphären genannt. Der führende umweltbildende Faktor bei ihrer Entstehung ist die physikalische Phase der Lebensumgebung: Luft-Wasser in der Aerobiosphäre, Wasser – Süßwasser und Salzwasser in der Hydrobiosphäre, feste Luft in der Terrabiosphäre und festes Wasser in der Lithobiosphäre. Sie zerfallen wiederum alle in Schichten: die Aerobiosphäre – in die Tropobiosphäre und Altobiosphäre; Hydrobiosphäre - in Photosphäre, Disphotosphäre und Aphotosphäre. Die strukturbildenden Faktoren sind hier neben der physikalischen Umgebung Energie (Licht und Wärme), besondere Bedingungen für die Entstehung und Entwicklung des Lebens – die evolutionären Richtungen des Eindringens von Biota auf das Land, in seine Tiefen, in die darüber liegenden Räume Erde, der Abgrund des Ozeans, sind zweifellos anders. Zusammen mit der Apobiosphäre, Parabiosphäre und anderen Sub- und Suprabiosphärenschichten bilden sie den sogenannten „Schichtkuchen des Lebens“ und die Geosphäre (Ökosphäre) seiner Existenz innerhalb der Grenzen der Megabiosphäre. Im systemischen Sinne handelt es sich bei den aufgeführten Formationen um große Funktionsteile von praktisch universellen oder subplanetaren Dimensionen. Wissenschaftler gehen davon aus, dass es in der Biosphäre mindestens 8–9 Ebenen relativ unabhängiger Stoffkreisläufe innerhalb der Verbindung von 7 materiellen und energieökologischen Hauptkomponenten und der 8. Ebene – der Information – gibt. Globale, regionale und lokale Stoffkreisläufe sind nicht geschlossen und „überschneiden“ sich teilweise innerhalb der Ökosystemhierarchie. Diese stofflich-energetische und teilweise auch informationelle „Kopplung“ sichert die Integrität ökologischer Supersysteme bis hin zur Biosphäre als Ganzes. Integrität und Nachhaltigkeit von Ökosystemen. Die Biosphäre wird in größerem Maße nicht durch äußere Faktoren, sondern durch innere Muster geformt. Die wichtigste Eigenschaft der Biosphäre ist die Wechselwirkung von lebenden und nicht lebenden Dingen, die sich im Gesetz der biogenen Migration von Atomen von V. I. Wernadskij widerspiegelt. Das Gesetz der biogenen Migration von Atomen ermöglicht es der Menschheit, biogeochemische Prozesse sowohl auf der Erde als Ganzes als auch in ihren Regionen bewusst zu steuern. Die Menge der lebenden Materie in der Biosphäre unterliegt bekanntlich keinen nennenswerten Veränderungen. Dieses Muster wurde von V.I. in Form des Gesetzes der Konstanz der Menge lebender Materie formuliert. Wernadskij: Die Menge an lebender Materie in der Biosphäre für einen bestimmten geologischen Zeitraum ist eine Konstante. In der Praxis ist dieses Gesetz eine quantitative Konsequenz des Gesetzes des inneren dynamischen Gleichgewichts für das globale Ökosystem – die Biosphäre. Da lebende Materie gemäß dem Gesetz der biogenen Migration von Atomen ein Energievermittler zwischen Sonne und Erde ist, muss ihre Menge entweder konstant sein oder ihre Energieeigenschaften müssen sich ändern. Das Gesetz der physikalischen und chemischen Einheit der lebenden Materie (alle lebende Materie der Erde ist physikalisch und chemisch vereint und schließt wesentliche Änderungen der letztgenannten Eigenschaft aus. Daher ist quantitative Stabilität für die lebende Materie des Planeten unvermeidlich. Sie ist völlig charakteristisch für die Anzahl der Arten. Als Speicher für Sonnenenergie muss lebende Materie gleichzeitig auf äußere (kosmische) Einflüsse und innere Veränderungen reagieren. Eine Abnahme oder Zunahme der Menge lebender Materie an einem Ort der Biosphäre sollte an einem anderen Ort zu einem genau umgekehrten Prozess führen, da die freigesetzten Nährstoffe vom Rest der lebenden Materie aufgenommen werden können oder ihr Mangel beobachtet wird. Dabei ist die Geschwindigkeit des Prozesses zu berücksichtigen, die bei anthropogenen Veränderungen viel geringer ist als bei direkten Eingriffen in die Natur durch den Menschen. Neben der Konstanz und Konstanz der Menge der lebenden Materie, die sich im Gesetz der physikalischen und chemischen Einheit der lebenden Materie widerspiegelt, kommt es in der belebten Natur zu einer ständigen Erhaltung der Informations- und Körperstruktur, trotz der Tatsache, dass sie sich verändert etwas mit dem Lauf der Evolution. Diese Eigenschaft wurde von Yu. Goldsmith (1981) festgestellt und als Gesetz zur Erhaltung der Struktur der Biosphäre – informativ und somatisch – oder als erstes Gesetz der Ökodynamik bezeichnet. Um die Struktur der Biosphäre zu erhalten, streben Lebewesen danach, einen Reifezustand oder ein ökologisches Gleichgewicht zu erreichen. Das Gesetz des Wunsches nach Wechseljahren – das zweite Gesetz der Ökodynamik von Yu. Goldsmith – gilt für die Biosphäre und andere Ebenen ökologischer Systeme, obwohl es Besonderheiten gibt – die Biosphäre ist ein geschlosseneres System als ihre Unterteilungen. Die Einheit der lebenden Materie der Biosphäre und die Homologie der Struktur ihrer Subsysteme führen dazu, dass die darauf entstandenen lebenden Elemente unterschiedlichen geologischen Zeitalters und ursprünglichen geografischen Ursprungs evolutionär eng miteinander verflochten sind. Die Verflechtung von Elementen unterschiedlicher räumlich-zeitlicher Genese auf allen ökologischen Ebenen der Biosphäre spiegelt die Regel oder das Prinzip der Heterogenese lebender Materie wider. Diese Addition erfolgt nicht chaotisch, sondern unterliegt den Prinzipien der ökologischen Komplementarität, der ökologischen Konformität (Kongruenz) und anderen Gesetzmäßigkeiten. Im Rahmen der Ökodynamik von Yu. Goldsmith ist dies ihr drittes Gesetz – das Prinzip der ökologischen Ordnung oder des ökologischen Mutualismus, das eine globale Eigenschaft aufgrund des Einflusses des Ganzen auf seine Teile und den umgekehrten Einfluss differenzierter Teile auf die anderen angibt Entwicklung des Ganzen usw., was insgesamt