Ved dechiffrering av insektgenomet ble det funnet at. Medisinsk genetikk
Hoppende gener
I midten av forrige århundre oppdaget den amerikanske forskeren Barbara McClintock fantastiske gener i mais som uavhengig kan endre deres posisjon på kromosomene. Nå kalles de "hoppende gener" eller transponerbare (mobile) elementer. Oppdagelsen ble ikke gjenkjent på lenge, og vurderte mobile elementer for å være et unikt fenomen som bare er karakteristisk for mais. Det var imidlertid for denne oppdagelsen at McClintock ble tildelt Nobelprisen i 1983 – i dag er det funnet hoppende gener i nesten alle studerte arter av dyr og planter.
Hvor kom hoppende gener fra, hva gjør de i en celle, er de nyttige? Hvorfor, med genetisk sunne foreldre, kan en Drosophila fruktfluefamilie, på grunn av hoppgener, produsere mutant avkom med høy frekvens eller til og med være barnløs? Hvilken rolle spiller hoppende gener i evolusjonen?
Det må sies at genene som sikrer cellefunksjonen er plassert på kromosomene i en viss rekkefølge. Takket være dette var det mulig å konstruere såkalte genetiske kart for mange arter av encellede og flercellede organismer. Imidlertid er det en størrelsesorden mer genetisk materiale mellom gener enn innenfor dem! Hvilken rolle denne "ballast"-delen av DNA spiller er ikke fullt ut fastslått, men det er her man oftest finner mobile elementer, som ikke bare beveger seg selv, men også kan ta med seg nabo-DNA-fragmenter.

Hvor kommer hoppgener fra? Det antas at i det minste noen av dem stammer fra virus, siden noen mobile elementer er i stand til å danne viruspartikler (for eksempel det mobile elementet sigøyner i fruktflua Drosophila melanogaster). Noen mobile elementer vises i genomet gjennom den såkalte horisontal overføring fra andre arter. For eksempel er det etablert at mobil uteligger-element (oversatt til russisk kalles det en tramp) Drosophila melanogaster gjentatte ganger gjeninnført i genomet til denne arten. Det er en versjon om at enkelte regulatoriske deler av DNA også kan ha autonomi og en tendens til "vagrancy".
Nyttig ballast
På den annen side oppfører de fleste hoppegenene seg, til tross for navnet, stille, selv om de utgjør en femtedel av det totale genetiske materialet Drosophila melanogaster eller nesten halvparten av det menneskelige genomet.
Redundansen til DNA, som ble nevnt ovenfor, har sin fordel: ballast-DNA (inkludert passive mobile elementer) tar støtet hvis fremmed DNA blir introdusert i genomet. Sannsynligheten for at et nytt element blir integrert i et nyttig gen og dermed forstyrrer dets drift reduseres hvis det er mye mer ballast-DNA enn signifikant DNA.
Noe overflødighet av DNA er nyttig på samme måte som "redundans" av bokstaver i ord: vi skriver "Maria Ivanovna", men sier "Marivan". Noen bokstaver går uunngåelig tapt, men meningen består. Det samme prinsippet fungerer på nivået av betydning for individuelle aminosyrer i et protein-enzymmolekyl: bare sekvensen av aminosyrer som danner det aktive senteret er strengt bevart. Dermed viser redundans seg på ulike nivåer å være en slags buffer som gir en reserve av systemstyrke. Slik viser det seg at mobile elementer som har mistet mobilitet, ikke er ubrukelige for genomet. Som de sier, "fra en tynn sau i det minste en dusk ull," selv om kanskje et annet ordtak ville være bedre egnet her - "hver bast i en linje."

Mobile elementer som har beholdt evnen til å hoppe beveger seg langs Drosophila-kromosomer med en frekvens på 10–2–10–5 per gen per generasjon, avhengig av type grunnstoff, genetisk bakgrunn og ytre forhold. Det betyr at ett av hundre hoppende gener i en celle kan endre posisjon etter neste celledeling. Som et resultat, etter flere generasjoner, kan fordelingen av mobile elementer langs kromosomet endre seg veldig betydelig.
Det er praktisk å studere denne fordelingen på polyten (flertrådede) kromosomer fra spyttkjertlene til Drosophila-larvene. Disse kromosomene er mange ganger tykkere enn vanlig, noe som i stor grad forenkler undersøkelsen deres under et mikroskop. Hvordan oppnås slike kromosomer? I cellene i spyttkjertlene multipliseres DNAet til hvert kromosom, som ved normal celledeling, men selve cellen deler seg ikke. Som et resultat endres ikke antall celler i kjertelen, men over 10-11 sykluser samler det seg flere tusen identiske DNA-tråder i hvert kromosom.

Det er delvis på grunn av polytenkromosomer at hoppgener i Drosophila er bedre studert enn i andre flercellede organismer. Som et resultat av disse studiene viste det seg at selv innenfor den samme Drosophila-populasjonen er det vanskelig å finne to individer som har kromosomer med samme fordeling av transponerbare elementer. Det er ingen tilfeldighet at det antas at de fleste av de spontane mutasjonene i Drosophila er forårsaket av bevegelsen til disse "hopperne".
Konsekvensene kan variere...
Basert på deres effekt på genomet, kan aktive mobile elementer deles inn i flere grupper. Noen av dem utfører funksjoner som er ekstremt viktige og nyttige for genomet. For eksempel, telomerisk DNAet som ligger i endene av kromosomene i Drosophila består av spesielle mobile elementer. Dette DNA er ekstremt viktig - tapet medfører tap av hele kromosomet under celledeling, noe som fører til celledød.
Andre mobile elementer er direkte "skadedyr". Det er i hvert fall det de anses å være for øyeblikket. For eksempel kan mobile elementer av R2-klassen spesifikt inkorporeres i leddyrgener som koder for et av de ribosomale proteinene, de cellulære "fabrikkene" for proteinsyntese. Personer med slike lidelser overlever bare fordi bare en del av de mange genene som koder for disse proteinene er skadet i genomet.

Det er også mobile elementer som bare beveger seg i reproduktive vev som produserer kjønnsceller. Dette forklares med det faktum at i forskjellige vev kan det samme mobile elementet produsere forskjellige lengder og funksjoner av enzymproteinmolekylene som kreves for bevegelse.
Et eksempel på sistnevnte er P-elementet Drosophila melanogaster, som kom inn i sine naturlige populasjoner gjennom horisontal overføring fra en annen art av Drosophila for ikke mer enn hundre år siden. Imidlertid er det knapt en befolkning på jorden nå Drosophila melanogaster, der P-elementet ikke ville bli funnet. Det skal bemerkes at de fleste av kopiene er defekte, dessuten ble den samme versjonen av defekten funnet nesten overalt. Sistnevntes rolle i genomet er unik: den er "intolerant" overfor sine medmennesker og spiller rollen som en undertrykker, og blokkerer deres bevegelse. Så beskyttelsen av Drosophila-genomet fra hoppene til den "fremmede" kan delvis utføres av dets egne derivater.
Det viktigste er å velge de riktige foreldrene!
De fleste hoppene til mobile elementer påvirker ikke utseendet til Drosophila, fordi de forekommer på ballast-DNA, men det er andre situasjoner når aktiviteten deres øker kraftig.

