Typer av RNA i cellen og deres struktur. Hva er DNA og RNA? DNA-struktur
Struktur
Nitrogenholdige baser i RNA kan danne hydrogenbindinger mellom cytosin og guanin, adenin og uracil, og mellom guanin og uracil. Imidlertid er andre interaksjoner også mulige, for eksempel kan flere adeniner danne en løkke, eller en løkke bestående av fire nukleotider, der det er et adenin-guanin-basepar. Et viktig strukturelt trekk ved RNA som skiller det fra DNA tilstedeværelse av en hydroksylgruppe ved 2"-posisjonen til ribose, noe som gjør at RNA-molekylet kan eksistere i A snarere enn B-konformasjonen som oftest observeres i DNA. A-formen har et dypt og smalt hovedspor og en grunn og bred moll rille Den andre konsekvensen av tilstedeværelsen av en 2" hydroksylgruppe er at den er konformasjonsplastisk. , det vil si at deler av RNA-molekylet som ikke deltar i dannelsen av dobbeltspiralen kan kjemisk angripe andre fosfatbindinger og bryte. dem ned Den "fungerende" formen av et enkeltstrenget RNA-molekyl, som proteiner, har ofte en tertiær struktur. Den tertiære strukturen er dannet på grunnlag av elementer i den sekundære strukturen dannet av hydrogenbindinger i et enkelt molekyl. Det finnes flere typer sekundære strukturelementer - stammeløkker, løkker og pseudoknoter. På grunn av det store antallet mulige baseparinger er å forutsi den sekundære strukturen til RNA en mye vanskeligere oppgave enn å forutsi den sekundære strukturen til proteiner, men det finnes nå effektive programmer som mfold.
Et eksempel på avhengigheten av funksjonen til RNA-molekyler på deres sekundære struktur er de interne ribosomentry-stedene (IRES). IRES er en struktur i 5"-enden av messenger-RNA som sikrer festing av et ribosom, og omgår den vanlige mekanismen for å initiere proteinsyntese, som krever tilstedeværelse av en spesiell modifisert base (cap) ved 5"-enden og proteininitieringsfaktorer . IRES ble opprinnelig oppdaget i virale RNA-er, men det er økende bevis for at cellulære mRNA-er også bruker en IRES-avhengig initieringsmekanisme under stressforhold.
Mange typer RNA, som rRNA og snRNA, fungerer i celler i form av komplekser med proteiner som assosieres med RNA-molekyler etter deres syntese eller (i eukaryoter) eksporterer fra kjernen til cytoplasma. Slike RNA-proteinkomplekser kalles ribonukleoproteinkomplekser eller ribonukleoproteiner.
Sammenligning med DNA
Det er tre hovedforskjeller mellom DNA og RNA:
DNA inneholder sukkeret deoksyribose, RNA inneholder ribose, som har en ekstra hydroksylgruppe sammenlignet med deoksyribose. Denne gruppen øker sannsynligheten for hydrolyse av molekylet, det vil si at den reduserer stabiliteten til RNA-molekylet.
Nukleotidet som er komplementært til adenin i RNA er ikke tymin, som i DNA, men uracil er den umetylerte formen av tymin.
DNA eksisterer i form av en dobbel helix, bestående av to separate molekyler. RNA-molekyler er i gjennomsnitt mye kortere og overveiende enkelttrådede.
Strukturanalyse av biologisk aktive RNA-molekyler, inkludert tRNA, rRNA, snRNA og andre molekyler som ikke koder for proteiner, viste at de ikke består av én lang helix, men av tallrike korte helixer som er plassert nær hverandre og danner noe som ligner på tertiær struktur av proteinet. Som et resultat kan RNA katalysere kjemiske reaksjoner, for eksempel består peptidyltransferasesenteret i ribosomet, involvert i dannelsen av peptidbindinger i proteiner, utelukkende av RNA.