zur Erhaltungsstabilität der gesamten Biosphäre führt. Gegenseitige Hilfe im Rahmen der ökologischen Ordnung oder systemischer Mutualismus wird durch das Gesetz der Ordnung der Raumfüllung und der räumlich-zeitlichen Gewissheit bekräftigt: Die Raumfüllung innerhalb eines natürlichen Systems ist aufgrund der Interaktion zwischen seinen Subsystemen geordnet Auf diese Weise können die homöostatischen Eigenschaften des Systems mit minimalen Widersprüchen zwischen den darin enthaltenen Teilen realisiert werden. Aus diesem Gesetz folgt, dass die langfristige Existenz von für die Natur „unnötigen“ Unfällen, einschließlich ihr fremder menschlicher Schöpfungen, unmöglich ist. Zu den Regeln der wechselseitigen Systemordnung in der Biosphäre gehört auch das Prinzip der Systemkomplementarität, das besagt, dass die Teilsysteme eines natürlichen Systems in ihrer Entwicklung eine Voraussetzung für die erfolgreiche Entwicklung und Selbstregulierung anderer Teilsysteme desselben Systems darstellen. Das vierte Gesetz der Ökodynamik von Yu. Goldsmith umfasst das Gesetz der Selbstkontrolle und Selbstregulierung von Lebewesen: Lebewesen und Systeme unter dem kontrollierenden Einfluss von Lebewesen sind in ihrem Prozess zur Selbstkontrolle und Selbstregulierung fähig Anpassung an Veränderungen in der Umwelt. In der Biosphäre erfolgen Selbstkontrolle und Selbstregulierung im Rahmen von Kaskaden- und Kettenprozessen allgemeiner Interaktion – während des Kampfes um die Existenz natürlicher Selektion (im weitesten Sinne dieses Konzepts), der Anpassung von Systemen und Subsystemen, der breiten Koevolution , usw. Darüber hinaus führen alle diese Prozesse zu positiven Ergebnissen „aus Sicht“ der Natur – der Erhaltung und Entwicklung der Ökosysteme der Biosphäre und dieser als Ganzes. Das Bindeglied zwischen Verallgemeinerungen struktureller und evolutionärer Natur ist die Regel der automatischen Aufrechterhaltung des globalen Lebensraums: Lebewesen erhält im Zuge der Selbstregulierung und Interaktion mit abiotischen Faktoren autodynamisch eine für ihre Entwicklung geeignete Lebensumgebung aufrecht. Dieser Prozess wird durch Veränderungen im kosmischen und globalen Maßstab der Ökosphäre begrenzt und findet in allen Ökosystemen und Biosystemen des Planeten als Kaskade der Selbstregulierung statt, die einen globalen Maßstab erreicht. Die Regel der automatischen Erhaltung des globalen Lebensraums ergibt sich aus den biogeochemischen Prinzipien von V.I. Wernadskij stellt die Regeln zur Erhaltung von Artenlebensräumen und relativer innerer Konsistenz dar und dient als Konstante für das Vorhandensein konservativer Mechanismen in der Biosphäre und bestätigt gleichzeitig die Regel der systemdynamischen Komplementarität. Der kosmische Einfluss auf die Biosphäre wird durch das Gesetz der Brechung kosmischer Einflüsse belegt: Kosmische Faktoren, die einen Einfluss auf die Biosphäre und insbesondere auf ihre Unterteilungen haben, unterliegen Veränderungen durch die Ökosphäre des Planeten und daher in Bezug auf Stärke und Zeit , können Erscheinungen abgeschwächt und verschoben werden oder sogar ganz ihre Wirkung verlieren. Die Verallgemeinerung hier ist wichtig, da es häufig zu synchronen Auswirkungen der Sonnenaktivität und anderer kosmischer Faktoren auf die Ökosysteme der Erde und die darin lebenden Organismen kommt. Es ist zu beachten, dass für viele Prozesse auf der Erde und in ihrer Biosphäre, obwohl sie dem Einfluss des Weltraums unterliegen, Zyklen der Sonnenaktivität mit Intervallen von 1850, 600, 400, 178, 169, 88, 83, 33, 22, 16 angenommen werden , 11, 5 (11 ,1), 6,5 und 4,3 Jahre müssen die Biosphäre selbst und ihre Teilbereiche nicht unbedingt in allen Fällen mit der gleichen Zyklizität reagieren. Die kosmischen Einflüsse des Biosphärensystems können ganz oder teilweise blockiert werden. Abschluss Ökosysteme sind die grundlegende Funktionseinheit in der Ökologie, da sie Organismen und die unbelebte Umwelt umfassen – Komponenten, die sich gegenseitig in ihren Eigenschaften und den notwendigen Bedingungen für die Erhaltung des Lebens in der auf der Erde existierenden Form beeinflussen. Die Kombination einer bestimmten physikalisch-chemischen Umgebung (Biotop) mit einer Gemeinschaft lebender Organismen (Biozönose) bildet ein Ökosystem. Es gibt Mikroökosysteme, Mesoökosysteme und das globale Ökosystem – die Biosphäre. Ökosysteme sind nicht ungeordnet verstreut; im Gegenteil, sie sind in ziemlich regelmäßige Zonen gruppiert, sowohl horizontal (in der Breite) als auch vertikal (in der Höhe). Die wichtigste Energiequelle für Ökosysteme ist die Sonne. Die auf die Erdoberfläche fallende Sonnenstrahlung beträgt 1,54 Millionen EJ pro Jahr. Der Großteil der Sonnenenergie, die die Planetenoberfläche erreicht, wird direkt in Wärme umgewandelt und erwärmt so Wasser oder Boden, was wiederum die Luft erwärmt. Ökosysteme existieren aufgrund der umweltfreundlichen und nahezu ewigen Sonnenenergie, deren Menge relativ konstant und reichlich vorhanden ist. Sonnenenergie auf der Erde verursacht zwei Stoffkreisläufe: große oder geologische und kleine, biologische (biotische). Beide Kreisläufe sind miteinander verbunden und stellen gewissermaßen einen einzigen Prozess dar. Die Existenz biogeochemischer Kreisläufe bzw. biogeochemischer Kreisläufe schafft die Möglichkeit zur Selbstregulierung (Homöostase) des Systems, was dem Ökosystem Stabilität verleiht: eine erstaunliche Konstanz des prozentualen Gehalts verschiedener Elemente. Es gibt eine ganze Reihe von Stoffkreisläufen. Der bedeutendste Kreislauf auf der Erde hinsichtlich der übertragenen Massen und des Energieverbrauchs ist der planetarische Wasserkreislauf oder Wasserkreislauf. Der biotische (biologische) Kreislauf bezeichnet den Stoffkreislauf zwischen Boden, Pflanzen, Tieren und Mikroorganismen. Von allen biogeochemischen