Overraskende nok er den kraftigste faktoren som induserer bevegelsen av hoppende gener dårlig foreldrevalg. Hva skjer for eksempel hvis du krysser kvinner fra en laboratoriepopulasjon? Drosophila melanogaster, som ikke har P-elementet (fordi deres forfedre ble fanget fra naturen for rundt hundre år siden), med hanner som bærer P-elementet? I hybrider, på grunn av den raske bevegelsen av det mobile elementet, kan det oppstå et stort antall forskjellige genetiske lidelser. Dette fenomenet, kalt hybrid dysgenese, er forårsaket av det faktum at det ikke er noen repressor i mors cytoplasma som forbyr bevegelsen av det transponerbare elementet.
Så hvis brudgom fra populasjon A og bruder fra populasjon B kan skape store familier, er det ikke alltid det motsatte er sant. En familie med genetisk sunne foreldre kan produsere et stort antall mutante eller infertile avkom, eller til og med være barnløse hvis far og mor har et annet sett med mobile elementer i genomet. Spesielt mange brudd oppstår hvis eksperimentet utføres ved en temperatur på 29 ° C. Påvirkningen av eksterne faktorer, lagt på den genetiske bakgrunnen, forsterker effekten av genommismatch, selv om disse faktorene i seg selv (selv ioniserende stråling) alene ikke er i stand til å forårsake slike massive bevegelser av mobile elementer.
Lignende hendelser i Drosophila melanogaster kan forekomme med deltakelse av andre familier av mobile elementer.
"Mobil" evolusjon
Det cellulære genomet kan betraktes som et slags økosystem av permanente og midlertidige medlemmer, der naboer ikke bare sameksisterer, men også samhandler med hverandre. Samspillet mellom vertsgener og mobile elementer er fortsatt dårlig forstått, men mange resultater kan gis - fra organismens død i tilfelle skade på et viktig gen til gjenoppretting av tidligere skadede funksjoner.
Det hender at hoppegenene selv samhandler med hverandre. Således er et fenomen som ligner immunitet kjent, når et mobilt element ikke kan trenge inn i umiddelbar nærhet til et allerede eksisterende. Imidlertid er ikke alle mobile elementer så delikate: For eksempel kan P-elementer lett trenge inn i hverandre og ta medspillerne ut av spillet.

I tillegg er det en slags selvregulering i antall mobile elementer i genomet. Faktum er at mobile elementer kan utveksle homologe regioner med hverandre - denne prosessen kalles rekombinasjon. Som et resultat av slik interaksjon kan mobile elementer, avhengig av deres orientering, miste ( sletting) eller utvide ( inversjon) fragmenter av verts-DNA lokalisert mellom dem. Hvis en betydelig del av et kromosom går tapt, vil genomet dø. Ved en inversjon eller liten delesjon skapes kromosomdiversitet, som anses som en nødvendig betingelse for evolusjon.
Hvis det oppstår rekombinasjoner mellom mobile elementer lokalisert på forskjellige kromosomer, er resultatet dannelsen av kromosomale omorganiseringer, som under påfølgende celledelinger kan føre til genomubalanse. Og et ubalansert genom, som et ubalansert budsjett, er delt veldig dårlig. Så døden til mislykkede genomer er en av grunnene til at aktive mobile elementer ikke fyller kromosomer på ubestemt tid.
Et naturlig spørsmål dukker opp: hvor betydelig er bidraget fra mobile elementer til evolusjonen? For det første introduseres de fleste mobile elementene, grovt sett, uansett hvor de skal være, som et resultat av at de kan skade eller endre strukturen eller reguleringen av genet de introduseres i. Da avviser naturlig utvalg mislykkede alternativer, og vellykkede alternativer med adaptive egenskaper fikses.

Hvis konsekvensene av introduksjonen av et mobilt element viser seg å være nøytrale, kan denne varianten vedvare i befolkningen, og gi noe mangfold i genstrukturen. Dette kan komme godt med under ugunstige forhold. Teoretisk sett, med den massive bevegelsen av mobile elementer, kan mutasjoner vises i mange gener samtidig, noe som kan være svært nyttig i tilfelle en skarp endring i levekårene.
Så for å oppsummere: det er mange mobile elementer i genomet og de er forskjellige; de kan samhandle både med hverandre og med vertsgener; kan skade og være uerstattelig. Genom-ustabilitet forårsaket av bevegelse av mobile elementer kan ende i tragedie for individet, men evnen til å endre seg raskt er en nødvendig betingelse for overlevelse av en populasjon eller art. Takket være dette skapes mangfold, som er grunnlaget for naturlig utvalg og påfølgende evolusjonære transformasjoner.
En analogi kan trekkes mellom hoppende gener og innvandrere: Noen innvandrere eller deres etterkommere blir likeverdige borgere, andre får oppholdstillatelse, og andre – de som ikke overholder lovene – blir deportert eller fengslet. Og massemigrasjoner av mennesker kan raskt endre selve staten.
Litteratur
Ratner V. A., Vasilyeva L. A. Induksjon av transposisjoner av mobile genetiske elementer ved stresspåvirkning. Russisk binding. 2000.
Gvozdev V. A. Mobile DNA of eukaryotes // Soros pedagogisk tidsskrift. 1998. Nr. 8.
) funnet i genomet til fruktfluen ( Drosophila ananassae) en fullstendig kopi av genomet til en parasittisk bakterie Wolbachia.
Wolbachia-bakterien lever i cytoplasmaet til vertsceller og er kjent for å ha lært å finregulere reproduksjonen, utviklingen og til og med utviklingen av vertene. Derfor kalles den ofte "mikrobemanipulatoren" eller "fluenes herre" (siden den lever i insektceller).
Forskningen begynte da Julie Dunning-Hotopp fra JCVI oppdaget hvordan noen Wolbachia-gener "samarbeider" med Drosophila-gener, som om de var en del av det samme genomet.
Michael Clark, en forsker ved University of Rochester, bosatte kolonien Drosophila ananassae i laboratoriet for å forstå, sammen med Warren, hva hemmeligheten er.
Wolbachia-genet i Drosophila-genomet (illustrasjon fra University of Rochester).
"I flere måneder trodde jeg at jeg tok feil," sier Clarke, "Jeg antok til og med at antibiotikaresistens hadde utviklet seg, fordi jeg fant hvert Wolbachia-gen om og om igjen. Da jeg endelig tok vevet som jeg hadde latt være alene for flere måneder siden, fant jeg ikke selve Wolbachia.»
Nå prøver Warren og Clark å forstå hva fordelen med å sette inn et så stort stykke DNA er for Drosophila - kanskje "fremmede" gener gir verten noen nye muligheter.

Og slik går Wolbachia-genene inn i vertens DNA (illustrasjon av Nicolle Rager Fuller, National Science).
Resultatene av studien ble publisert i en artikkel i tidsskriftet Science. I den antyder forfatterne at horisontal genoverføring (overføring av gener mellom ubeslektede arter) skjer mye hyppigere mellom bakterier og flercellede organismer i vår verden enn tidligere antatt.
Å dechiffrere de molekylærgenetiske manipulasjonsmekanismene utført av Wolbachia med vertene vil gi mennesker kraftige nye midler til å påvirke levende organismer og naturen som helhet.
Imidlertid er ikke alle insekter mottakelige for den dårlige påvirkningen fra Wolbachia. For eksempel "lærte" sommerfugler fra de samoanske øyene å beskytte hannene sine. Jeg lurer på om malariamyggen som de ønsker å infisere med denne bakterien vil lære å bekjempe den?
Eksempel på den allrussiske testen i biologi
11. klasse
Instruksjoner for utførelse av arbeidet
Testen inkluderer 14 oppgaver. Det settes av 1 time og 30 minutter (90 minutter) til å gjennomføre biologiarbeidet.
Svar på oppgaver er en tallsekvens, et tall, et ord (frase) eller et kort fritt svar, som skrives ned i feltet som er avsatt for dette arbeidet. Hvis du skriver ned et feil svar, stryk det ut og skriv et nytt ved siden av.
Når du gjennomfører oppgaver kan du bruke et utkast. Oppføringer i utkastet tas ikke hensyn til ved karaktersetting av arbeid. Vi anbefaler deg å fullføre oppgavene i den rekkefølgen de er gitt. For å spare tid kan du hoppe over en oppgave du ikke kan fullføre umiddelbart og gå videre til neste. Hvis du har tid igjen etter å ha fullført alt arbeidet, kan du gå tilbake til de tapte oppgavene.
Poengene du får for utførte oppgaver summeres.
Prøv å fullføre så mange oppgaver som mulig og få flest poeng.
Forklaringer for utvalget av det all-russiske testarbeidet
Når du skal gjøre deg kjent med et eksempelprøvearbeid, bør du huske på at oppgavene som er inkludert i prøven ikke gjenspeiler alle ferdighets- og innholdsproblemene som vil bli testet som en del av det all-russiske testarbeidet. En fullstendig liste over innholdselementer og ferdigheter som kan testes i arbeidet er gitt i kodifikatoren av innholdselementer og krav til nivået på opplæring av kandidater for utvikling av en CD i biologi. Formålet med prøvetestarbeidet er å gi en ide om strukturen til VPR, antall og form på oppgaver, og deres kompleksitetsnivå.
1. I forsøket belyste forsøkslederen en del av dråpen med amøbene i den. Etter kort tid begynte protozoene å bevege seg aktivt i én retning.
1.1. Hvilken egenskap ved organismer illustrerer eksperimentet?
Forklaring: det er 7 egenskaper ved levende organismer (det er ved disse egenskapene at levende ting skiller seg fra ikke-levende ting): ernæring, respirasjon, irritabilitet, mobilitet, utskillelse, reproduksjon, vekst. Amøber beveger seg fra den lyse delen av dråpen til den mørke delen, når de reagerer på lys, det vil si at vi velger egenskapen - irritabilitet.
Svar: irritabilitet.
1.2. Gi et eksempel på et lignende fenomen hos planter.
Forklaring: her kan vi skrive et hvilket som helst eksempel på en reaksjon (manifestation av irritabilitet) hos planter.
Svar: lukking av fangstapparatet i kjøttetende planter ELLER vending av blader mot solen eller bevegelse av solsikker i løpet av dagen etter solen ELLER bøyning av stilker på grunn av endringer i landskapet (miljøet).
2. I skogkanten lever og samhandler mange planter, dyr, sopp og mikroorganismer. Tenk på en gruppe som inkluderer huggorm, ørn, pinnsvin, viviparøse og gresshoppe. Fullfør oppgavene.
2.1. Merk objektene vist på fotografiene og tegningene som er inkludert i gruppen ovenfor.