Messenger (informasjon) RNA - RNA, som fungerer som et mellomledd i overføring av informasjon kodet i DNA til ribosomer, molekylære maskiner som syntetiserer proteiner i en levende organisme. Den kodende sekvensen til mRNA bestemmer aminosyresekvensen til polypeptidkjeden til et protein. Imidlertid koder de aller fleste RNA ikke for protein. Disse ikke-kodende RNA-ene kan transkriberes fra individuelle gener (f.eks. ribosomale RNA-er) eller avledes fra introner. De klassiske, godt studerte typene ikke-kodende RNA er overførings-RNA ( tRNA) og rRNA, som er involvert i translasjonsprosessen. Det er også klasser av RNA som er ansvarlige for genregulering, mRNA-behandling og andre roller. I tillegg er det ikke-kodende RNA-molekyler som kan katalysere kjemiske reaksjoner, slik som kutting og ligering av RNA-molekyler. I analogi med proteiner som er i stand til å katalysere kjemiske reaksjoner - enzymer (enzymer), kalles katalytiske RNA-molekyler ribozymer.
Deltar i sendingen
Informasjon om aminosyresekvensen til et protein finnes i mRNA. Tre påfølgende nukleotider (kodon) tilsvarer én aminosyre. I eukaryote celler behandles transcyribert forløper-mRNA eller pre-mRNA for å danne modent mRNA. Prosessering innebærer fjerning av ikke-proteinkodende sekvenser (introner). Etter dette eksporteres mRNA fra kjernen til cytoplasmaet, hvor det festes til ribosomer, som oversetter mRNA ved hjelp av tRNA koblet til aminosyrer.
I anukleerte celler (bakterier og archaea) kan ribosomer feste seg til mRNA umiddelbart etter transkripsjon av en del av RNA. Hos både eukaryoter og prokaryoter ender livssyklusen til mRNA med kontrollert ødeleggelse av enzymer ribonukleaser.
Transport (tRNA) er små molekyler, bestående av omtrent 80 nukleotider, med en konservativ tertiær struktur. De overfører spesifikke aminosyrer til stedet for peptidbindingssyntese i ribosomet. Hvert tRNA inneholder et sted for aminosyrebinding og et antikodon for gjenkjennelse og binding til mRNA-kodoner. Antikodonet danner hydrogenbindinger med kodonet, som plasserer tRNA i en posisjon som letter dannelsen av en peptidbinding mellom den siste aminosyren i peptidet som dannes og aminosyren festet til tRNA.
Ribosomalt RNA (rRNA) er den katalytiske komponenten i ribosomer. Eukaryote ribosomer inneholder fire typer rRNA-molekyler: 18S, 5.8S, 28S og 5S. Tre av de fire typene rRNA syntetiseres i kjernen. I cytoplasmaet kombineres ribosomale RNA-er med ribosomale proteiner for å danne et nukleoprotein kalt et ribosom. Ribosomet fester seg til mRNA og syntetiserer proteinet. rRNA utgjør opptil 80 % av RNA som finnes i cytoplasmaet til en eukaryot celle.
En uvanlig type RNA som fungerer som tRNA og mRNA (tmRNA) finnes i mange bakterier og plastider. Når ribosomet stopper ved defekte mRNAer uten stoppkodoner, fester tmRNA et lite peptid som leder proteinet til nedbrytning.
FUNKSJONER TIL RNA
Det finnes forskjellige typer RNA. Messenger (informasjon) RNA - RNA, som fungerer som et mellomledd i overføring av informasjon kodet i DNA til ribosomer, molekylære maskiner som syntetiserer proteiner i en levende organisme. Overfør RNA (tRNA) og rRNA som er involvert i translasjonsprosessen. Det er også klasser av RNA som er ansvarlige for genregulering, mRNA-behandling og andre roller. I tillegg er det ikke-kodende RNA-molekyler som kan katalysere kjemiske reaksjoner, slik som kutting og ligering av RNA-molekyler.