Kreisläufen ist der Kohlenstoffkreislauf zweifellos der intensivste. Der größte Teil des in geologischen Epochen erzeugten Sauerstoffs blieb nicht in der Atmosphäre, sondern wurde von der Lithosphäre in Form von Karbonaten, Sulfaten, Eisenoxiden usw. fixiert. Diese Masse beträgt 590 x 1014 Tonnen gegenüber 39 x 1014 Tonnen Sauerstoff, der in der Biosphäre zirkuliert in Form von Gas oder Sulfaten, gelöst in kontinentalen und ozeanischen Gewässern. Stickstoff ist ein essentielles biogenes Element, da er Bestandteil von Proteinen und Nukleinsäuren ist. Der Stickstoffkreislauf ist einer der komplexesten, da er sowohl Gas- als auch Mineralphasen umfasst und gleichzeitig die idealsten Kreisläufe darstellt. Der Phosphorkreislauf in der Biosphäre ist mit Stoffwechselprozessen bei Pflanzen und Tieren verbunden. Dieses wichtige und notwendige Element des Protoplasmas, das in Landpflanzen und Algen zu 0,01–0,1 %, bei Tieren zu 0,1 % bis zu mehreren Prozent enthalten ist, zirkuliert und wandelt sich allmählich von organischen Verbindungen in Phosphate um, die von Pflanzen wieder genutzt werden können. Es gibt zahlreiche gasförmige Schwefelverbindungen, wie zum Beispiel Schwefelwasserstoff H2S und Schwefeldioxid SO2. Die Aufrechterhaltung der lebenswichtigen Aktivität von Organismen und der Stoffzirkulation in Ökosystemen, d. h. die Existenz von Ökosystemen, hängt vom ständigen Energiefluss ab, den alle Organismen für ihre lebenswichtigen Funktionen und Selbstreproduktion benötigen. Im Gegensatz zu Stoffen, die kontinuierlich durch verschiedene Blöcke des Ökosystems zirkulieren, die jederzeit wiederverwendet werden können und in den Kreislauf gelangen, ist Energie einmalig nutzbar, d. h. es gibt einen linearen Energiefluss durch das Ökosystem. Innerhalb eines Ökosystems werden energiehaltige Substanzen von autotrophen Organismen erzeugt und dienen Heterotrophen als Nahrung. Nahrungsverbindungen sind Mechanismen zur Energieübertragung von einem Organismus auf einen anderen. Innerhalb jedes Ökosystems haben Nahrungsnetze eine genau definierte Struktur, die durch die Art und Anzahl der Organismen gekennzeichnet ist, die auf jeder Ebene der verschiedenen Nahrungsketten vertreten sind. Die Bildung von Ökosystemen ist ein dynamischer Prozess. In Ökosystemen kommt es ständig zu Veränderungen im Zustand und in der Lebensaktivität ihrer Mitglieder sowie im Verhältnis der Populationen. Die vielfältigen Veränderungen, die in jeder Gemeinschaft stattfinden, lassen sich in zwei Haupttypen einteilen: zyklische und progressive. Die Sukzession ist ein natürlicher, gezielter Prozess und die Veränderungen, die in der einen oder anderen Phase auftreten, sind charakteristisch für jede Gemeinschaft und hängen nicht von ihrer Artenzusammensetzung oder geografischen Lage ab. Das globale Ökosystem ist die Biosphäre. Wie bereits erwähnt, wird die Biosphäre in Geobiosphäre, Hydrobiosphäre und Aerobiosphäre unterteilt. Die Biosphäre wird in größerem Maße nicht durch äußere Faktoren, sondern durch innere Muster geformt. Die wichtigste Eigenschaft der Biosphäre ist die Wechselwirkung von lebenden und nicht lebenden Dingen, die sich im Gesetz der biogenen Migration von Atomen von V.I. widerspiegelt. Wernadski. Die auf der Erde existierenden Ökosysteme sind vielfältig. Es gibt Mikroökosysteme (zum Beispiel der Stamm eines verrottenden Baumes), Mesoökosysteme (Wald, Teich usw.), Makroökosysteme (Kontinent, Ozean usw.) und die globale Biosphäre. Große terrestrische Ökosysteme werden Biome genannt. Jedes Biom enthält eine Reihe kleinerer, miteinander verbundener Ökosysteme. Es gibt verschiedene Klassifizierungen von Ökosystemen: immergrüner tropischer Regenwald; Wüste: grasig und strauchig; tropische Graslinsen und Savanne; gemäßigte Steppe; gemäßigter Laubwald: boreale Nadelwälder. Tundra: arktisch und alpin. Arten von Süßwasserökosystemen: Band (stilles Wasser): Seen, Teiche usw.; Lotisch (fließendes Wasser): Flüsse, Bäche usw.; Feuchtgebiete: Sümpfe und sumpfige Wälder; Arten von Meeresökosystemen: offenes Meer (pelagisch); Gewässer des Festlandsockels (Küstengewässer); Auftriebsgebiete (fruchtbare Gebiete mit produktiver Fischerei); Flussmündungen (Küstenbuchten, Meerengen, Flussmündungen, Salzwiesen usw.). Jedes Ökosystem besteht aus zwei Hauptkomponenten: Organismen und Faktoren aus ihrer unbelebten Umgebung. Die Gesamtheit der Organismen (Pflanzen, Tiere, Mikroben) wird als Biota eines Ökosystems bezeichnet. Die Art der Interaktion zwischen verschiedenen Kategorien von Organismen ist ihre biotische Struktur. Sonnenenergie auf der Erde verursacht zwei Stoffkreisläufe: einen großen oder geologischen, der sich am deutlichsten im Wasserkreislauf und der atmosphärischen Zirkulation manifestiert, und einen kleinen, biologischen (biotischen), der sich auf der Grundlage eines großen entwickelt und aus einem kontinuierlichen, zyklisch, aber zeitlich und räumlich ungleichmäßig und begleitet von mehr oder weniger erheblichen Verlusten bei der natürlichen Umverteilung von Materie, Energie und Information innerhalb von Ökosystemen verschiedener Organisationsebenen. Beide Kreisläufe sind miteinander verbunden und stellen gewissermaßen einen einzigen Prozess dar. Das Zusammenspiel abiotischer Faktoren und lebender Organismen des Ökosystems geht mit einem kontinuierlichen Stoffkreislauf zwischen Biotop und Biozönose in Form abwechselnder organischer und mineralischer Verbindungen einher. Der Austausch chemischer Elemente zwischen lebenden Organismen und der anorganischen Umgebung, der in verschiedenen Phasen innerhalb eines Ökosystems abläuft, wird als biogeochemischer Kreislauf oder biogeochemischer Kreislauf bezeichnet. Die Existenz solcher Zyklen schafft die Möglichkeit zur Selbstregulierung (Homöostase) des Systems, was dem Ökosystem Stabilität verleiht: eine