RNR tipai ląstelėje ir jų sandara. Kas yra DNR ir RNR? DNR struktūra
Struktūra
Azoto bazės RNR gali sudaryti vandenilinius ryšius tarp citozino ir guanino, adenino ir uracilo bei tarp guanino ir uracilo. Tačiau galimos ir kitos sąveikos, pavyzdžiui, keli adeninai gali sudaryti kilpą arba kilpą, susidedančią iš keturių nukleotidų, kuriuose yra adenino ir guanino bazių pora. Svarbi struktūrinė RNR ypatybė, skirianti ją nuo DNR hidroksilo grupės buvimas 2" ribozės padėtyje, kuri leidžia RNR molekulei egzistuoti A, o ne B konformacijoje, dažniausiai stebimoje DNR. A forma turi gilų ir siaurą pagrindinį griovelį ir seklią bei plačią minorinę Antroji 2 colių hidroksilo grupės pasekmė yra ta, kad ji yra plastiška. ty RNR molekulės dalys, kurios nedalyvauja formuojant dvigubą spiralę, gali chemiškai atakuoti kitus fosfato ryšius ir nutrūkti. juos žemyn „darbinė“ vienos grandinės RNR molekulės forma, kaip ir baltymų, dažnai turi tretinę struktūrą. Tretinė struktūra susidaro remiantis antrinės struktūros elementais, kuriuos sudaro vandenilio ryšiai vienoje molekulėje. Yra keli antrinės struktūros elementų tipai – stiebo kilpos, kilpos ir pseudoknotai. Dėl didelio galimų bazių porų skaičiaus nuspėti antrinę RNR struktūrą yra daug sunkesnė užduotis nei nuspėti antrinę baltymų struktūrą, tačiau dabar yra veiksmingų programų, tokių kaip mfold.
RNR molekulių funkcijos priklausomybės nuo jų antrinės struktūros pavyzdys yra vidinės ribosomų įėjimo vietos (IRES). IRES yra struktūra 5" pasiuntinio RNR gale, užtikrinanti ribosomos prijungimą, apeinant įprastą baltymų sintezės inicijavimo mechanizmą, kuriam reikalinga speciali modifikuota bazė (dangtelis) 5" gale ir baltymų inicijavimo faktoriai. . Iš pradžių IRES buvo aptiktas virusinėse RNR, tačiau vis daugėja įrodymų, kad ląstelių mRNR streso sąlygomis taip pat naudoja nuo IRES priklausomą inicijavimo mechanizmą.
Daugelis RNR tipų, tokių kaip rRNR ir snRNR, ląstelėse veikia kaip kompleksai su baltymais, kurie asocijuojasi su RNR molekulėmis po jų sintezės arba (eukariotuose) eksporto iš branduolio į citoplazmą. Tokie RNR-baltymų kompleksai vadinami ribonukleoproteinų kompleksais arba ribonukleoproteinais.
Palyginimas su DNR
Yra trys pagrindiniai skirtumai tarp DNR ir RNR:
DNR yra cukraus dezoksiribozės, RNR yra ribozės, kuri, palyginti su dezoksiriboze, turi papildomą hidroksilo grupę. Ši grupė padidina molekulės hidrolizės tikimybę, tai yra, sumažina RNR molekulės stabilumą.
Nukleotidas, papildantis adeniną RNR, nėra timinas, kaip DNR, o uracilas yra nemetilinta timino forma.
DNR egzistuoja dvigubos spiralės, susidedančios iš dviejų atskirų molekulių, pavidalu. RNR molekulės vidutiniškai yra daug trumpesnės ir daugiausia viengrandinės.
Biologiškai aktyvių RNR molekulių, įskaitant tRNR, rRNR, snRNR ir kitas baltymų nekoduojančias molekules, struktūrinė analizė parodė, kad jos susideda ne iš vienos ilgos spiralės, o iš daugybės trumpų spiralių, esančių arti viena kitos ir sudarančių kažką panašaus į tretinė baltymo struktūra. Dėl to RNR gali katalizuoti chemines reakcijas, pavyzdžiui, ribosomos peptidiltransferazės centras, dalyvaujantis formuojant peptidinius ryšius baltymuose, susideda tik iš RNR.