1 - viviparøs øgle
2 - hoggorm
3 - pinnsvinlag
4 - vanlig gresshoppe
5 - ørn
2.2. Klassifiser disse organismene etter deres plassering i næringskjeden. I hver celle skriver du ned nummeret eller navnet på ett av objektene i gruppen.
Næringskjede: pinnsvin - vanlig gresshoppe - viviparøse - hoggorm - ørn.
Forklaring: vi starter næringskjeden med produsenten (grønn plante - produsent av organiske stoffer) - pinnsvinet, deretter 1. ordens forbruker (forbrukerne inntar organiske stoffer og har flere bestillinger) - den vanlige gresshoppen, viviparøse øgle (2. ordens forbruker) , hoggorm (3. ordens forbruker), ørn (4. ordens forbruker).
2.3. Hvordan vil reduksjonen i antall pinnsvin i laget påvirke antall ørner? Begrunn svaret ditt.
Svar: når antallet pinnsvin i laget reduseres, reduseres antallet av alle påfølgende komponenter og til slutt ørnene, det vil si antallet ørner.
3. Se på bildet, som viser et diagram over karbonkretsløpet i naturen. Angi navnet på stoffet angitt med et spørsmålstegn.

Forklaring: Spørsmålstegnet betegner karbondioksid (CO2), siden CO2 dannes ved forbrenning, respirasjon og nedbrytning av organisk materiale, og ved fotosyntese dannes det (og løses også opp i vann).
Svar: karbondioksid (CO2).
4. Peter blandet like mengder enzym og dets substrat i 25 reagensglass. Rørene ble stående i samme tid ved forskjellige temperaturer, og reaksjonshastigheten ble målt. Basert på resultatene av eksperimentet konstruerte Peter en graf (x-aksen viser temperaturen (i grader Celsius), og y-aksen viser reaksjonshastigheten (i vilkårlige enheter).

Beskriv avhengigheten av hastigheten til en enzymatisk reaksjon på temperaturen.
Svar: når temperaturen stiger til 30C, øker reaksjonshastigheten, og begynner deretter å avta. Den optimale temperaturen er 38C.
5. Etabler sekvensen av underordning av elementer av biologiske systemer, starter med den største.
Manglende elementer:
1 person
2. Biceps
3. Muskelcelle
4. Hånd
5. Aminosyre
6. Aktinprotein
Skriv ned den tilsvarende tallrekkefølgen.
Forklaring: Ordner elementer som starter med det høyeste nivået:
mennesket er organisme
hånd - orgel
biceps - stoff
muskelcelle - cellulær
aktinprotein - molekylært (proteiner består av aminosyrer)
aminosyre - molekylær
Svar: 142365.
6. Proteiner utfører mange viktige funksjoner i menneske- og dyrekropper: de forsyner kroppen med byggemateriale, er biologiske katalysatorer eller regulatorer, gir bevegelse og noe transporterer oksygen. For at kroppen ikke skal oppleve problemer, trenger en person 100-120 g protein per dag.
6.1. Ved å bruke tabelldataene, beregne mengden protein som en person fikk under middagen hvis kostholdet hans inkluderte: 20 g brød, 50 g rømme, 15 g ost og 75 g torsk. Avrund svaret til hele tall.
Forklaring: 100 g brød inneholder 7,8 g protein, deretter inneholder 20 g brød 5 ganger mindre protein - 1,56 g rømme inneholder 3 g protein, deretter inneholder 50 g 2 ganger mindre - 1,5 g g ost - 20 g protein, i 15 g ost - 3 g, i 100 g torsk - 17,4 g protein, i 75 g torsk - 13,05 g.
Totalt: 1,56 + 1,5 + 3 + 13,05 = 19,01 (som er omtrent lik 19).
Svar: 19
ELLER
6.1 En person drakk en kopp sterk kaffe som inneholdt 120 mg koffein, som ble fullstendig absorbert og jevnt fordelt gjennom blodet og andre kroppsvæsker. Hos personen som studeres kan volumet av kroppsvæsker betraktes som lik 40 liter. Beregn hvor lenge etter inntak (i timer) koffein vil slutte å virke på denne personen hvis koffein slutter å virke ved en konsentrasjon på 2 mg/l, og konsentrasjonen reduseres med 0,23 mg per time. Avrund svaret ditt til tideler.
Forklaring: 120 mg koffein ble fordelt over hele menneskekroppen i et volum på 40 liter, det vil si at konsentrasjonen ble 3 mg/l. Ved en konsentrasjon på 2 mg/l slutter koffein å virke, det vil si at kun 1 mg/l er effektivt. For å finne ut antall timer deler du 1 mg/l på 0,23 mg (nedgang i konsentrasjon per time), vi får 4,3 timer.
Svar: 4,3 timer.
6.2. Nevn et av enzymene som produseres av kjertlene i fordøyelsessystemet:
Svar: veggene i magen produserer pepsin, som i et surt miljø bryter ned proteiner til dipeptider. Lipase bryter ned lipider (fett). Nukleaser bryter ned nukleinsyrer. Amylase bryter ned stivelse. Maltase bryter ned maltose til glukose. Laktase bryter ned laktose til glukose og galaktose. Du må skrive ett enzym.
7. Bestem opprinnelsen til sykdommene som er oppført. Skriv ned tallene for hver sykdom på listen i den aktuelle cellen i tabellen. Flere tall kan skrives i tabellceller.
Liste over menneskelige sykdommer:
1. Hemofili
2. Vannkopper
3. Skjørbuk
4. Hjerteinfarkt
5. Kolera
Forklaring: Se Human Diseases for CDF
8. Den genealogiske metoden er mye brukt i medisinsk genetikk. Det er basert på å kompilere en persons stamtavle og studere arven til en bestemt egenskap. I slike studier brukes visse notasjoner. Studer et fragment av slektstreet til en familie, hvor noen av medlemmene har en sammensmeltet øreflipp.