erstaunliche Konstanz des Anteils verschiedener Elemente. Dabei gilt das Prinzip der Funktionsweise von Ökosystemen: Die Gewinnung von Ressourcen und die Entsorgung von Abfällen erfolgen im Rahmen des Kreislaufs aller Elemente. Liste der verwendeten Literatur 1.Bigon M.I. Ökologie, Individuen, Populationen und Gemeinschaften. M.: 1989. - 290 S. 2.Beysenova A.S., Shildebaev Zh.B., Sautbaeva G.Z. Ökologie. Almaty: „Gylym, 2001. – 201 S. 3.Akimova T.A., Khaskin V.V. Ökologie. M.: UNITY Publishing House, 1998. - 233 S. .Gorelov A.A. Ökologie. Vorlesungskurs. M.: "Center" 1997. - 280 S. .Stadnitsky G.V., Rodionov A.I. Ökologie. M.: UNITY Publishing House, 1996. - 272 S. .Sadanov A.K., Svanbaeva Z.S., Ökologie. Almaty: „Agrouniversity“, 1999. – 197 S. .Shilov I.A. Ökologie. M.: Höhere Schule, 2000. - 348 S. .Reimers N.F. Ökologie (Theorie, Gesetze, Regeln, Prinzipien und Hypothesen). M.: „Junges Russland“, 1994. - 260 S. .Tsvetkova L.I., Alekseev M.I., Usenov B.P. und andere. Ökologie. Lehrbuch für technische Hochschulen. M.; ASV, St. Petersburg: Khimizdat, 1999, 185 S. .Girusov E.V. Ökologie und Ökonomie des Umweltmanagements. M.: Recht und Gesetz, EINHEIT. 1998 – 232 S. .Wronski V.A. Angewandte Ökologie: Lehrbuch. Nutzen. Rostow am Don: 1995. - 197 S. .Budyko M.I. Globale Ökologie. M.: Mysl, 1977.-248 S. .Alekseenko V.A. Umweltgeochemie. Lehrbuch. M.: Logos, 2000 - 410 S. .Petrov K.M. Allgemeine Ökologie von S-P. Lernprogramm. "Chemie", 1997. - 218 S. .Andersen J.M. Ökologie und Umweltwissenschaften. Biosphäre, Ökosystem, Mensch. L., 1985. 376 S. .Girenok F.I. Ökologie. Zivilisation. Noosphere.M.: 191987. - 281 S. .Auf der Suche nach Balance: Ökologie im System gesellschaftspolitischer Prioritäten M.: 1992. - 427 S. .Geschichte der Interaktion zwischen Gesellschaft und Natur: Faktoren und Konzept. M.: ANSSSR 1990. .Stepanovskikh A.S. Angewandte Ökologie: Umweltschutz. Lehrbuch für Universitäten. M.: EINHEIT DANA. 2003. 751 S. Ähnliche Werke wie – Stoffkreislauf und Energieflüsse in Ökosystemen
Die Begriffe „Stofffluss“ und „Energiefluss“ sollten klar definiert werden. Der Stofffluss ist die Bewegung in Form chemischer Elemente und ihrer Verbindungen vom Erzeuger zum Zersetzer (durch oder ohne Verbraucher). Unter Energiefluss versteht man die Übertragung von Energie in Form chemischer Bindungen organischer Verbindungen (Nahrung) entlang der Nahrungskette von einer trophischen Ebene zur anderen (höheren).
Dabei ist zu beachten, dass die zugeführte Energie im Gegensatz zu Stoffen, die ständig durch verschiedene Blöcke des Ökosystems zirkulieren und immer wieder in den Kreislauf eintreten können, nur einmal genutzt werden kann.
Als universelles Naturphänomen wird der einseitige Energiefluss durch die Gesetze der Thermodynamik bestimmt. Nach dem ersten Hauptsatz kann Energie von einer Form (der Energie des Lichts) in eine andere (die potentielle Energie der Nahrung) übergehen, sie entsteht jedoch nie wieder und verschwindet nicht spurlos.
Der zweite Hauptsatz der Thermodynamik besagt, dass es keinen einzigen Prozess geben kann, der mit der Umwandlung von Energie verbunden ist, ohne dass ein Teil davon verloren geht. Aus diesem Grund kann beispielsweise eine Umwandlung von Nahrung in die Substanz, aus der der Körper des Organismus besteht, nicht mit 100 % Effizienz erfolgen.
Somit wird das Funktionieren aller Ökosysteme durch einen konstanten Energiefluss bestimmt, der für alle Organismen zur Aufrechterhaltung ihrer Existenz und Selbstreproduktion notwendig ist.
Auch in Ökosystemen gibt es Konkurrenzbeziehungen. In diesem Zusammenhang ist das Gesetz der Energiemaximierung (G. Odum – E. Odum) von großem Interesse: Im Wettbewerb mit anderen Ökosystemen überlebt dasjenige, das am besten zur Energieversorgung beiträgt und seine maximale Menge am effizientesten nutzt (Konserven). Laut Gesetz schafft das System zu diesem Zweck: 1) Akkumulatoren (Speicher) hochwertiger Energie (z. B. Fettreserven); 2) verbraucht eine bestimmte Menge der angesammelten Energie, um die Versorgung mit neuer Energie sicherzustellen; 3) sorgt für die Zirkulation verschiedener Substanzen; 4) schafft Regulierungsmechanismen, die die Stabilität des Systems und seine Fähigkeit zur Anpassung an sich ändernde Bedingungen unterstützen; 5) stellt den Austausch mit anderen Systemen her, die notwendig sind, um den Bedarf an besonderen Energiearten zu decken.
Es ist notwendig, einen wichtigen Umstand hervorzuheben: Das Gesetz der Energiemaximierung gilt auch in Bezug auf Informationen und kann daher (nach N. F. Reimers) auch als das Gesetz der Energie- und Informationsmaximierung betrachtet werden: das System, das am meisten dazu beiträgt Empfang, Produktion und effektive Nutzung von Energie und Informationen.
Beachten wir, dass die maximale Versorgung mit einer Substanz als solche noch keine Garantie für den Erfolg des Systems im Wettbewerb mit anderen ähnlichen Systemen darstellt.
Es wurde bereits früher darauf hingewiesen, dass zwischen den Organismen der Biozönose starke Nahrungsbeziehungen bzw. eine Nahrungskette entstehen und etabliert werden. Letzteres besteht aus drei Hauptgliedern: Produzenten, Konsumenten und Zersetzern.
Nahrungsketten, die mit photosynthetischen Organismen beginnen, werden als Weideketten (oder Weideketten) bezeichnet, und Ketten, die mit abgestorbenem Pflanzenmaterial, Kadavern und tierischen Exkrementen beginnen, werden als Detritalketten bezeichnet.
Die Position jedes Glieds in der Nahrungskette wird als trophische Ebene bezeichnet. es zeichnet sich durch unterschiedliche Intensitäten des Stoff- und Energieflusses aus. Die erste trophische Ebene sind immer Produzenten; der zweite - pflanzenfressende Verbraucher; der dritte sind Fleischfresser, die sich von pflanzenfressenden Formen ernähren; vierte Ebene – Verzehr anderer Fleischfresser usw.