Messenger (informacinė) RNR – RNR, kuri tarnauja kaip tarpininkas perduodant DNR užkoduotą informaciją į ribosomas, molekulines mašinas, kurios sintetina baltymus gyvame organizme. MRNR kodavimo seka nustato baltymo polipeptidinės grandinės aminorūgščių seką. Tačiau didžioji dauguma RNR nekoduoja baltymų. Šios nekoduojančios RNR gali būti transkribuotos iš atskirų genų (pvz., ribosomų RNR) arba gautos iš intronų. Klasikiniai, gerai ištirti nekoduojančių RNR tipai yra perdavimo RNR ( tRNR) ir rRNR, kurios dalyvauja vertimo procese. Taip pat yra RNR klasių, atsakingų už genų reguliavimą, mRNR apdorojimą ir kitus vaidmenis. Be to, yra nekoduojančių RNR molekulių, kurios gali katalizuoti chemines reakcijas, tokias kaip RNR molekulių pjaustymas ir surišimas. Analogiškai su baltymais, galinčiais katalizuoti chemines reakcijas – fermentus (fermentus), katalizinės RNR molekulės vadinamos ribozimais.
Dalyvauja transliacijoje
Informacija apie baltymo aminorūgščių seką yra mRNR. Trys iš eilės einantys nukleotidai (kodonas) atitinka vieną aminorūgštį. Eukariotinėse ląstelėse transciribuota pirmtako mRNR arba pre-mRNR yra apdorojama, kad susidarytų brandi mRNR. Apdorojimas apima nekoduojančių baltymų sekų (intronų) pašalinimą. Po to mRNR eksportuojama iš branduolio į citoplazmą, kur ji yra prijungta prie ribosomų, kurios verčia mRNR naudojant tRNR, sujungtas su aminorūgštimis.
Ląstelėse su branduoliais (bakterijose ir archėjose) ribosomos gali prisijungti prie mRNR iš karto po RNR dalies transkripcijos. Tiek eukariotuose, tiek prokariotuose mRNR gyvavimo ciklas baigiasi kontroliuojamu fermentų ribonukleazių sunaikinimu.
Transportas (tRNR) yra mažos molekulės, susidedančios iš maždaug 80 nukleotidų, turinčios konservatyvią tretinę struktūrą. Jie perneša specifines aminorūgštis į peptidinių jungčių sintezės vietą ribosomoje. Kiekvienoje tRNR yra vieta aminorūgščių prijungimui ir antikodonas, skirtas atpažinti ir prijungti prie mRNR kodonų. Antikodonas sudaro vandenilinius ryšius su kodonu, dėl kurio tRNR yra tokia padėtis, kuri palengvina peptidinio ryšio tarp paskutinės susidariusio peptido aminorūgšties ir prie tRNR prijungtos aminorūgšties susidarymą.
Ribosominė RNR (rRNR) yra katalizinis ribosomų komponentas. Eukariotinėse ribosomose yra keturių tipų rRNR molekulės: 18S, 5.8S, 28S ir 5S. Trys iš keturių rRNR tipų yra sintetinami branduolyje. Citoplazmoje ribosominės RNR jungiasi su ribosomų baltymais ir sudaro nukleoproteiną, vadinamą ribosoma. Ribosoma prisijungia prie mRNR ir sintetina baltymą. rRNR sudaro iki 80% RNR, esančios eukariotinės ląstelės citoplazmoje.
Neįprastas RNR tipas, veikiantis kaip tRNR ir mRNR (tmRNR), randamas daugelyje bakterijų ir plastidų. Kai ribosoma sustoja prie defektinių mRNR be stop kodonų, tmRNR prijungia mažą peptidą, kuris nukreipia baltymą į skaidymą.
RNR FUNKCIJOS
Yra įvairių tipų RNR. Messenger (informacinė) RNR – RNR, kuri tarnauja kaip tarpininkas perduodant DNR užkoduotą informaciją į ribosomas, molekulines mašinas, kurios sintetina baltymus gyvame organizme. Perkelkite RNR (tRNR) ir rRNR, kurios dalyvauja vertimo procese. Taip pat yra RNR klasių, atsakingų už genų reguliavimą, mRNR apdorojimą ir kitus vaidmenis. Be to, yra nekoduojančių RNR molekulių, kurios gali katalizuoti chemines reakcijas, tokias kaip RNR molekulių pjaustymas ir surišimas.