Bruk det foreslåtte opplegget for å finne ut om denne egenskapen er dominant eller recessiv og om den er knyttet til kjønnskromosomer.
Forklaring: egenskapen er recessiv, siden den i første generasjon ikke vises i det hele tatt, og i andre generasjon vises den bare hos 33% av barna. Egenskapen er ikke kjønnsbundet, slik den vises hos både gutter og jenter.
Svar: recessiv, ikke kjønnsbundet.
9. Vladimir ønsket alltid å ha grovt hår som sin far (dominerende egenskap (A)). Men håret hans var mykt, som morens. Bestem genotypene til familiemedlemmer basert på hårkvalitet. Skriv inn svarene dine i tabellen.
Forklaring: mykt hår er en recessiv egenskap (a), far er heterozygot for denne egenskapen, siden sønnen er homozygot recessiv (aa), som moren. Det er:
R: Aa x aa
G: Ah, a x a
F1: Aa - 50 % av barn med grovt hår
aa - 50 % av barn med mykt hår.
Svar:
| Mor | Far | Sønn |
| ahh | Ahh | ahh |
10. Ekaterina bestemte seg for å donere blod som donor. Da hun tok blod, viste det seg at Catherine hadde gruppe III. Ekaterina vet at moren hennes har blodtype I.

10.1. Hva slags blod kan Catherines far ha?
Forklaring: Basert på dataene i tabellen kan Catherines far ha blodtype III eller IV.
Svar: III eller IV.
10.2. Basert på reglene for blodoverføring, avgjør om Catherine kan være blodgiver for faren sin.

Forklaring: Ekaterina, med blodtype I, er en universell donor (forutsatt at Rh-faktorene samsvarer), det vil si at blod kan overføres fra henne til faren.
Svar: kanskje.
11. Funksjonen til organellen vist i figuren er oksidasjon av organiske stoffer og lagring av energi under syntesen av ATP. Den indre membranen til denne organellen spiller en viktig rolle i disse prosessene.

11.1. Hva er navnet på denne organellen?
Svar: Bildet viser en mitokondrier.
11.2. Forklar hvordan pakkingen av den indre membranen i en organell henger sammen med funksjonen den utfører.
Svar: ved hjelp av folder på den indre membranen øker den den indre overflaten av organellen og et større antall organiske stoffer kan oksideres, samt at en større mengde ATP kan produseres av ATP-syntaser - enzymatiske komplekser som produserer energi i form av ATP (hovedenergimolekylet).
12. mRNA-fragmentet har følgende sekvens:
UGTSGAAUGUUUUGTSUG
Bestem sekvensen til DNA-seksjonen som fungerte som mal for syntesen av dette RNA-molekylet, og sekvensen til proteinet som er kodet av dette fragmentet av mRNA. Når du fullfører oppgaven, bruk regelen for komplementaritet og den genetiske kodetabellen.

Regler for bruk av tabellen
Det første nukleotidet i tripletten er tatt fra den venstre vertikale raden; den andre - fra den øverste horisontale raden og den tredje - fra den høyre vertikale raden. Der linjene som kommer fra alle tre nukleotidene krysser hverandre, er den ønskede aminosyren lokalisert.
Forklaring: del sekvensen inn i tripletter (tre nukleotider hver): UGC GAA UGU UUG TsUG. La oss skrive den tilsvarende sekvensen av nukleotider i DNA (omvendt komplementær sekvens av nukleotider, tar i betraktning at A-T (i RNA U), G-C.
Det vil si DNA-kjeden: ACG CTT ACA AAU GAU.
Ved å bruke RNA-sekvensen finner vi den tilsvarende aminosyresekvensen. Den første aminosyren er cis, deretter glu, cis, leu, leu.
Protein: cis-glu-cis-ley-ley.
12.3. Ved dechiffrering av tomatgenomet ble det funnet at i et fragment av et DNA-molekyl er andelen tymin 20 %. Ved å bruke Chargaffs regel, som beskriver de kvantitative forholdene mellom ulike typer nitrogenholdige baser i DNA (G+T = A+C), beregner du mengden (i%) av nukleotider med cytosin i denne prøven.
Forklaring: hvis mengden av tymin er 20 %, er mengden adenin også 20 % (siden de er komplementære). 60 % gjenstår for guanin og cytosin (100 - (20 + 20)), det vil si 30 % hver.
Svar: cytosin utgjør 30 %.
13. Moderne evolusjonsteori kan representeres som følgende diagram.

Svar: antagelig hadde forfedrene til sjiraffen forskjellig halslengde, men siden sjiraffer trengte å nå høyt voksende grønne blader, var det bare sjiraffer med lang hals som overlevde, det vil si de som passet best (denne egenskapen ble festet fra generasjon til generasjon, dette førte til en endring i den genetiske sammensetningen av befolkningen). Under naturlig utvalg overlevde altså bare individer med de lengste halsene, og halsens lengde økte gradvis.
14. Bildet viser cordaite, en utdødd treaktig gymnosperm-plante som levde for 370-250 millioner år siden.

Bruk et fragment av en geokronologisk tabell for å bestemme epoken og periodene der denne organismen levde. Hvilke planter var deres mulige forfedre?
Geokronologisk tabell