Es gibt Verbraucher erster, zweiter, dritter und vierter Ordnung, die unterschiedliche Ebenen der Nahrungskette besetzen (Abb. 9).
Reis. 9.
Es liegt auf der Hand, dass die Lebensmittelspezialisierung der Verbraucher dabei eine große Rolle spielt. Arten mit einem breiten Ernährungsspektrum können auf unterschiedlichen trophischen Ebenen in Nahrungsketten einbezogen werden. Die Ernährung eines Menschen umfasst beispielsweise sowohl pflanzliche Lebensmittel als auch das Fleisch von Pflanzenfressern und Fleischfressern. Daher fungiert es in verschiedenen Nahrungsketten als Verbraucher der Ordnungen I, II oder III.
Da bei der Übertragung von Energie von einer Ebene auf eine andere Energie verloren geht, kann die Energiekette nicht lang sein: Sie besteht normalerweise aus 4...6 Gliedern (Tabelle 1).
1. Typische Diagramme von Nahrungsketten (nach V. M. Ivonin, 1996)
Allerdings kommen solche Ketten in reiner Form in der Natur meist nicht vor, da die gleiche Art gleichzeitig in verschiedenen Gliedern vorkommen kann. Dies liegt daran, dass es in der Natur nur wenige Monophagen gibt; Oligophagen und Polyphagen kommen deutlich häufiger vor. Beispielsweise sind Raubtiere, die sich von verschiedenen Pflanzen- und Fleischfressern ernähren, Glieder in vielen Ketten. Infolgedessen werden in jeder Biozönose evolutionär Komplexe von Nahrungsketten gebildet, die ein einziges Ganzes darstellen. So entstehen Stromnetze, die sehr komplex sind.
Daraus können wir schließen, dass die Nahrungskette der Hauptkanal der Energieübertragung in der Gemeinschaft ist (zwischen Pflanzen – Produzenten, Tieren – Verbrauchern und Mikroorganismen – Zersetzern) (Abb. 10). Bereits im Diagramm wird deutlich, dass die Idee von Nahrungsketten und trophische Ebenen sind eher eine Abstraktion. Im Labor kann eine lineare Kette mit klar getrennten Ebenen erstellt werden. In der Natur gibt es jedoch tatsächlich trophische Netzwerke, in denen viele Populationen gleichzeitig mehreren trophischen Ebenen angehören. Derselbe Organismus konsumiert sowohl Tiere als auch Pflanzen; das Raubtier kann sich von Verbrauchern erster und zweiter Ordnung ernähren; viele Tiere fressen sowohl lebende als auch tote Pflanzen.
Aufgrund der Komplexität der trophischen Beziehungen hat der Verlust einer Art oft kaum Auswirkungen auf die Gemeinschaft. Andere „Benutzer“ beginnen, die Nahrung der ausgestorbenen Art zu konsumieren,

Reis. 10.
Die Arten, die sich davon ernähren, finden neue Nahrungsquellen: Im Allgemeinen bleibt das Gleichgewicht in der Gemeinschaft erhalten.
Die von den Produzenten aufgenommene Energie fließt durch die Nahrungsketten und wird nach und nach verbraucht. Am Ende einer Nahrungskette ist die Energiemenge immer geringer als am Anfang. Bei der Photosynthese binden Pflanzen im Durchschnitt nur etwa 1 % der auf sie treffenden Sonnenenergie. Ein Tier, das eine Pflanze gefressen hat, verdaut einen Teil der Nahrung nicht und scheidet sie in Form von Exkrementen aus. Normalerweise werden 20...60 % der pflanzlichen Nahrung verdaut; Die aufgenommene Energie wird zur Erhaltung des Lebens des Tieres genutzt. Die Funktion von Zellen und Organen geht mit der Freisetzung von Wärme einher, d. h. dadurch wird ein erheblicher Teil der Nahrungsenergie bald an die Umwelt abgegeben. Ein relativ kleiner Teil der Nahrung wird für den Aufbau von neuem Gewebe und die Bildung von Fettreserven verwendet. Als nächstes erhält ein Raubtier, das einen Pflanzenfresser gefressen hat und die dritte trophische Ebene darstellt, nur die Energie aus der von der Pflanze angesammelten Energie, die im Körper seiner Beute (zweite Ebene) in Form einer Zunahme der Biomasse zurückgehalten wird.
Es ist bekannt, dass in jeder Phase der Stoff- und Energieübertragung in der Nahrungskette etwa 90 % der Energie verloren gehen und nur etwa ein Zehntel davon an den nächsten Verbraucher gelangt, also die Energieübertragung in den Nahrungsverbindungen der Organismen gehorcht der „Zehn-Prozent-Regel“ (Lindemann-Prinzip). Beispielsweise beträgt die Energiemenge, die tertiäre Fleischfresser (fünfte trophische Ebene) erreicht, nur etwa 10 -4 der von den Produzenten aufgenommenen Energie. Dies erklärt die begrenzte Anzahl (5...6) der Glieder (Stufen) in der Nahrungskette, unabhängig von der Komplexität der Artenzusammensetzung der Biozönose.

Reis. elf.
Betrachtet man den Energiefluss in Ökosystemen, ist auch leicht zu verstehen, warum die Biomasse mit zunehmendem trophischen Niveau abnimmt. Hier manifestiert sich das dritte Grundprinzip der Funktionsweise von Ökosystemen: Je größer die Biomasse einer Population, desto niedriger sollte das trophische Niveau sein, das sie einnimmt, oder anders: Am Ende langer Nahrungsketten kann es keine große Biomasse geben.
Die drei oben aufgeführten Grundprinzipien der Ökosystemfunktion – der Nährstoffkreislauf, der Fluss der Sonnenenergie und die Abnahme der Biomasse mit zunehmendem trophischen Niveau – können in Form eines allgemeinen Diagramms dargestellt werden (Abb. 11). Wenn wir Organismen nach ihren Ernährungsverhältnissen ordnen und für jeden von ihnen den „Input“ und „Output“ von Energie und Nährstoffen angeben, wird deutlich, dass Nährstoffe innerhalb des Ökosystems kontinuierlich recycelt werden und der Energiefluss durch dieses hindurch verläuft.
Die Aufrechterhaltung der lebenswichtigen Aktivität von Organismen und der Stoffzirkulation in Ökosystemen, d. h. die Existenz von Ökosystemen, hängt vom ständigen Energiefluss ab, den alle Organismen für ihre lebenswichtigen Funktionen und Selbstreproduktion benötigen.
Im Gegensatz zu Stoffen, die kontinuierlich durch verschiedene Blöcke des Ökosystems zirkulieren, die jederzeit wiederverwendet werden können und in den Kreislauf gelangen, kann Energie nur einmal genutzt werden, d. h. es findet ein linearer Energiefluss durch das Ökosystem statt.