Apibendrinant visa tai, kas išdėstyta pirmiau, galima pastebėti, kad RNR funkcijos:
- paveldimos informacijos saugojimas (kai kuriems virusams);
- dalyvavimas vertimo ir transkripcijos (baltymų sintezės) procese;
- genų veiklos reguliavimas;
- katalizuoja daugybę cheminių reakcijų
ląstelėje yra trijų tipų RNR
Perneškite RNR (tRNR). Pernešimo RNR daugiausia randama ląstelės citoplazmoje. Funkcija yra perkelti aminorūgštis į ribosomas, į baltymų sintezės vietą. Iš bendro RNR kiekio ląstelėje t-RNR sudaro apie 10%.
Ribosominė RNR (r-RNR). Ribosominė RNR yra esminė ribosomos struktūros dalis. Iš viso ląstelėje esančio RNR kiekio r-RNR sudaro apie 90%.
Messenger RNR (i-RNR) arba pasiuntinio RNR (m-RNR). Esama branduolyje ir citoplazmoje. Jo funkcija yra perduoti informaciją apie baltymo struktūrą iš DNR į baltymų sintezės vietą ribosomose. mRNR sudaro maždaug 0,5–1% visos ląstelės RNR.
Yra RNR molekulėje vietoj timino. RNR nukleotiduose vietoj dezoksiribozės yra ribozės. RNR grandinėje nukleotidai yra sujungti kovalentiniais ryšiais tarp vieno nukleotido ribozės ir kito nukleotido fosforo rūgšties liekanos.
Organizme RNR randama kompleksų su baltymais – ribonukleoproteinų – pavidalu.
Yra žinomi 2 RNR molekulių tipai:
1) Dvigrandė RNR būdinga kai kuriems virusams – jie tarnauja paveldimos informacijos saugojimui ir atkūrimui (atlieka chromosomų funkcijas).
2) Daugumoje ląstelių vienos grandinės RNR vykdo informacijos apie aminorūgščių seką baltymuose perdavimą iš chromosomos į ribosomą.
Viengrandės RNR turi erdvinė organizacija: dėl azoto bazių sąveikos tarpusavyje, taip pat su cukraus-fosfato pagrindo fosfatais ir hidroksilais, grandinė susilanksto į kompaktišką rutulinio tipo struktūrą. Funkcija: perduoda informaciją iš chromosomos į ribosomas apie AK seką baltymuose, kurie turi būti sintetinami.
Yra keletas vienos grandinės RNR tipų, atsižvelgiant į jų funkciją arba vietą ląstelėje:
1. Ribosominė RNR (rRNR) sudaro didžiąją dalį RNR citoplazmoje (80-90%). Dydžiai 3000-5000 bazinių porų. Antrinė struktūra yra dviejų spiralių plaukų segtukų forma. r-RNR yra struktūrinis ribosomų komponentas – ląstelių organelių, kuriose vyksta baltymų sintezė. Ribosomos yra lokalizuotos citoplazmoje, branduolyje, mitochondrijose ir chloroplastuose. Jie susideda iš dviejų subvienetų – didelio ir mažo. Mažasis subvienetas susideda iš vienos rRNR molekulės ir 33 baltymų molekulių, didysis subvienetas – 3 rRNR molekulės ir 50 baltymų. Ribosominiai baltymai atlieka fermentines ir struktūrines funkcijas.
rRNR funkcijos:
1) konstrukcinis komponentas ribosomos- jų vientisumas būtinas baltymų biosintezei;
2) užtikrinti teisingą ribosomos prisijungimą prie m-RNR;
3) užtikrinti teisingą ribosomos prisijungimą prie t-RNR;
2. Matrica (mRNR) - 2-6% viso RNR kiekio.
Susideda iš sekcijų:
1) cistronai - nustato aminorūgščių seką baltymuose, kuriuos jie koduoja ir turi unikalią nukleotidų seką;
2) neverčiamos sritys yra molekulės galuose ir turi bendrus nukleotidų sudėties modelius.
Cap – speciali struktūra 5′ m-RNR gale – yra 7-metilguanozino trifosfatas, susidaręs fermentiniu būdu transkripcijos metu.
Dangtelio funkcijos:
1) apsaugo 5′ galą nuo skilimo egzonukleazėmis,
2) naudojamas specifiniam m-RNR atpažinimui vertimo metu.
Precistroninė netransliuota sritis – 3-15 nukleotidų. Funkcija: užtikrina teisingą m-RNR 5′ galo sąveiką su ribosoma.