Forklaring: Gymnospermer dukket sannsynligvis opp under paleozoikum. perioder: Perm, Karbon (muligens devon). De oppsto fra trebregner (mer primitive planter blomstret i paleozoikumtiden, og gymnospermer spredte seg vidt og blomstret i mesozoikum).
Tid: Paleozoikum
Perioder: Perm, Karbon, Devon
Mulige forfedre: trebregner
2 018 Federal Service for Supervision in Education and Science of the Russian Federation
Den 05.09.2011 kl. 09:36 sa Limarev:
Limarev V.N.
Avkoding av det menneskelige genom.
Fragment fra boken til L.G. Puchko: "Radietetisk erkjennelse av mennesket"
For å løse problemet med å dechiffrere genomet, ble et internasjonalt prosjekt "menneskelig genom" organisert med et budsjett på milliarder av dollar.
I 2000 var det menneskelige genomet så godt som kartlagt. Genene ble talt, identifisert og registrert i databaser. Dette er enorme mengder informasjon.
Registrering av det menneskelige genomet i digitalisert form tar omtrent 300 terabyte dataminne, noe som tilsvarer 3 tusen harddisker med en kapasitet på 100 gigabyte.
Det viste seg. At en person ikke har hundretusener, som tidligere antatt, men i overkant av 30 tusen gener. Fluen har fruktfluer, det er bare halvparten så mange av dem - omtrent 13 tusen, og musen har nesten samme antall som en person. Det er bare omtrent 1% av gener som er unike for mennesker i det dechiffrerte genomet. Det meste av DNA-helixen, som det viste seg, er ikke okkupert av gener, men av såkalte "tomme seksjoner", der gener rett og slett ikke er kodet, så vel som doble fragmenter som gjentas etter hverandre, betydningen og betydningen av som er uklart.
Kort sagt, gener viste seg ikke engang å være livets byggesteiner, men bare elementer av blåkopien som bygningen av kroppen er bygget i henhold til. Byggesteiner, som generelt ble antatt før fremveksten av genetikk, er proteiner.
Det har blitt helt åpenbart at 1% av gener som er unike for mennesker, ikke kan kode for en så enorm mengde informasjon som skiller en person fra en mus. Hvor er all informasjonen lagret? For mange forskere blir det faktum ubestridelig at uten det guddommelige prinsippet er det umulig å forklare menneskets natur. En rekke forskere antyder at det, innenfor rammen av eksisterende ideer om menneskekroppen, i prinsippet er umulig å tyde det menneskelige genomet.
Verden er ikke kjent - den er kjent (mine kommentarer til artikkelen).
1) Tenk på fragmentet: "Uten det guddommelige prinsippet er det umulig å forklare menneskets natur."
Informasjonen presentert ovenfor indikerer ikke på noen måte dette.
Genomet har faktisk en mer kompleks struktur enn tidligere antatt.
Men tross alt består datamaskinen som er nevnt i artikkelen ikke bare av minneceller.
En datamaskin har to minner: langsiktig og operativ, samt en prosessor der informasjon behandles. Det elektromagnetiske feltet er også involvert i informasjonsbehandling. For å dechiffrere genominformasjon, er det nødvendig å forstå hvordan det oppstår, ikke bare lagring av informasjon, men også behandlingen. Jeg innrømmer også ideen om at noe av informasjonen lagres registrert gjennom et elektromagnetisk felt. Og også utenfor en person, som jeg allerede skrev, i spesielle informasjonssentre for Supreme Mind.
Bare forestill deg en kontinuerlig tekst kodet i binær kode 0 eller 1 i morsekode, mens du ikke vet hvilket språk den er skrevet på (engelsk eller fransk....) og du vet ikke at denne kontinuerlige teksten består av ord, setninger , avsnitt, kapitler, bind, hyller, skap osv.
Det er nesten det samme i biologi, bare alt her er kodet med en firesifret kode og vi har så langt dechiffrert rekkefølgen på elementære gener + - / *, men vi kan ikke språket og følgelig ord, setninger, avsnitt, kapitler, bind, hyller, skap osv. For oss er det dechiffrerte genomet fortsatt en solid tekst med 4-gradskode, og det er nesten umulig å studere det hele direkte.
Men det viser seg at i visse tidsperioder (både i individet og hans kohort av generasjoner og i arten, slekten) noen gener og deres komplekser (ansvarlig for ord, setninger, avsnitt, kapitler, bind, hyller, skap osv. .) er aktive , og i andre evolusjonsperioder er de passive, noe jeg indirekte bestemte av ulike polygene egenskaper (som vist i emnet Generell periodisk evolusjonslov).
Det er foreløpig bare to metoder for å studere gener, dette er en enkel laboratorieberegning av summen av gener (DNA) i en prøve og det er en enhet som teller mengden protein RNA som produseres sitter fast på den produserte elektroniske brikken spesifikt DNA, men siden en enorm mengde DNA til enhver tid er aktiv og følgelig et stort antall forskjellige proteiner produseres gjennom RNA, er det svært vanskelig å skille «disse nudlene med en skje, gaffel og japanske spisepinner» i denne suppen og finn det du leter etter - finn årsak-og-virkning-forhold mellom spesifikt DNA (som et DNA-kompleks) og dets innflytelse på en polygen egenskap.
Det ser ut til at jeg har funnet en enkel metode for å sortere ut hele denne suppa av DNA, RNA og deres proteiner som bestemmer graden av en polygen egenskap.
Som det viste seg, er hver polygen egenskap i utviklingsrekkefølgen til et individ (kohort av generasjoner, arter og slekter) periodisk, derfor må den være periodisk i aktiviteten til RNA og DNA, og derfor trenger du bare å finne (først gå inn på genetiske detaljer) korrelasjonen mellom den metriske endringen i den polygene egenskapen (i individ, kohort av generasjoner, arter, slekter...) og den tilsvarende aktiviteten til RNA, DNA, proporsjonal med disse periodene.
Til 50-årsdagen for oppdagelsen av DNA-strukturen
A.V. Zelenin
PLANTEGENOM
A.V. Zelenin
Zelenin Alexander Vladimirovich- Doktor i biologiske vitenskaper,
Laboratoriesjef, Institutt for molekylærbiologi oppkalt etter. V.A. Engelhardt RAS.
De imponerende prestasjonene til Human Genome-programmet, samt suksessen med arbeidet med å tyde de såkalte ultrasmå (virus), små (bakterier, gjær) og mellomstore (rundorm, Drosophila) genomer, gjorde det mulig å gå over til en storstilt studie av store og ekstra store plantegenomer. Det presserende behovet for en detaljert studie av genomene til de økonomisk viktigste plantene ble understreket på et møte om plantegenomikk holdt i 1997 i USA [,]. I løpet av årene siden den gang har utvilsomme suksesser blitt oppnådd på dette området. I 2000 dukket det opp en publikasjon om fullstendig sekvensering (etablering av den lineære nukleotidsekvensen til alt kjernefysisk DNA) av genomet til den lille sennep - Arabidopsis, og i 2001 - om den foreløpige (utkastet) sekvenseringen av risgenomet. Arbeid med å sekvensere store og ultrastore plantegenomer (mais, rug, hvete) har blitt rapportert gjentatte ganger, men disse meldingene inneholdt ikke spesifikk informasjon og var snarere intensjonserklæringer.
Det er forventet at dechiffrering av plantegenomer vil åpne for brede muligheter for vitenskap og praksis. Først av alt vil identifisering av nye gener og kjeden av deres genetiske regulering øke planteproduktiviteten betydelig gjennom bruk av bioteknologiske tilnærminger. Oppdagelsen, isoleringen, reproduksjonen (kloningen) og sekvenseringen av gener som er ansvarlige for slike viktige funksjoner til planteorganismen som reproduksjon og produktivitet, variasjonsprosesser, motstand mot ugunstige miljøfaktorer, samt homolog sammenkobling av kromosomer, er assosiert med fremveksten av nye muligheter for å forbedre utvelgelsesprosessen. Til slutt kan isolerte og klonede gener brukes til å oppnå transgene planter med fundamentalt nye egenskaper og analysere mekanismene for regulering av genaktivitet.
Viktigheten av å studere plantegenomer understrekes også av det faktum at så langt er antallet lokaliserte, klonede og sekvenserte plantegener lite og varierer etter ulike estimater mellom 800 og 1200. Dette er 10-15 ganger mindre enn, for f.eks. for eksempel hos mennesker.
USA er fortsatt den utvilsomme lederen i storstilt studie av plantegenomer, selv om det utføres intensiv forskning på risgenomet i Japan, og de siste årene i Kina. I tillegg til amerikanske laboratorier, tok europeiske forskningsgrupper en aktiv del i å tyde Arabidopsis-genomet. Den tilsynelatende ledelsen i USA skaper alvorlig bekymring blant europeiske forskere, noe de tydelig ga uttrykk for på et møte med meningsfull tittel "Prospects for Genomics in the Postgenomic Era", holdt i Frankrike sent i 2000. Amerikansk vitenskaps fremskritt når det gjelder å studere genomene til landbruksplanter og skape transgene planteformer, truer ifølge europeiske forskere at i en ikke så fjern fremtid (fra to til fem tiår), når befolkningsveksten vil sette menneskeheten i møte med en generell matkrise vil europeisk økonomi og vitenskap bli avhengig av amerikansk teknologi. I denne forbindelse ble det kunngjort opprettelsen av et fransk-tysk vitenskapelig program for studiet av plantegenomer (Plantgene) og investering av betydelige midler i det.
Åpenbart bør problemene med plantegenomikk tiltrekke seg oppmerksomhet fra russiske forskere og vitenskapsarrangører, så vel som styrende organer, siden vi snakker ikke bare om vitenskapelig prestisje, men også om landets nasjonale sikkerhet. Om ett eller to tiår vil mat bli den viktigste strategiske ressursen.
VANSKELIGHETER MED Å STUDERE PLANTEGENOMER
Å studere plantegenomer er en mye mer kompleks oppgave enn å studere genomet til mennesker og andre dyr. Dette skyldes følgende omstendigheter:
enorme genomstørrelser, som når titalls og til og med hundrevis av milliarder av nukleotidpar (bp) for individuelle plantearter: genomene til de viktigste økonomisk viktige plantene (unntatt ris, lin og bomull) er enten i størrelse med det menneskelige genomet eller overskrider det. mange ganger (tabell);KROMOSOMALE GENOMSTUDIERKraftige svingninger i antall kromosomer i ulike planter – fra to hos noen arter til flere hundre hos andre, og det er ikke mulig å identifisere en streng sammenheng mellom genomstørrelsen og antall kromosomer;
En overflod av polyploide (som inneholder mer enn to genomer per celle) dannes med lignende, men ikke identiske genomer (allopolyploidi);
Den ekstreme anrikningen av plantegenomer (opptil 99%) med "ubetydelig" (ikke-kodende, det vil si som ikke inneholder gener) DNA, noe som i stor grad kompliserer sammenføyningen (arrangementet i riktig rekkefølge) av sekvenserte fragmenter til en felles stor- DNA-region i størrelse (kontig);
Ufullstendig (sammenlignet med genomene til Drosophila, mennesker og mus) morfologisk, genetisk og fysisk kartlegging av kromosomer;
Den praktiske umuligheten av å isolere individuelle kromosomer i ren form ved bruk av metoder som vanligvis brukes til dette formålet for kromosomer fra mennesker og dyr (strømsortering og bruk av cellehybrider);
Vanskeligheten med kromosomkartlegging (bestemme plasseringen på kromosomet) av individuelle gener ved bruk av hybridisering på stedet, på grunn av både det høye innholdet av "ubetydelig" DNA i plantegenomer og særegenhetene ved den strukturelle organiseringen av plantekromosomer;
Den evolusjonære avstanden mellom planter og dyr, som i alvorlig grad kompliserer bruken av informasjon hentet fra sekvensering av genomet til mennesker og andre dyr for å studere plantegenomer;
Den lange prosessen med reproduksjon av de fleste planter, noe som reduserer deres genetiske analyse betydelig.
Kromosomale (cytogenetiske) studier av genomer generelt og planter spesielt har en lang historie. Begrepet "genom" ble foreslått for å betegne et haploid (enkelt) sett av kromosomer med genene de inneholder i første kvartal av det 20. århundre, det vil si lenge før rollen til DNA som bærer av genetisk informasjon ble etablert.
Beskrivelse av genomet til en ny, tidligere ustudert genetisk flercellet organisme begynner vanligvis med studiet og beskrivelsen av det komplette settet av kromosomene (karyotype). Dette gjelder selvfølgelig også for planter, et stort antall av dem har ikke engang begynt å bli studert.
Allerede ved begynnelsen av kromosomstudier ble genomer av beslektede plantearter sammenlignet basert på analysen av meiotisk konjugering (forening av homologe kromosomer) i interspesifikke hybrider. I løpet av de siste 100 årene har evnen til kromosomanalyse utvidet seg dramatisk. I dag brukes mer avanserte teknologier for å karakterisere plantegenomer: ulike varianter av den såkalte differensialfargingen, som gjør det mulig å identifisere individuelle kromosomer basert på morfologiske egenskaper; hybridisering på stedet, gjør det mulig å lokalisere spesifikke gener på kromosomer; biokjemiske studier av cellulære proteiner (elektroforese og immunkjemi) og til slutt et sett med metoder basert på analyse av kromosomalt DNA frem til dets sekvensering.