Der einseitige Energiezufluss als universelles Naturphänomen erfolgt aufgrund der Gesetze der Thermodynamik. Erstes Gesetz besagt, dass Energie von einer Form (z. B. Licht) in eine andere (z. B. die potenzielle Energie von Nahrungsmitteln) umgewandelt, aber nicht erzeugt oder zerstört werden kann. Zweites Gesetz besagt, dass es keinen einzigen Prozess geben kann, der mit der Energieumwandlung verbunden ist, ohne dass ein Teil davon verloren geht. Bei solchen Umwandlungen wird ein Teil der Energie in unzugängliche Wärmeenergie umgewandelt und geht somit verloren. Daher kann es nicht zu Umwandlungen beispielsweise von Nahrungssubstanzen in die Substanz, aus der der Körper des Organismus besteht, kommen, die mit 100-prozentiger Effizienz erfolgen.
Lebewesen sind somit Energiewandler. Und bei jeder Energieumwandlung geht ein Teil davon in Form von Wärme verloren. Letztendlich wird die gesamte Energie, die in den biotischen Kreislauf eines Ökosystems gelangt, als Wärme abgegeben. Lebende Organismen nutzen für ihre Arbeit nicht unbedingt Wärme als Energiequelle – sie nutzen Licht und chemische Energie.
Ökologische Pyramiden. Innerhalb jedes Ökosystems haben Nahrungsnetze eine genau definierte Struktur, die durch die Art und Anzahl der Organismen gekennzeichnet ist, die auf jeder Ebene der verschiedenen Nahrungsketten vertreten sind. Um die Beziehungen zwischen Organismen in einem Ökosystem zu untersuchen und grafisch darzustellen, verwenden sie in der Regel ökologische Pyramiden anstelle von Nahrungsnetzdiagrammen. Ökologische Pyramiden drücken die trophische Struktur eines Ökosystems in geometrischer Form aus. Sie sind in Form von Rechtecken gleicher Breite aufgebaut, die Länge der Rechtecke muss jedoch proportional zum Wert des Messobjekts sein. Von hier aus können Sie gelangen Pyramiden aus Zahlen, Biomasse und Energie.
Ökologische Pyramiden spiegeln die grundlegenden Merkmale jeder Biozönose wider, wenn sie ihre trophische Struktur zeigen:
Ihre Höhe ist proportional zur Länge der jeweiligen Nahrungskette, d. h. zur Anzahl der darin enthaltenen trophischen Ebenen;
Ihre Form spiegelt mehr oder weniger die Effizienz der Energieumwandlungen beim Übergang von einer Ebene zur anderen wider.
Zahlenpyramiden. Sie stellen die einfachste Annäherung an die Untersuchung der trophischen Struktur eines Ökosystems dar. In diesem Fall wird zunächst die Anzahl der Organismen in einem bestimmten Gebiet gezählt, nach trophischen Ebenen gruppiert und in Form eines Rechtecks dargestellt, dessen Länge (oder Fläche) proportional zur Anzahl der in einem bestimmten Gebiet lebenden Organismen ist ( oder in einem bestimmten Volumen, wenn es sich um ein aquatisches Ökosystem handelt). Als Grundregel gilt, dass es in jeder Umgebung mehr Pflanzen als Tiere, mehr Pflanzenfresser als Fleischfresser, mehr Insekten als Vögel usw. gibt.
Bevölkerungspyramiden spiegeln die Dichte der Organismen auf jeder trophischen Ebene wider. Die Konstruktion verschiedener Bevölkerungspyramiden ist sehr vielfältig. Oft stehen sie auf dem Kopf.
Beispielsweise gibt es in einem Wald deutlich weniger Bäume (Primärproduzenten) als Insekten (Pflanzenfresser).
Biomassepyramide. Spiegelt die Nahrungsbeziehungen im Ökosystem besser wider, da die Gesamtmasse der Organismen berücksichtigt wird (Biomasse) jede trophische Ebene. Die Rechtecke in den Biomassepyramiden stellen die Masse der Organismen auf jeder trophischen Ebene pro Flächen- oder Volumeneinheit dar. Die Form der Biomassepyramide ähnelt häufig der Form der Bevölkerungspyramide. Charakteristisch ist eine Abnahme der Biomasse auf jeder aufeinanderfolgenden trophischen Ebene.
Biomassepyramiden sowie Zahlen können nicht nur gerade, sondern auch umgekehrt sein. Umgekehrte Biomassepyramiden sind charakteristisch für aquatische Ökosysteme, in denen sich Primärproduzenten wie phytoplanktonische Algen sehr schnell teilen und ihre Konsumenten – zooplanktonische Krebstiere – viel größer sind, aber einen langen Reproduktionszyklus haben. Dies gilt insbesondere für Süßwasserumgebungen, in denen die Primärproduktivität durch mikroskopisch kleine Organismen bereitgestellt wird, deren Stoffwechselraten erhöht sind, d. h. die Biomasse ist gering, die Produktivität jedoch hoch.
Energiepyramide. Die grundlegendste Möglichkeit, Verbindungen zwischen Organismen auf verschiedenen trophischen Ebenen darzustellen, sind Energiepyramiden. Sie stellen die Energieumwandlungseffizienz und Produktivität von Nahrungsketten dar und werden durch Zählen der Energiemenge (kcal) ermittelt, die pro Flächeneinheit und Zeiteinheit angesammelt und von Organismen auf jeder trophischen Ebene verbraucht wird. Daher ist es relativ einfach, die in der Biomasse gespeicherte Energiemenge zu bestimmen, es ist jedoch schwieriger, die Gesamtenergiemenge abzuschätzen, die auf jeder trophischen Ebene absorbiert wird. Nachdem wir ein Diagramm erstellt haben (Abb. 12.28), können wir feststellen, dass Destruktoren, deren Bedeutung in der Biomassepyramide gering erscheint, und umgekehrt in der Bevölkerungspyramide; Sie erhalten einen erheblichen Teil der Energie, die durch das Ökosystem fließt. Darüber hinaus verbleibt nur ein Teil dieser Energie in den Organismen auf jeder trophischen Ebene des Ökosystems und wird in Biomasse gespeichert; der Rest wird zur Befriedigung der Stoffwechselbedürfnisse der Lebewesen verwendet: Existenzerhaltung, Wachstum, Fortpflanzung. Auch Tiere verbrauchen eine erhebliche Menge Energie für die Muskelarbeit.
R. Lindeman formulierte es erstmals 1942 Energiepyramidengesetz, was in Lehrbüchern oft als „Gesetz von 10 %“ bezeichnet wird. Nach diesem Gesetz von einem trophische Ebene der ökologischen Pyramide Im Durchschnitt gelangen nicht mehr als 10 % der Energie auf eine andere Ebene.