For å oppsummere alt ovenfor, kan det bemerkes at funksjonene til RNA:
- lagring av arvelig informasjon (for noen virus);
- deltakelse i prosessen med translasjon og transkripsjon (proteinsyntese);
- regulering av genaktivitet;
- katalysere en rekke kjemiske reaksjoner
det er tre typer RNA i en celle
Overfør RNA (tRNA). Overførings-RNA finnes hovedsakelig i cytoplasmaet til cellen. Funksjonen er å overføre aminosyrer til ribosomer, til stedet for proteinsyntese. Av det totale RNA-innholdet i en celle utgjør t-RNA ca. 10 %.
Ribosomalt RNA (r-RNA). Ribosomalt RNA utgjør en vesentlig del av strukturen til ribosomet. Av det totale RNA-innholdet i en celle utgjør r-RNA ca. 90 %.
Messenger RNA (i-RNA), eller messenger RNA (m-RNA). Inneholdt i kjernen og cytoplasma. Dens funksjon er å overføre informasjon om strukturen til proteinet fra DNA til stedet for proteinsyntese i ribosomer. mRNA står for omtrent 0,5-1 % av det totale RNA-innholdet i cellen.
Tilstede i RNA-molekylet i stedet for tymin. RNA-nukleotider inneholder ribose i stedet for deoksyribose. I en RNA-kjede er nukleotider forbundet med kovalente bindinger mellom ribosen til ett nukleotid og fosforsyreresten til et annet.
I kroppen finnes RNA i form av komplekser med proteiner - ribonukleoproteiner.
Det er 2 typer RNA-molekyler kjent:
1) Dobbelttrådet RNA er karakteristisk for noen virus - de tjener til å lagre og reprodusere arvelig informasjon (utføre funksjonene til kromosomer).
2) I de fleste celler utfører enkeltstrengede RNA-er overføring av informasjon om aminosyresekvensen i proteiner fra kromosomet til ribosomet.
Enkeltrådet RNA har romlig organisering: på grunn av interaksjonen av nitrogenholdige baser med hverandre, så vel som med fosfater og hydroksyler i sukker-fosfat-ryggraden, foldes kjeden til en kompakt kulestruktur. Funksjon: overføre informasjon fra kromosom til ribosomer om AK-sekvensen i proteiner som må syntetiseres.
Det finnes flere typer enkelttrådet RNA basert på deres funksjon eller plassering i cellen:
1. Ribosomalt RNA (rRNA) utgjør hoveddelen av RNA i cytoplasmaet (80-90%). Størrelser 3000-5000 basepar. Den sekundære strukturen er i form av dobbeltspiralformede hårnåler. r-RNA er en strukturell komponent av ribosomer - celleorganeller hvor proteinsyntese finner sted. Ribosomer er lokalisert i cytoplasma, nukleolus, mitokondrier og kloroplaster. De består av to underenheter - store og små. Den lille underenheten består av ett rRNA-molekyl og 33 proteinmolekyler, den store underenheten - 3 rRNA-molekyler og 50 proteiner. Ribosomale proteiner utfører enzymatiske og strukturelle funksjoner.
Funksjoner av rRNA:
1) strukturell komponent ribosomer- deres integritet er nødvendig for proteinbiosyntese;
2) sikre korrekt binding av ribosomet til m-RNA;
3) sikre korrekt binding av ribosomet til t-RNA;
2. Matrise (mRNA) - 2-6 % av den totale mengden RNA.
Består av seksjoner:
1) cistroner - bestemme sekvensen av aminosyrer i proteinene de koder for og har en unik nukleotidsekvens;
2) utranslaterte regioner er lokalisert i endene av molekylet og har generelle mønstre av nukleotidsammensetning.
Cap - en spesiell struktur i 5′-enden av m-RNA - er 7-metylguanosintrifosfat, dannet enzymatisk under transkripsjon.
Cap funksjoner:
1) beskytter 5′-enden mot spaltning av eksonukleaser,
2) brukes for spesifikk gjenkjennelse av m-RNA under translasjon.
Precistronic uoversatt region - 3-15 nukleotider. Funksjon: å sikre korrekt interaksjon av 5'-enden av m-RNA med ribosomet.
Cistron: inneholder initierings- og termineringskodoner - spesielle nukleotidsekvenser som er ansvarlige for begynnelsen og slutten av informasjonsoverføring fra en gitt cistron.