Cistronas: yra inicijavimo ir pabaigos kodonai – specialios nukleotidų sekos, atsakingos už informacijos perdavimo iš tam tikro cistrono pradžią ir pabaigą.
Postcistronic netransliuota sritis – esanti 3′ gale, turi heksanukleotidą (dažnai AAUAAA) ir 20-250 adenilo nukleotidų grandinę. Funkcija – tarpląstelinio m-RNR stabilumo palaikymas.
3. Perneškite RNR (tRNR) - 15% visos RNR, susideda iš 70-93 nukleotidų porų. Funkcija: aminorūgšties perkėlimas į baltymų sintezės vietą jie „atpažįsta“ (pagal komplementarumo principą) mRNR sritį, atitinkančią perkeltą aminorūgštį. Kiekvienai iš 20 AA yra specifinės tRNR (paprastai daugiau nei viena). Visos tRNR turi sudėtingą struktūrą, pavaizduotą dobilo lapo pavidalu.

Dobilo lapą sudaro 5 skyriai:
1) 3′ galas - akceptoriaus šaka (AK liekana čia prijungta eterio jungtimi),
2) antikidoninė šaka – esanti priešais akceptoriaus vietą, susideda iš trijų nesuporuotų (turinčių laisvus ryšius) nukleotidų (antikodono) ir konkrečiai porų (antilygiagrečių, komplementarių) su m-RNR kodonu.
Kodonas- m-RNR 3 nukleotidų rinkinys (tripletas), kuris nustato šios aminorūgšties vietą susintetintoje polipeptidinėje grandinėje. Tai genetinio kodo vienetas, kurio pagalba visa genetinė informacija „įrašoma“ į DNR ir RNR molekules.
3) T-šaka (pseudouredino kilpa – yra pseudouredino) – atkarpa, kuri prisitvirtina prie ribosomos.
4) D-šaka (dehidrouredino kilpa – yra dehidrouredino) – sekcija, užtikrinanti sąveiką su aminorūgštį atitinkančiu fermentu aminoacil-tRNR sintetaze.
5) Papildoma nedidelė šaka. Funkcijos dar nebuvo ištirtos.
6) Branduolinė RNR (nRNR) – ląstelės branduolio komponentas. Mažas polimeras, stabilus, kurio vaidmuo vis dar neaiškus.
Visų rūšių RNR sintetinama ląstelės branduolyje ant DNR matricos, veikiant fermentams polimerazės. Tokiu atveju susidaro ribonukleotidų seka, kuri yra komplementari su dezoksiribonukleotidų seka DNR – tai yra transkripcijos procesas.
Visų tipų RNR yra skirtos išgauti informaciją apie baltymo struktūrą iš DNR ir užtikrinti baltymų biosintezę pagal šią informaciją. RNR yra viena polinukleotidų grandinė, sudaryta iš keturių pagrindinių ribonukleotidų tipų – AMP, GMP, CMP ir UMP. RNR būdingi smulkūs nukleotidai su neįprastomis azoto bazėmis – dihidrouracilu, 3-metiluracilu, 1-metilguaninu ir kt. (iki 50 rūšių). Ypač daug jų yra aminoacil-tRNR (iki 10 % visų nukleotidų). RNR adenino ir guanino kiekis neatitinka uracilo ir citozino kiekio.
Išskiriami šie RNR tipai:
Messenger arba pasiuntinio RNR(m- arba i-RNR), m.m. 25000-1000000 Taip, susideda iš 75-300 nukleotidų, susintetintų branduolyje iš pre-m-RNR; sudaro 5-7% visų ląstelių RNR. Pusinės eliminacijos laikas yra kelios minutės. Visų eukariotų mRNR 5' gale yra speciali struktūra, vadinama dangtelis. Dangtelis yra 7-metilguanozino trifosfatas. Prieš baigiantis transkripcijai, dangtelis susidaro fermentiniu būdu branduolyje. Manoma, kad dangtelis, viena vertus, apsaugo mRNR 5' galą nuo jo skilimo 5' egzonukleazėmis, kita vertus, yra naudojamas specifiniam atpažinimui vertimo sistemoje. Po dangteliu seka netransliuojamas skyrius, kuriame (nuo 3-15 nukleotidų prieš starto kodoną) yra rRNR sekai komplementari nukleotidų seka. Jo vaidmuo yra užtikrinti teisingą 5' galo sąveiką su ribosoma. Transliacijos skyrius baigiasi stop kodonas, dažnai seka heksanukleotidas AAAAAAA. Daugumoje mRNR 3' galas turi 20-250 adenilo nukleotidų poliadenilato grandinę, kuri nėra transkripcijos rezultatas, bet yra fermentiškai prisijungusi prie iRNR brendimo metu branduolyje. Daroma prielaida, kad poliadenilato seka yra atsakinga už mRNR intracelulinio stabilumo palaikymą ir lemia jos gyvavimo trukmę. Kodo elementas yra nukleotidų (kodono) tripletas, koduojantis aminorūgštį. Įrodyta, kad linijinėje iRNR molekulėje susidaro keli dvigrandžiai plaukų segtukai, kurių galuose yra transkripcijos pradžios ir pabaigos „ženklai“. Antrinėje struktūroje yra išlenkta grandinė; Remiantis kai kuriais duomenimis, tretinėje struktūroje polinukleotidų grandinė yra sujungta (žaizta) su transportinio baltymo informoferiu.