Ris. 1. Karyotyper av korn: a - rug (14 kromosomer), b - durumhvete (28 kromosomer), c - myk hvete (42 kromosomer), d - bygg (14 kromosomer)Karyotypene til korn, først og fremst hvete og rug, har blitt studert i mange år. Det er interessant at i forskjellige arter av disse plantene er antallet kromosomer forskjellig, men alltid et multiplum av syv. Individuelle kornarter kan identifiseres pålitelig ved deres karyotype. For eksempel består ruggenomet av syv par store kromosomer med intenst fargede heterokromatiske blokker i endene, ofte kalt segmenter eller bånd (fig. 1a). Hvete-genomene har allerede 14 og 21 par kromosomer (fig. 1, b, c), og fordelingen av heterokromatiske blokker i dem er ikke den samme som i rugkromosomer. De individuelle genomene til hvete, betegnet A, B og D, skiller seg også fra hverandre En økning i antall kromosomer fra 14 til 21 fører til en skarp endring i hvetens egenskaper, noe som gjenspeiles i navnene deres: durum. eller makaroni, hvete og myk, eller brød, hvete . D-genet, som inneholder gener for glutenproteiner, er ansvarlig for tilegnelsen av høye bakeegenskaper av myk hvete, som gir deigen den såkalte spiringen. Det er dette genomet som vies spesiell oppmerksomhet i seleksjonsforbedring av brødhvete. En annen 14-kromosom kornblanding, bygg (fig. 1, d), brukes vanligvis ikke til å lage brød, men den fungerer som hovedråstoffet for produksjon av så vanlige produkter som øl og whisky.
Kromosomene til noen ville planter som brukes til å forbedre kvaliteten til de viktigste jordbruksartene, for eksempel de ville slektningene til hveten - Aegilops, studeres intensivt. Nye planteformer skapes gjennom kryssing (fig. 2) og utvalg. De siste årene har betydelige forbedringer i forskningsmetoder gjort det mulig å begynne å studere genomene til planter hvis karyotypetrekk (hovedsakelig små kromosomstørrelser) gjorde dem tidligere utilgjengelige for kromosomanalyse. Først nylig ble derfor alle kromosomer av bomull, kamille og lin identifisert for første gang.

Ris. 2. Karyotyper av hvete og hvete-Aegilops hybrid
a - heksaploid vanlig hvete ( Triticum astivum), bestående av A-, B- og O-genomer; b - tetraploid hvete ( Triticum timopheevi), bestående av A- og G-genom. inneholder gener for resistens mot de fleste hvetesykdommer; c - hybrider Triticum astivum X Triticum timopheevi, motstandsdyktig mot meldugg og rust, er erstatningen av deler av kromosomene tydelig synligPRIMÆR STRUKTUR AV DNA
Etter hvert som molekylær genetikk utviklet seg, utvidet selve konseptet med et genom. Nå tolkes dette begrepet både i klassisk kromosomal og i moderne molekylær forstand: hele genetiske materialet til et individuelt virus, celle og organisme. Naturligvis, etter å ha studert den komplette primærstrukturen til genomer (som den komplette lineære sekvensen av nukleinsyrebaser ofte kalles) til en rekke mikroorganismer og mennesker, kom spørsmålet om sekvensering av plantegenomer opp.
Av de mange planteorganismene ble to valgt for studier - Arabidopsis, som representerer klassen av tofrøbladede blader (genomstørrelse 125 millioner bp), og ris fra klassen av enfrøbladede blader (420-470 millioner bp). Disse genomene er små sammenlignet med andre plantegenomer og inneholder relativt få gjentatte seksjoner av DNA. Slike egenskaper ga håp om at de utvalgte genomene ville være tilgjengelige for relativt rask bestemmelse av deres primære struktur.