Postcistronisk utranslatert region - lokalisert i 3′-enden, inneholder et heksanukleotid (ofte AAUAAA) og en kjede på 20-250 adenylnukleotider. Funksjon - opprettholde intracellulær stabilitet av m-RNA.
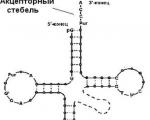
3. Overfør RNA (tRNA) - 15 % av totalt RNA, består av 70-93 nukleotidpar. Funksjon: overføring av en aminosyre til stedet for proteinsyntese de "gjenkjenner" (ved prinsippet om komplementaritet) regionen til mRNA som tilsvarer den overførte aminosyren. For hver av de 20 AA-ene er det spesifikke tRNA-er (vanligvis mer enn én). Alle tRNA-er har en kompleks struktur, avbildet i form av et kløverblad.

Kløverbladet inneholder 5 seksjoner:
1) 3′-ende - akseptorgren (AK-resten er festet her med en eterbinding),
2) antikidongren - plassert motsatt akseptorstedet, består av tre uparede (har frie bindinger) nukleotider (antikodon) og spesifikt par (antiparallell, komplementær) med m-RNA-kodonet.
Kodon- et sett med 3 nukleotider (triplett) i m-RNA, som bestemmer plasseringen av denne aminosyren i den syntetiserte polypeptidkjeden. Dette er en enhet av genetisk kode ved hjelp av hvilken all genetisk informasjon "registreres" i DNA- og RNA-molekyler.
3) T-gren (pseudouredin-løkke - inneholder pseudouredin) - en seksjon som festes til ribosomet.
4) D-gren (dehydrouredinløkke - inneholder dehydrouredin) - en seksjon som sikrer interaksjon med enzymet aminoacyl-tRNA-syntetase tilsvarende aminosyren.
5) Ekstra liten gren. Funksjonene er ennå ikke studert.
6) Kjernefysisk RNA (nRNA) - en komponent i cellekjernen. Lav polymer, stabil, rollen som fortsatt er uklar.
Alle typer RNA syntetiseres i cellekjernen på en DNA-matrise under påvirkning av enzymer polymeraser. I dette tilfellet dannes en sekvens av ribonukleotider som er komplementær til sekvensen av deoksyribonukleotider i DNA - dette er prosessen med transkripsjon.
Alle typer RNA er designet for å trekke ut informasjon om strukturen til et protein fra DNA og sikre proteinbiosyntese i samsvar med denne informasjonen. RNA er en enkelt polynukleotidkjede konstruert av fire hovedtyper av ribonukleotider - AMP, GMP, CMP og UMP. RNA er preget av mindre nukleotider med uvanlige nitrogenholdige baser - dihydrouracil, 3-metyluracil, 1-metylguanin, etc. (opptil 50 typer). Det er spesielt mange av dem i aminoacyl-tRNA (opptil 10 % av alle nukleotider). I RNA samsvarer ikke innholdet av adenin og guanin med innholdet av uracil og cytosin.
Følgende typer RNA skilles:
Messenger eller messenger RNA(m- eller i-RNA), m.m. 25000-1000000 Ja, består av 75-300 nukleotider, syntetisert i kjernen fra pre-m-RNA; utgjør 5-7 % av alt cellulært RNA. Halveringstid er flere minutter. I 5'-enden av alle eukaryote mRNA er det en spesiell struktur kalt lokk. Cap er 7-metylguanosintrifosfat. Cap-dannelse skjer enzymatisk i kjernen før transkripsjonen er fullført. Det antas at hetten på den ene siden beskytter 5'-enden av mRNA fra spaltningen av 5'-eksonukleaser, og på den annen side brukes til spesifikk gjenkjennelse i translasjonssystemet. Hetten etterfølges av delen som ikke er kringkastet, hvor det (fra 3-15 nukleotider før startkodonet) er en nukleotidsekvens komplementær til rRNA-sekvensen. Dens rolle er å sikre korrekt interaksjon av 5'-enden med ribosomet. Kringkastingsdelen avsluttes stopp kodon, ofte etterfulgt av heksanukleotidet AAAAAAA. I de fleste mRNA-er inneholder 3'-enden en polyadenylatkjede på 20-250 adenylnukleotider, som ikke er et resultat av transkripsjon, men er festet til mRNA enzymatisk under modning i kjernen. Det antas at polyadenylatsekvensen er ansvarlig for å opprettholde den intracellulære stabiliteten til mRNA og bestemmer levetiden. Kodeelementet er en triplett av nukleotider (kodon) som koder for en aminosyre. Det er vist at flere dobbelttrådete hårnåler dannes i et lineært mRNA-molekyl, i endene som det er "tegn" på transkripsjonsinitiering og -avslutning. I den sekundære strukturen er det en buet kjede; I følge noen data er polynukleotidkjeden i den tertiære strukturen forbundet (sår) med transportproteinet informofer.