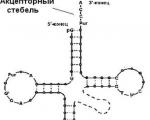
Perkelkite RNR ( tRNR ) – apie 15 proc. Pernešimo RNR turi mažą molekulinę masę (~ 25 000) ir yra tirpioje citoplazmos frakcijoje, atliekančios aminorūgščių pernešimo į baltymų sintezės vietą - ribosomą - funkciją. Sudėtyje yra apie 75 nukleotidai. Ląstelėje yra mažiausiai 20 tipų tRNR molekulių. Kiekvienas ar daugiau tRNR tipų atitinka vieną iš 20 aminorūgščių, reikalingų baltymų biosintezei. Visų tRNR antrinė struktūra primena " dobilo lapelis» (12.3 pav.) ir turi 4 pagrindines sekcijas. 1) Priimtojas vieta turi nukleotidų seką CCA 3" gale. Aminorūgščių karboksilo grupė pridedama prie adenozilo liekanos 3"-hidroksilo grupės. Pernešimo tRNR, susietos su aminorūgštimis, vadinamos aminoacil-tRNR(aatRNR). Jie atlieka adapterio funkcija verčiant trijų raidžių nukleorūgščių kodą į 20 raidžių aminorūgščių seką polipeptidinėje grandinėje. 2) Antikodono kilpa reikalingas prisijungti prie mRNR tripleto arba kodono. 3) Pseudouridilo kilpa(TψC) susideda iš 7 nukleotidų ir turi pseudouridino liekaną; tarnauja tRNR prijungimui prie ribosomos; 4) Dihidrouridino kilpa(D) susideda iš 8-12 nukleotidų liekanų. Reikalingas prisijungimui prie aminoacil-tRNR sintetazės, kuri dalyvauja atpažįstant savo tRNR aminorūgštimi. Tretinę struktūrą vaizduoja erdvinė struktūra alkūnės lenkimo (L formos) pavidalu.
Ryžiai. 12.3. tRNR struktūra
Ribosominės RNR (rRNR)– 80-85%, turi skirtingą ir žymiai didesnę molekulinę masę (35000-1000000, kas atitinka 100-3100 nukleotidų), yra struktūriniai ribosomų komponentai.
Ribosomos užtikrina specifinį kontaktą tarp iRNR ir tRNR, dėl to iš konkretaus geno nuskaityta nukleotidų seka paverčiama atitinkamo baltymo aminorūgščių seka. Žinduolių ribosomos susideda iš dviejų nukleoproteino subvienetų – didelio, kurio sedimentacijos konstanta yra 60S, ir mažo – 40S (prokariotuose – atitinkamai 50S ir 30S). 60S subvienetą sudaro 5S ribosominė RNR (rRNR), 5,8S rRNR ir 28S rRNR. Mažą 40S vienetą sudaro viena 18S rRNR ir apie 30 polipeptidinių grandinių. Visos ribosominės RNR, išskyrus 5S RNR, turi bendrą pirmtaką – 45S RNR, lokalizuotą branduolyje. Branduolėje labai metilintos ribosominės RNR yra supakuotos su ribosomų baltymais.