Ris. 3. Arabidopsis - liten sennep - en liten plante fra korsblomstfamilien ( Brassicaceae). På et område som i areal tilsvarer én side i magasinet vårt, kan du vokse opp til tusen individuelle Arabidopsis-organismerGrunnlaget for å velge Arabidopsis var ikke bare den lille størrelsen på genomet, men også den lille størrelsen på organismen, som gjør den lett å dyrke under laboratorieforhold (fig. 3). Vi tok hensyn til dens korte reproduksjonssyklus, takket være hvilken det er mulig å raskt utføre kryssings- og seleksjonseksperimenter, detaljert genetikk, enkel manipulering med endrede vekstforhold (endre saltsammensetningen i jorda, tilsette forskjellige næringsstoffer, etc.) og teste effekten på planter av ulike mutagene faktorer og patogener (virus, bakterier, sopp). Arabidopsis har ingen økonomisk verdi, derfor ble dets genom, sammen med musegenomet, kalt et referansegenom, eller, mindre nøyaktig, et modellgenom.*
* Utseendet til begrepet "modellgenom" i russisk litteratur er et resultat av en unøyaktig oversettelse av det engelske uttrykket modellgenom. Ordet "modell" betyr ikke bare adjektivet "modell", men også substantivet "prøve", "standard", "modell". Det ville vært mer riktig å snakke om et prøvegenom, eller et referansegenom.Intensivt arbeid med sekvensering av Arabidopsis-genomet startet i 1996 av et internasjonalt konsortium som inkluderte vitenskapelige institusjoner og forskningsgrupper fra USA, Japan, Belgia, Italia, Storbritannia og Tyskland. I desember 2000 ble omfattende informasjon tilgjengelig som oppsummerte bestemmelsen av den primære strukturen til Arabidopsis-genomet. For sekvensering brukte vi klassisk, eller hierarkisk, teknologi: først ble individuelle små seksjoner av genomet studert, hvorfra større seksjoner (contigs) ble laget, og i sluttfasen, strukturen til individuelle kromosomer. Kjerne-DNAet til Arabidopsis-genomet er fordelt på fem kromosomer. I 1999 ble resultatene av sekvensering av to kromosomer publisert, og publisering av informasjon om primærstrukturen til de resterende tre fullførte sekvenseringen av hele genomet.
Av 125 millioner nukleotidpar er den primære strukturen på 119 millioner bestemt, som er 92 % av hele genomet. Bare 8 % av Arabidopsis-genomet, som inneholdt store blokker med repeterende DNA-seksjoner, viste seg å være utilgjengelig for studier. Når det gjelder fullstendighet og grundighet av sekvensering av eukaryote genomer, forblir Arabidopsis i de tre beste mesterne sammen med den encellede gjærorganismen Saccharomyces cerevisiae og flercellet dyreorganisme Caenorhabditis eleganse(se tabell).
Rundt 15 tusen individuelle gener som koder for proteiner ble funnet i Arabidopsis-genomet. Omtrent 12 tusen av disse finnes i to kopier per haploid (enkelt) genom, så det totale antallet gener er 27 tusen. Antall gener i Arabidopsis er ikke mye forskjellig fra antallet gener i organismer som mennesker og mus. men størrelsen på genomet er 25-30 ganger mindre. Denne omstendigheten er assosiert med viktige trekk i strukturen til individuelle Arabidopsis-gener og den generelle strukturen til dets genom.
Arabidopsis-gener er kompakte, og inneholder bare noen få eksoner (proteinkodende regioner), atskilt av korte (omtrent 250 bp) ikke-kodende DNA-strekninger (introner). Gapene mellom individuelle gener er i gjennomsnitt 4,6 tusen nukleotidpar. Til sammenligning påpeker vi at menneskelige gener inneholder mange titalls og til og med hundrevis av eksoner og introner, og intergene regioner har størrelser på 10 tusen nukleotidpar eller mer. Det antas at tilstedeværelsen av et lite kompakt genom bidro til den evolusjonære stabiliteten til Arabidopsis, siden dets DNA ble mindre et mål for ulike skadelige midler, spesielt for introduksjon av viruslignende repeterende DNA-fragmenter (transposoner) i genom.
Andre molekylære trekk ved Arabidopsis-genomet inkluderer berikelse av eksoner med guanin og cytosin (44 % i eksoner og 32 % i introner) sammenlignet med dyregener, samt tilstedeværelsen av to ganger gjentatte (dupliserte) gener. Det antas at denne doblingen skjedde som et resultat av fire samtidige hendelser, som besto i dobling (repetisjon) av en del av Arabidopsis-genene, eller fusjon av beslektede genomer. Disse hendelsene, som fant sted for 100-200 millioner år siden, er en manifestasjon av den generelle tendensen til polyploidisering (en multippel økning i antall genomer i en organisme), karakteristisk for plantegenomer. Noen fakta viser imidlertid at i Arabidopsis er de dupliserte genene ikke-identiske og fungerer annerledes, noe som kan skyldes mutasjoner i deres regulatoriske regioner.
Et annet objekt for fullstendig DNA-sekvensering var ris. Genomet til denne planten er også lite (12 kromosomer, noe som gir totalt 420-470 millioner bp), bare 3,5 ganger større enn Arabidopsis. Imidlertid, i motsetning til Arabidopsis, er ris av enorm økonomisk betydning, og er grunnlaget for ernæring for mer enn halvparten av menneskeheten, derfor er ikke bare milliarder av forbrukere viktig interessert i å forbedre egenskapene, men også en hær på flere millioner dollar av mennesker som er aktivt involvert i den svært arbeidskrevende prosessen med å dyrke den.
Noen forskere begynte å studere risgenomet tilbake på 80-tallet av forrige århundre, men dette arbeidet nådde en seriøs skala først på 90-tallet. I 1991 ble et program for å dechiffrere strukturen til risgenomet opprettet i Japan, og kombinerte innsatsen fra mange forskningsgrupper. I 1997, på grunnlag av dette programmet, ble International Rice Genome Project organisert. Deltakerne bestemte seg for å konsentrere innsatsen om å sekvensere en av risunderartene ( Oriza sativajaponica), i studien som allerede hadde oppnådd betydelig fremgang på det tidspunktet. Human Genome-programmet ble et seriøst insentiv og, billedlig talt, en ledestjerne for slikt arbeid.
Som en del av dette programmet ble strategien for "kromosomal" hierarkisk deling av genomet, som deltakerne i det internasjonale konsortiet brukte for å dechiffrere risgenomet, testet. Men hvis, når man studerer det menneskelige genomet, fraksjoner av individuelle kromosomer ble isolert ved hjelp av forskjellige teknikker, ble materiale spesifikt for individuelle riskromosomer og deres individuelle seksjoner oppnådd ved lasermikrodisseksjon (kutte ut mikroskopiske objekter). På objektglasset hvor riskromosomene befinner seg, under påvirkning av en laserstråle, brennes alt unntatt kromosomet eller dets seksjoner beregnet for analyse ut. Det gjenværende materialet brukes til kloning og sekvensering.
Tallrike rapporter er publisert om resultatene av sekvensering av individuelle fragmenter av risgenomet, utført med høy nøyaktighet og detaljer som er karakteristiske for hierarkisk teknologi. Det ble antatt at bestemmelsen av den fullstendige primærstrukturen til risgenomet ville være fullført innen utgangen av 2003-midten av 2004, og resultatene, sammen med data om den primære strukturen til Arabidopsis-genomet, ville bli mye brukt i den komparative genomikken. av andre planter.
Tidlig i 2002 publiserte imidlertid to forskningsgrupper - den ene fra Kina, den andre fra Sveits og USA - resultatene av fullstendig grov (grov) sekvensering av risgenomet, utført ved bruk av totalkloningsteknologi. I motsetning til en trinn-for-trinn (hierarkisk) studie, er den totale tilnærmingen basert på samtidig kloning av hele det genomiske DNA i en av de virale eller bakterielle vektorene og oppnå et betydelig (stort for middels og store genomer) antall individuelle kloner som inneholder forskjellige DNA-segmenter. Basert på analysen av disse sekvenserte seksjonene og overlappingen av identiske endedeler av DNA, dannes en contig - en kjede av DNA-sekvenser koblet sammen. Den generelle (totale) contig representerer den primære strukturen til hele genomet eller i det minste til et individuelt kromosom.
I en slik skjematisk presentasjon virker strategien med total kloning ukomplisert. Faktisk møter den alvorlige vanskeligheter knyttet til behovet for å få et stort antall kloner (det er generelt akseptert at genomet eller regionen som studeres må overlappes av kloner minst 10 ganger), et gigantisk volum av sekvensering og ekstremt komplekst arbeid med å slå sammen kloner, som krever deltakelse fra bioinformatikkspesialister. En alvorlig hindring for total kloning er mangfoldet av repeterende DNA-regioner, hvor antallet, som allerede nevnt, øker kraftig ettersom genomstørrelsen øker. Derfor brukes den totale sekvenseringsstrategien først og fremst til å studere genomene til virus og mikroorganismer, selv om den ble brukt med hell for å studere genomet til en flercellet organisme, Drosophila.