Overfør RNA ( tRNA ) – ca 15 %. Overførings-RNA-er har en liten molekylvekt (~ 25 000) og er inneholdt i den løselige fraksjonen av cytoplasmaet, og utfører funksjonen med å overføre aminosyrer til stedet for proteinsyntese - ribosomet. Inneholder ca. 75 nukleotider. Cellen inneholder minst 20 typer tRNA-molekyler. Hver eller flere typer tRNA tilsvarer en av de 20 aminosyrene som er nødvendige for proteinbiosyntese. Den sekundære strukturen til alle tRNA-er ligner " kløverblad» (Fig. 12.3) og har 4 hovedseksjoner. 1) Akseptør stedet har nukleotidsekvensen CCA i 3"-enden. Karboksylgruppen til aminosyren blir tilsatt 3"-hydroksylgruppen til adenosylresten. Overførings-tRNA-er knyttet til aminosyrer kalles aminoacyl-tRNA(aatRNA). De opptrer adapter funksjon når du oversetter trebokstavskoden til nukleinsyrer til 20-bokstavssekvensen av aminosyrer i polypeptidkjeden. 2) Antikodonløkke nødvendig for å feste seg til en triplett eller kodon av mRNA. 3) Pseudouridyl-løkke(TψC) består av 7 nukleotider og inneholder en pseudouridinrest; tjener til å binde tRNA til ribosomet; 4) Dihydrouridinløkke(D) består av 8-12 nukleotidrester. Nødvendig for binding til aminoacyl-tRNA-syntetase, som er involvert i gjenkjennelsen av dets tRNA av en aminosyre. Den tertiære strukturen er representert av en romlig struktur i form av en albuebøyning (L-form).
Ris. 12.3. Strukturen til tRNA
Ribosomalt RNA (rRNA)– 80-85 %, har forskjellige og betydelig høyere molekylvekter (35000-1000000, som tilsvarer 100-3100 nukleotider), er strukturelle komponenter i ribosomer.
Ribosomer gir spesifikk kontakt mellom mRNA og tRNA, som et resultat av at nukleotidsekvensen avlest fra et spesifikt gen blir oversatt til aminosyresekvensen til det tilsvarende proteinet. Pattedyrribosomer består av to nukleoproteinunderenheter - en stor med en sedimentasjonskonstant på 60S og en liten - 40S (i prokaryoter - henholdsvis 50S og 30S). 60S-underenheten inneholder 5S ribosomalt RNA (rRNA), 5.8S rRNA og 28S rRNA. Den lille 40S-enheten inkluderer et enkelt 18S rRNA og omtrent 30 polypeptidkjeder. Alle ribosomale RNA, med unntak av 5S RNA, har en felles forløper - 45S RNA, lokalisert i kjernen. I nukleolen er høyt metylerte ribosomale RNA-er pakket med ribosomale proteiner.
Prokaryote ribosomer og ribosomer av mitokondrier og plastider inneholder færre komponenter, men er strukturelt og funksjonelt svært like eukaryote. Den sekundære strukturen til rRNA dannes på grunn av korte dobbelttrådede deler av molekylet - hårnåler. Omtrent 2/3 av rRNA er organisert i hårnåler, 1/3 er representert av enkelttrådede regioner rike på purin-nukleotider, som proteiner hovedsakelig binder til. Ribosomale proteiner, som histoner, har en grunnleggende karakter og utfører både strukturelle og enzymatiske roller.