Prokariotinėse ribosomose ir mitochondrijų bei plastidų ribosomose yra mažiau komponentų, tačiau jos struktūriškai ir funkciškai labai panašios į eukariotus. Antrinė rRNR struktūra susidaro dėl trumpų dvigrandžių molekulės atkarpų – plaukų segtukų. Maždaug 2/3 rRNR yra suskirstyta į plaukų segtukus, 1/3 sudaro vienos grandinės sritys, kuriose gausu purino nukleotidų, prie kurių daugiausia jungiasi baltymai. Ribosominiai baltymai, kaip ir histonai, turi pagrindinį pobūdį ir atlieka tiek struktūrinį, tiek fermentinį vaidmenį.
Tarpininko, kurio funkcija yra paversti DNR saugomą paveldimą informaciją į darbinę formą, vaidmenį atlieka ribonukleino rūgštys – RNR.
Yra žinomos dviejų ir viengrandžių RNR molekulės. Dvigrandė RNR padeda saugoti ir atkurti kai kurių virusų paveldimą informaciją, t.y. jie atlieka chromosomų funkcijas. Viengrandės RNR neša informaciją apie aminorūgščių seką baltymuose iš chromosomos į jų sintezės vietą ir dalyvauja sintezės procesuose.
Skirtingai nuo DNR molekulių, ribonukleorūgštys yra pavaizduotos viena polinukleotidų grandine, kurią sudaro keturių tipų nukleotidai, kuriuose yra cukraus, ribozės, fosfato ir vienos iš keturių azoto bazių – adenino, guanino, uracilo arba citozino. RNR sintetinama ant DNR molekulių, naudojant RNR polimerazės fermentus, laikantis komplementarumo ir antiparaleliškumo principo, o uracilas papildo DNR adeniną RNR. Visą ląstelėje veikiančių RNR įvairovę galima suskirstyti į tris pagrindinius tipus: mRNR, tRNR, rRNR.
Matrica, arba informacija, RNR (mRNR arba mRNR). Transkripcija. Norint susintetinti nurodytų savybių baltymus, į jų konstravimo vietą siunčiamos „instrukcijos“ apie aminorūgščių įtraukimo į peptidinę grandinę tvarką. Ši instrukcija yra nukleotidų sekoje matrica, arba pasiuntinio RNR(mRNR, mRNR), susintetinti atitinkamose DNR skyriuose. iRNR sintezės procesas vadinamas transkripcija.
MRNR sintezė prasideda nuo RNR polimerazės atradimo specialioje DNR molekulėje, kuri nurodo vietą, kur prasideda transkripcija. propaguotojas Prisijungusi prie promotoriaus, RNR polimerazė išvynioja gretimą DNR spiralės posūkį. Šiuo metu dvi DNR grandinės išsiskiria, o vienoje iš jų fermentas sintezuoja mRNR. Ribonukleotidų surinkimas į grandinę vyksta atsižvelgiant į jų komplementarumą DNR nukleotidams, taip pat antiparaleliškai DNR šablono grandinės atžvilgiu. Dėl to, kad RNR polimerazė gali surinkti polinukleotidą tik nuo 5" galo iki 3" galo, tik viena iš dviejų DNR grandinių, būtent ta, kuri yra prieš fermentą 3 colių galu, gali būti šablonas. transkripcijai ( 3" → 5" Ši grandinė vadinama kodogeninis
tRNR- RNR, kurios funkcija yra transportuoti aminorūgštis į baltymų sintezės vietą. tRNR taip pat tiesiogiai dalyvauja pratęsiant polipeptidinę grandinę, jungdamosi (būdamos komplekse su aminorūgštimi) prie mRNR kodono ir suteikdamos kompleksinę konformaciją, reikalingą naujam peptidiniam ryšiui susidaryti.
Kiekviena aminorūgštis turi savo tRNR yra vienos grandinės RNR, tačiau savo funkcine forma ji turi "dobilo" arba "dobilo" konformaciją. Aminorūgštis kovalentiškai prijungta prie 3" molekulės galo, naudojant fermentą aminoacil-tRNR sintetazę, būdingą kiekvienam tRNR tipui. C vietoje yra aminorūgštį atitinkantis antikodonas.
(rRNR)- kelios RNR molekulės, kurios sudaro ribosomos pagrindą. Pagrindinė rRNR funkcija yra atlikti vertimo procesą – nuskaityti informaciją iš mRNR naudojant tRNR adapterio molekules ir katalizuoti peptidinių ryšių tarp aminorūgščių, prijungtų prie tRNR, susidarymą.
| Parametrų pavadinimas | Reikšmė |
| Straipsnio tema: | RNR tipai. |
| Rubrika (teminė kategorija) | Sportas |
Yra trys pagrindiniai RNR tipai, kurie skiriasi struktūra, molekulių dydžiu, vieta ląstelėje ir atliekamomis funkcijomis.