Verbraucher fungieren als steuerndes und stabilisierendes Glied im Ökosystem. Verbraucher erzeugen in der Volkszählung ein Spektrum an Vielfalt und verhindern so das Monopol der Dominanten. Regel zur Kontrolle des Verbraucherwerts kann zu Recht als sehr grundlegend angesehen werden. Nach kybernetischer Auffassung sollte das Kontrollsystem komplexer aufgebaut sein als das kontrollierte, dann wird der Grund für die Vielfalt der Verbrauchertypen klar. Die steuernde Bedeutung der Verbraucher hat auch eine energetische Grundlage. Der Energiefluss durch die eine oder andere trophische Ebene kann nicht absolut durch die Verfügbarkeit von Nahrungsmitteln in der zugrunde liegenden trophischen Ebene bestimmt werden. Es bleibt bekanntlich immer eine ausreichende „Reserve“ übrig, da die völlige Vernichtung von Lebensmitteln zum Tod der Verbraucher führen würde. Diese allgemeinen Muster werden im Rahmen von Populationsprozessen, Gemeinschaften, Ebenen der ökologischen Pyramide und Biozönosen insgesamt beobachtet.
Um Präsentationsvorschauen zu nutzen, erstellen Sie ein Google-Konto und melden Sie sich an: https://accounts.google.com
Folienunterschriften:
Kreislauf von Materie und Energie in der Natur
Der Stoffkreislauf ist der sich wiederholende Prozess der Umwandlung und Bewegung der Materie in der Natur, der mehr oder weniger zyklisch abläuft. Alle Stoffe auf unserem Planeten befinden sich im Umlauf. In der Natur gibt es zwei Hauptkreisläufe: Große (geologische) Kleine (biogeochemische)
Der Große Kreislauf der Stoffe Der Große Kreislauf dauert Millionen von Jahren und entsteht durch die Wechselwirkung der Sonnenenergie mit der Tiefenenergie der Erde. Verbunden mit geologischen Prozessen, der Bildung und Zerstörung von Gesteinen und der anschließenden Bewegung von Zerstörungsprodukten.
Kleiner Stoffkreislauf Der kleine Stoffkreislauf (biogeochemisch) findet innerhalb der Biosphäre auf der Ebene der Biozönose statt. Sein Wesen ist die Bildung lebender Materie aus anorganischen Verbindungen während des Prozesses der Photosynthese und die Umwandlung organischer Materie während der Zersetzung zurück in anorganische Verbindungen. Biogeochemische Kreisläufe – Wernadskij V.I.
Wasserkreislauf Tr Abfluss inf Wasserverdunstung Dampfkondensation Niederschlagsabfluss Transpiration Infiltration
Unter Transpiration versteht man den Prozess, bei dem sich Wasser durch eine Pflanze bewegt und über die äußeren Organe der Pflanze, wie Blätter, Stängel und Blüten, verdunstet. Wasser ist für das Pflanzenleben lebensnotwendig, doch nur ein kleiner Teil des über die Wurzeln zugeführten Wassers wird direkt für Wachstum und Stoffwechsel genutzt.
Der Wasserkreislauf
Wasserkreislauf Der größte Teil des Wassers konzentriert sich in den Ozeanen. Von ihrer Oberfläche verdunstendes Wasser versorgt natürliche und künstliche Landökosysteme. Je näher ein Gebiet am Meer liegt, desto mehr Niederschläge gibt es. Das Land gibt ständig Wasser an den Ozean zurück: Ein Teil der Feuchtigkeit verdunstet, am aktivsten in Wäldern, ein Teil wird von Flüssen gesammelt: Sie erhalten Regen- und Schmelzwasser. Der Feuchtigkeitsaustausch zwischen Meer und Land erfordert sehr hohe Energiekosten: Etwa 30 % der auf die Erde gelangenden Sonnenenergie werden dafür aufgewendet.
Einfluss des Menschen auf den Wasserkreislauf Der Wasserkreislauf in der Biosphäre befand sich vor der Entwicklung der Zivilisation im Gleichgewicht, d. h. Der Ozean erhielt so viel Wasser aus den Flüssen, wie er durch Verdunstung verbrauchte. Mit der Entwicklung der Zivilisation begann dieser Kreislauf gestört zu werden. Insbesondere Wälder verdunsten immer weniger Wasser, weil... ihre Fläche nimmt ab und die Bodenoberfläche nimmt im Gegenteil zu, weil Die Fläche der bewässerten Agrarflächen nimmt zu. landet. Die Flüsse der südlichen Regionen wurden flacher. Wasser verdunstet an der Meeresoberfläche schlechter, weil... Ein erheblicher Teil davon ist mit einem Ölfilm bedeckt. All dies verschlechtert die Wasserversorgung der Biosphäre.
Dürren kommen immer häufiger vor und es kommt zu Umweltkatastrophen. Beispielsweise dauerte in Afrika in der Sahelzone – einer Halbwüstenregion, die die Sahara von den nördlichen Ländern des Kontinents trennt – seit mehr als 35 Jahren eine katastrophale Dürre an. Süßwasser, das vom Land ins Meer und andere Gewässer zurückfließt, ist oft verschmutzt. Das Wasser vieler russischer Flüsse ist praktisch nicht mehr trinkbar. Der Anteil an Süßwasser, der lebenden Organismen zur Verfügung steht, ist recht gering, daher muss es sparsam genutzt und nicht verschmutzt werden! Jeder vierte Mensch auf der Erde hat keinen Mangel an sauberem Trinkwasser. In vielen Teilen der Welt gibt es nicht genügend Wasser für die industrielle Produktion und Bewässerung.
Verschiedene Komponenten der Hydrosphäre nehmen auf unterschiedliche Weise und mit unterschiedlicher Geschwindigkeit am Wasserkreislauf teil. Für die vollständige Erneuerung des Wassers in Gletschern dauert es 8000 Jahre, Grundwasser – 5000 Jahre, Ozean – 3000 Jahre, Boden – 1 Jahr. Atmosphärischer Dampf und Flusswasser erneuern sich innerhalb von 10–12 Tagen vollständig. Der Wasserkreislauf in der Natur dauert etwa 1 Million Jahre.
Sauerstoffkreislauf Sauerstoff ist eines der häufigsten Elemente in der Biosphäre. Der Sauerstoffgehalt in der Atmosphäre beträgt fast 21 %. Sauerstoff ist Bestandteil von Wassermolekülen und lebenden Organismen (Proteine, Fette, Kohlenhydrate, Nukleinsäuren). Sauerstoff wird von Produzenten (Grünpflanzen) produziert. Ozon spielt eine wichtige Rolle im Sauerstoffkreislauf. Die Ozonschicht befindet sich in einer Höhe von 20–30 km über dem Meeresspiegel. Der Sauerstoffgehalt in der Atmosphäre wird durch zwei Hauptprozesse beeinflusst: 1) Photosynthese 2) Zersetzung organischer Stoffe, bei denen diese verbraucht werden.