Rollen til en mellommann, hvis funksjon er å oversette den arvelige informasjonen som er lagret i DNA til en fungerende form, spilles av ribonukleinsyrer - RNA.
To- og en-trådet RNA-molekyler er kjent. Dobbelttrådet RNA tjener til å lagre og reprodusere arvelig informasjon i noen virus, dvs. de utfører funksjonene til kromosomer. Enkeltrådede RNA-er bærer informasjon om sekvensen av aminosyrer i proteiner fra kromosomet til stedet for deres syntese og deltar i synteseprosesser.
I motsetning til DNA-molekyler er ribonukleinsyrer representert av en enkelt polynukleotidkjede, som består av fire typer nukleotider som inneholder sukker, ribose, fosfat og en av fire nitrogenholdige baser - adenin, guanin, uracil eller cytosin. RNA syntetiseres på DNA-molekyler ved bruk av RNA-polymerase-enzymer i samsvar med prinsippet om komplementaritet og antiparallelisme, og uracil er komplementær til DNA-adenin i RNA. Hele variasjonen av RNA som opererer i cellen kan deles inn i tre hovedtyper: mRNA, tRNA, rRNA.
Matrise, eller informasjon, RNA (mRNA eller mRNA). Transkripsjon. For å syntetisere proteiner med spesifiserte egenskaper, sendes "instruksjoner" til stedet for deres konstruksjon om rekkefølgen for inkludering av aminosyrer i peptidkjeden. Denne instruksjonen er inneholdt i nukleotidsekvensen matrise, eller messenger-RNA(mRNA, mRNA) syntetisert i de tilsvarende delene av DNA. Prosessen med mRNA-syntese kalles transkripsjon.
Syntesen av mRNA begynner med oppdagelsen av RNA-polymerase av en spesiell region i DNA-molekylet, som indikerer stedet der transkripsjonen begynner - promotør Etter binding til promoteren, vikler RNA-polymerase av den tilstøtende svingen av DNA-helixen. To DNA-tråder divergerer på dette punktet, og på en av dem syntetiserer enzymet mRNA. Sammenstillingen av ribonukleotider til en kjede skjer i samsvar med deres komplementaritet til DNA-nukleotider, og også antiparallell med hensyn til DNA-templatetråden. På grunn av det faktum at RNA-polymerase er i stand til å sette sammen et polynukleotid bare fra 5"-enden til 3"-enden, kan bare én av de to DNA-trådene, nemlig den som vender mot enzymet med sin 3-tommers ende, tjene som mal. for transkripsjon ( 3" → 5"). Denne kjeden kalles kodogen
tRNA- RNA, hvis funksjon er å transportere aminosyrer til stedet for proteinsyntese. tRNA-er tar også en direkte del i forlengelsen av polypeptidkjeden ved å slutte seg til - å være i kompleks med en aminosyre - til mRNA-kodonet og gi den komplekse konformasjonen som er nødvendig for dannelsen av en ny peptidbinding.
Hver aminosyre har sitt eget tRNA tRNA er enkelttrådet RNA, men i sin funksjonelle form har det en "kløverblad" eller "kløverblad". Aminosyren er kovalent festet til 3"-enden av molekylet ved hjelp av enzymet aminoacyl-tRNA-syntetase, spesifikt for hver type tRNA. På sted C er det et antikodon som tilsvarer aminosyren.
(rRNA)- flere RNA-molekyler som danner grunnlaget for ribosomet. Hovedfunksjonen til rRNA er å utføre translasjonsprosessen - lese informasjon fra mRNA ved hjelp av tRNA-adaptermolekyler og katalysere dannelsen av peptidbindinger mellom aminosyrer festet til tRNA.
| Parameternavn | Betydning |
| Artikkel emne: | Typer RNA. |
| Rubrikk (tematisk kategori) | Sport |
Det er tre grunnleggende typer RNA, forskjellig i struktur, størrelse på molekyler, plassering i cellen og utførte funksjoner.