Ribosominės RNR (rRNR) sintetinami daugiausia branduolyje ir sudaro maždaug 85 % visos ląstelės RNR. Οʜᴎ yra ribosomų dalis ir dalyvauja formuojant aktyvųjį ribosomos centrą, kuriame vyksta baltymų biosintezės procesas.
Perneškite RNR (tRNR) susidaro DNR esančiame branduolyje, tada pereina į citoplazmą. Οʜᴎ sudaro apie 10% ląstelių RNR ir yra mažiausios RNR, susidedančios iš 70–100 nukleotidų. Kiekviena tRNR prijungia specifinę aminorūgštį ir perneša ją į polipeptido surinkimo vietą ribosomoje. Visos žinomos tRNR dėl viena kitą papildančios sąveikos sudaro antrinę struktūrą, panašią į dobilo lapą. tRNR molekulė turi dvi aktyvias vietas: viename gale yra tripletas antikodonas, o kitame – akceptorinis galas (20 pav.).
Kiekviena aminorūgštis atitinka trijų nukleotidų derinį – tripletą. Amino rūgštis koduojantys tripletai – DNR kodonai – perduodami informacijos forma iš iRNR tripletų (kodonų). Dobilo lapo viršuje yra nukleotidų tripletas, kuris yra komplementarus atitinkamam mRNR kodonui. Šis tripletas skiriasi tRNR, turinčioms skirtingas aminorūgštis, ir koduoja būtent tą aminorūgštį, kurią neša ši tRNR. Jis vadinamas antikodonu.
Akceptoriaus galas yra aminorūgšties „nusileidimo vieta“.
Informacija arba matrica, RNR (mRNR) sudaro apie 5% visos ląstelių RNR. Οʜᴎ yra sintetinami vienos iš DNR molekulės grandinių dalyje ir perduoda informaciją apie baltymo struktūrą iš ląstelės branduolio į ribosomas, kur ši informacija yra įgyvendinama. Atsižvelgiant į priklausomybę nuo kopijų apimties informacija mRNR molekulė gali būti skirtingo ilgio.
Tačiau skirtingi RNR tipai yra viena funkcinė sistema, skirta paveldimos informacijos įgyvendinimui baltymų sintezės būdu.
RNR molekulės randamos ląstelės branduolyje, citoplazmoje, ribosomose, mitochondrijose ir plastiduose.
Visų tipų RNR, išskyrus virusų genetinę RNR, nėra pajėgios savaime daugintis ir susikaupti. Nukleino rūgštis. Nukleotidas. Dezoksiribonukleorūgštis arba DNR. Ribonukleino rūgštis arba RNR. Azoto bazės: adeninas, guaninas, citozinas, timinas, uracilas. Komplementarumas. Perneškite RNR (tRNR). Ribosominė RNR (rRNR). Messenger RNR (mRNR). 1. Kokia yra nukleotido sandara? 2. Kokia yra DNR molekulės sandara? 3. Koks yra papildomumo principas? 4. Kokie yra molekulių sandaros panašumai ir skirtumai 5. DNR ir RNR? 6. Kokius žinote RNR molekulių tipus? Kokia jų funkcija? 7. Vienos DNR grandinės fragmentas turi tokią sudėtį: A-A-G-G-C-C-C-T-T-. Naudodami papildomumo principą užbaikite antrąją grandinę.
DNR molekulėje timinai sudaro 24% visų azoto bazių. Nustatykite kitų azoto bazių skaičių šioje molekulėje.
Nobelio premija 1962 m. buvo įteikta dviem mokslininkams – J. Watsonui ir F. Crickui, kurie 1953 m. ᴦ. pasiūlė DNR molekulės struktūros modelį. Tai buvo patvirtinta eksperimentiškai. Šis atradimas turėjo didelę reikšmę genetikos, molekulinės biologijos ir kitų mokslų raidai. Virusai, skirtingai nei kiti organizmai, turi viengrandę DNR ir dvigrandę RNR.
RNR tipai. - koncepcija ir rūšys. Kategorijos „RNR tipai“ klasifikacija ir ypatybės. 2017 m., 2018 m.