Der Sauerstoffkreislauf ist ein langsamer Prozess. Es dauert etwa 2000 Jahre, bis der gesamte Sauerstoff in der Atmosphäre vollständig erneuert ist. Zum Vergleich: Die vollständige Erneuerung des Kohlendioxids in der Atmosphäre erfolgt in etwa 3 Jahren. Sauerstoff wird von den meisten lebenden Organismen zur Atmung verwendet. Sauerstoff wird bei der Brennstoffverbrennung in Verbrennungsmotoren, in den Öfen von Wärmekraftwerken, in Flugzeug- und Raketentriebwerken usw. verwendet. Zusätzlicher anthropogener Verbrauch kann das Gleichgewicht des Sauerstoffkreislaufs stören. Bisher gleicht die Biosphäre menschliche Eingriffe aus: Verluste werden durch grüne Pflanzen ausgeglichen. Mit der weiteren Reduzierung der Waldfläche und der Verbrennung von immer mehr Brennstoffen wird der Sauerstoffgehalt in der Atmosphäre abnehmen.
ES IST WICHTIG!!! Wenn der Sauerstoffgehalt in der Luft auf 16 % sinkt, verschlechtert sich der Gesundheitszustand einer Person (besonders das Herz leidet), bei 7 % verliert eine Person das Bewusstsein und bei 3 % tritt der Tod ein.
Kohlenstoffzyklus
Kohlenstoffkreislauf Kohlenstoff ist die Grundlage organischer Verbindungen und kommt in Form von Proteinen, Fetten und Kohlenhydraten in allen lebenden Organismen vor. Kohlenstoff gelangt in Form von Kohlendioxid in die Atmosphäre. In der Atmosphäre, in der der Großteil des Kohlendioxids konzentriert ist, findet ständig ein Austausch statt: Pflanzen nehmen bei der Photosynthese Kohlendioxid auf und alle Organismen geben es bei der Atmung ab. Bis zu 50 % des Kohlenstoffs in Form von CO 2 werden durch Zersetzer – Bodenmikroorganismen – in die Atmosphäre zurückgeführt. Kohlenstoff verlässt den Kreislauf als Calciumcarbonat.
Menschlicher Einfluss auf den Kohlenstoffkreislauf Technogene menschliche Aktivitäten stören das natürliche Gleichgewicht des Kohlenstoffkreislaufs: 1) Bei der Verbrennung organischer Brennstoffe werden jährlich etwa 6 Milliarden Tonnen CO 2 in die Atmosphäre emittiert: a) Stromerzeugung in Wärmekraftwerken b ) Autoabgase 2) Waldzerstörung. In den letzten 100 Jahren ist der Kohlendioxidgehalt in der Atmosphäre stetig und schnell gestiegen. Kohlendioxid + Methan + Wasserdampf + Ozon + Stickoxide = Treibhausgas. Die Folge ist der Treibhauseffekt – die globale Erwärmung, die zu großen Naturkatastrophen führen kann.
Stickstoffkreislauf In freier Form ist Stickstoff ein Bestandteil der Luft – 78 %. Stickstoff ist eines der wichtigsten Elemente für das Leben von Organismen. Stickstoff ist Bestandteil aller Proteine. Das Stickstoffmolekül ist sehr stark, weshalb die meisten Organismen nicht in der Lage sind, Luftstickstoff aufzunehmen. Stickstoff wird von lebenden Organismen nur in Form von Verbindungen mit Wasserstoff und Sauerstoff aufgenommen. Die Fixierung von Stickstoff in chemischen Verbindungen erfolgt durch Vulkan- und Gewitteraktivität, vor allem aber durch die Aktivität von Mikroorganismen – Stickstofffixierern (stickstofffixierende Bakterien und Blaualgen).
Stickstoff gelangt in Form von Nitraten in die Wurzeln von Pflanzen, die für die Synthese organischer Stoffe (Proteine) verwendet werden. Tiere nehmen Stickstoff über pflanzliche oder tierische Nahrung auf. Durch den Abbau abgestorbener organischer Stoffe gelangt Stickstoff wieder in die Atmosphäre. Bodenbakterien zersetzen Proteine in anorganische Substanzen – Gase – Ammoniak, Stickoxide, die in die Atmosphäre gelangen. Stickstoff, der in Gewässer gelangt, passiert auch die Nahrungskette Pflanze-Tier-Mikroorganismus und gelangt wieder in die Atmosphäre.
Menschlicher Einfluss auf den Stickstoffkreislauf Durch menschliches Handeln wird das natürliche Gleichgewicht des Stickstoffkreislaufs gestört. Beim Pflügen von Land nimmt die Aktivität von Mikroorganismen - Stickstofffixierern - fast um das Fünffache ab, wodurch der Stickstoffgehalt im Boden abnimmt, was zu einer Verringerung der Bodenfruchtbarkeit führt. Daher führt der Mensch überschüssige Nitrate in den Boden ein, die in Mineraldüngern enthalten sind. Bei der Verbrennung und Verarbeitung von Gas, Öl und Kohle gelangen große Mengen Stickoxide in die Atmosphäre und fallen in Form von saurem Regen. Die Wiederherstellung des natürlichen Stickstoffkreislaufs ist durch eine Reduzierung der Produktion von Stickstoffdüngern, eine Reduzierung der industriellen Emissionen von Stickoxiden in die Atmosphäre usw. möglich.
Phosphorkreislauf
Im Gegensatz zu den Wasser-, Kohlenstoff-, Stickstoff- und Sauerstoffkreisläufen, die geschlossen sind, ist der Phosphorkreislauf offen, weil Phosphor bildet keine flüchtigen Verbindungen, die in die Atmosphäre gelangen. Phosphor ist in Gesteinen enthalten und gelangt von dort aus bei der natürlichen Zerstörung von Gesteinen oder bei der Ausbringung von Phosphordünger auf Felder in die Ökosysteme. Pflanzen nehmen anorganische Phosphorverbindungen auf und Tiere, die sich von diesen Pflanzen ernähren, reichern Phosphor in ihren Geweben an. Nach der Zersetzung toter Tier- und Pflanzenkörper wird nicht der gesamte Phosphor in den Kreislauf einbezogen. Ein Teil davon wird aus dem Boden in Gewässer (Flüsse, Seen, Meere) gespült und setzt sich am Boden ab. Mit den von Menschen gefangenen Fischen gelangt Phosphor in geringen Mengen an Land zurück.
Einfluss des Menschen auf den Phosphorkreislauf Der Transfer von Phosphor vom Land in die Ozeane hat unter menschlichem Einfluss deutlich zugenommen. Wenn Wälder zerstört und Böden gepflügt werden, erhöht sich das Volumen des Oberflächenwasserflusses, und außerdem gelangen Phosphordünger, die von Feldern in Flüsse und Seen ausgebracht werden. Da die Phosphorreserven an Land begrenzt sind und die Rückführung aus dem Meer schwierig ist, kann es in Zukunft zu einem Mangel an Phosphor in der Landwirtschaft kommen, der zu Ertragseinbußen (vor allem im Getreideanbau) führen wird.