Ribosomale RNA (rRNA) syntetiseres hovedsakelig i nukleolus og utgjør omtrent 85 % av alt RNA i cellen. Οʜᴎ er en del av ribosomer og deltar i dannelsen av det aktive sentrum av ribosomet, hvor prosessen med proteinbiosyntese skjer.
Overfør RNA (tRNA) dannes i kjernen på DNA, for så å bevege seg inn i cytoplasmaet. Οʜᴎ utgjør omtrent 10 % av cellulært RNA og er de minste RNA-ene, bestående av 70-100 nukleotider. Hvert tRNA fester en spesifikk aminosyre og transporterer den til polypeptidsamlingsstedet i ribosomet. Alle kjente tRNA-er, på grunn av komplementære interaksjoner, danner en sekundær struktur formet som et kløverblad. Et tRNA-molekyl har to aktive steder: et triplettantikodon i den ene enden og en akseptorende i den andre (fig. 20).
Hver aminosyre tilsvarer en kombinasjon av tre nukleotider - en triplett. Aminosyrekodende tripletter – DNA-kodoner – overføres i form av informasjon fra mRNA-tripletter (kodoner). På toppen av kløverbladet er det en triplett av nukleotider som er komplementær til det tilsvarende mRNA-kodonet. Denne tripletten er forskjellig for tRNA som bærer forskjellige aminosyrer, og koder for nøyaktig aminosyren som bæres av dette tRNA. Det kalles antikodon.
Akseptorenden er "landingsstedet" for aminosyren.
Informasjon, eller matrise, RNA (mRNA) utgjør omtrent 5 % av alt cellulært RNA. Οʜᴎ syntetiseres på en del av en av kjedene til DNA-molekylet og overfører informasjon om strukturen til proteinet fra cellekjernen til ribosomene, hvor denne informasjonen implementeres. Tar hensyn til avhengigheten av volumet av kopiert informasjon et mRNA-molekyl kan ha forskjellige lengder.
Imidlertid representerer forskjellige typer RNA et enkelt funksjonelt system som tar sikte på å implementere arvelig informasjon gjennom proteinsyntese.
RNA-molekyler finnes i cellens kjerne, cytoplasma, ribosomer, mitokondrier og plastider.
Alle typer RNA, med unntak av det genetiske RNA fra virus, er ikke i stand til selvduplisering og selvmontering. Nukleinsyre. Nukleotid. Deoksyribonukleinsyre eller DNA. Ribonukleinsyre eller RNA. Nitrogenbaser: adenin, guanin, cytosin, tymin, uracil. Komplementaritet. Overfør RNA (tRNA). Ribosomalt RNA (rRNA). Messenger RNA (mRNA). 1. Hva er strukturen til et nukleotid? 2. Hva er strukturen til DNA-molekylet? 3. Hva er komplementaritetsprinsippet? 4. Hva er likhetene og hvilke forskjeller er det i strukturen til molekyler 5. DNA og RNA? 6. Hvilke typer RNA-molekyler kjenner du til? Hva er deres funksjon? 7. Et fragment av en DNA-streng har følgende sammensetning: A-A-G-G-C-C-C-T-T-. Bruk prinsippet om komplementaritet, fullfør den andre kjeden.
I DNA-molekylet står tyminer for 24 % av det totale antallet nitrogenholdige baser. Bestem antall andre nitrogenholdige baser i dette molekylet.
Nobelprisen 1962 ᴦ. ble tildelt to forskere - J. Watson og F. Crick, som i 1953 ᴦ. foreslått en modell av strukturen til DNA-molekylet. Det ble bekreftet eksperimentelt. Denne oppdagelsen var av stor betydning for utviklingen av genetikk, molekylærbiologi og andre vitenskaper. Virus inneholder, i motsetning til andre organismer, enkelttrådet DNA og dobbelttrådet RNA.
Typer RNA. - konsept og typer. Klassifisering og funksjoner i kategorien "RNA-typer." 2017, 2018.