Resultatene av total sekvensering av dette genomet ble "overlagret" på et stort utvalg informasjon om dets kromosomale, gen og molekylære struktur oppnådd over en nesten 100-års periode med studier av Drosophila. Og likevel, når det gjelder graden av sekvensering, er Drosophila-genomet (66% av den totale genomstørrelsen) betydelig dårligere enn Arabidopsis-genomet (92%), til tross for deres ganske like størrelser - henholdsvis 180 millioner og 125 millioner nukleotidpar. . Derfor har det nylig blitt foreslått å kalle teknologien som brukes til å sekvensere Drosophila-genomet blandet.
For å sekvensere risgenomet tok de ovennevnte forskningsgruppene to av underartene, den mest dyrkede i asiatiske land - Oriza spytt L. ssp indicaj Og Oriza spytt L. sspjaponica. Resultatene av deres forskning er sammenfallende på mange måter, men er også forskjellige på mange måter. Dermed uttalte representanter for begge gruppene at de oppnådde contig overlapping på omtrent 92-93% av genomet. Det er vist at omtrent 42 % av risgenomet er representert av korte DNA-repetisjoner bestående av 20 nukleotidpar, og flertallet av mobile DNA-elementer (transposoner) er lokalisert i intergene regioner. Informasjon om størrelsen på risgenomet varierer imidlertid betydelig.
For den japanske underarten er genomstørrelsen bestemt til å være 466 millioner nukleotidpar, og for den indiske underarten - 420 millioner årsaken til denne uoverensstemmelsen er ikke klar. Det kan være en konsekvens av ulike metodiske tilnærminger for å bestemme størrelsen på den ikke-kodende delen av genomene, det vil si at den kanskje ikke gjenspeiler den sanne tilstanden. Men det er mulig at det virkelig eksisterer en 15 % forskjell i størrelsen på de studerte genomene.
Det andre alvorlige avviket ble avslørt i antall oppdagede gener: for den japanske underarten - fra 46 022 til 55 615 gener per genom, og for den indiske underarten - fra 32 000 til 50 000 årsaken til dette avviket.
Ufullstendigheten og inkonsekvensen av den mottatte informasjonen er notert i kommentarene til de publiserte artiklene. Det er også håp om at gapene i kunnskap om risgenomet vil bli eliminert ved å sammenligne dataene fra "grov sekvensering" med resultatene av detaljert, hierarkisk sekvensering utført av deltakere i International Rice Genome Project.
SAMMENLIGNENDE OG FUNKSJONELL GENOMI AV PLANTER
De omfattende dataene som er oppnådd, hvorav halvparten (resultatene fra den kinesiske gruppen) er offentlig tilgjengelige, åpner utvilsomt for brede perspektiver både for studiet av risgenomet og for plantegenomikk generelt. En sammenligning av egenskapene til Arabidopsis- og risgenomene viste at de fleste genene (opptil 80%) identifisert i Arabidopsis-genomet også finnes i risgenomet, men for omtrent halvparten av genene som finnes i ris, er analoger ( ortologer) har ennå ikke blitt funnet i Arabidopsis-genomet. Samtidig er 98 % av gener hvis primærstruktur er etablert for andre kornsorter, blitt identifisert i risgenomet.
Den betydelige (nesten to ganger) avviket i antall gener i ris og Arabidopsis er forvirrende. Samtidig er dataene fra det grove transkripsjonen av risgenomet, oppnådd ved hjelp av total sekvensering, praktisk talt ikke sammenlignet med de omfattende resultatene av å studere risgenomet ved hjelp av metoden for hierarkisk kloning og sekvensering, det vil si hva som er gjort. for Drosophila-genomet er ikke oppnådd. Derfor er det fortsatt uklart om forskjellen i antall gener i Arabidopsis og ris gjenspeiler den sanne tilstanden eller er forklart av forskjeller i metodiske tilnærminger.
I motsetning til Arabidopsis-genomet gis ikke informasjon om tvillinggener i risgenomet. Det er mulig at deres relative overflod kan være større i ris enn i Arabidopsis. Denne muligheten støttes indirekte av data om tilstedeværelsen av polyploide former for ris. Større klarhet i dette spørsmålet kan forventes etter fullføringen av International Rice Genome Project og oppnåelse av et detaljert bilde av den primære DNA-strukturen til dette genomet. Alvorlige grunner for et slikt håp er gitt av det faktum at etter publiseringen av arbeider om grov sekvensering av risgenomet, økte antallet publikasjoner om strukturen til dette genomet kraftig, spesielt dukket det opp informasjon om den detaljerte sekvenseringen av kromosomene. 1 og 4.
Å vite, i det minste omtrentlig, antall gener i planter er av grunnleggende betydning for sammenlignende plantegenomikk. Først ble det antatt at siden alle blomstrende planter er svært nær hverandre i sine fenotypiske egenskaper, burde deres genom også være nære. Og hvis vi studerer Arabidopsis-genomet, vil vi få informasjon om de fleste genomene til andre planter. Indirekte bekreftelse av denne antagelsen er gitt av resultatene av sekvensering av musegenomet, som er overraskende nær det menneskelige genomet (omtrent 30 tusen gener, hvorav bare 1 tusen viste seg å være forskjellige).
Det kan antas at årsaken til forskjellene i genomene til Arabidopsis og ris ligger i deres tilhørighet til forskjellige klasser av planter - tofrøbladede og enfrøbladede. For å avklare dette problemet er det ekstremt ønskelig å kjenne i det minste den grove primærstrukturen til en annen enfrøplante. Den mest realistiske kandidaten kan være mais, hvis genom er omtrent lik det menneskelige genomet, men fortsatt betydelig mindre enn genomet til andre kornsorter. Matverdien til mais er velkjent.
Det enorme materialet som oppnås ved å sekvensere genomene til Arabidopsis og ris, blir gradvis grunnlaget for en storstilt studie av plantegenomer ved bruk av komparative genomiske metoder. Slike studier har generell biologisk betydning, da de gjør det mulig å etablere hovedprinsippene for organiseringen av plantegenomet som helhet og deres individuelle kromosomer, å identifisere fellestrekk ved strukturen til gener og deres regulatoriske regioner, og å vurdere forhold mellom den funksjonelt aktive (gen) delen av kromosomet og ulike ikke-proteinkodende intergene DNA-regioner. Komparativ genetikk blir også stadig viktigere for utviklingen av menneskelig funksjonell genomikk. Det er for komparative studier at genomene til pufferfisk og mus ble sekvensert.
Ikke mindre viktig er studiet av individuelle gener som er ansvarlige for syntesen av individuelle proteiner som bestemmer spesifikke funksjoner i kroppen. Det er i påvisning, isolering, sekvensering og etablering av funksjonen til individuelle gener at den praktiske, primært medisinske, betydningen av Human Genome-programmet ligger. Denne omstendigheten ble notert for flere år siden av J. Watson, som understreket at Human Genome-programmet vil bli fullført først når funksjonene til alle menneskelige gener er bestemt.
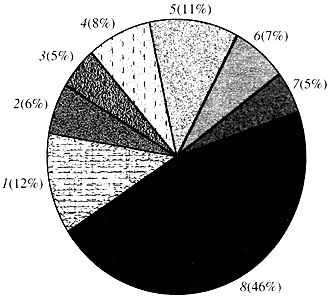
Ris. 4. Klassifisering etter funksjon av Arabidopsis-gener
1 - gener for vekst, deling og DNA-syntese; 2 - RNA-syntese gener (transkripsjon); 3 - gener for proteinsyntese og modifikasjon; 4 - gener for utvikling, aldring og celledød; 5 - gener for cellulær metabolisme og energimetabolisme; 6 - gener for intercellulær interaksjon og signaloverføring; 7 - gener for å støtte andre cellulære prosesser; 8 - gener med ukjent funksjonNår det gjelder funksjonen til plantegener, vet vi mindre enn en tidel av det vi vet om menneskelige gener. Selv i Arabidopsis, hvis genom er mye mer studert enn det menneskelige genomet, forblir funksjonen til nesten halvparten av genene ukjent (fig. 4). I mellomtiden har planter, i tillegg til gener som er felles for dyr, et betydelig antall gener som er spesifikke (eller i det minste hovedsakelig) for dem. Vi snakker om gener involvert i vanntransport og syntese av cellevegger, som er fraværende hos dyr, om gener som sikrer dannelse og funksjon av kloroplaster, fotosyntese, nitrogenfiksering og syntese av en rekke aromatiske produkter. Denne listen kan fortsettes, men det er allerede klart hvor vanskelig oppgaven står overfor plantefunksjonell genomikk.
Fullstendig genomsekvensering gir nær sann informasjon om det totale antallet gener til en gitt organisme, gjør at mer eller mindre detaljert og pålitelig informasjon om deres struktur kan plasseres i databanker, og letter arbeidet med å isolere og studere individuelle gener. Genomsekvensering betyr imidlertid ikke å etablere funksjonen til alle gener.
En av de mest lovende tilnærmingene til funksjonell genomikk er basert på å identifisere fungerende gener som mRNA-transkripsjon (lesing) skjer på. Denne tilnærmingen, inkludert bruk av moderne mikroarray-teknologi, gjør det mulig å identifisere opptil titusenvis av fungerende gener samtidig. Nylig, ved å bruke denne tilnærmingen, har studiet av plantegenomer begynt. For Arabidopsis var det mulig å få rundt 26 tusen individuelle transkripsjoner, noe som i stor grad letter muligheten for å bestemme funksjonen til nesten alle genene. I poteter var det mulig å identifisere rundt 20 000 tusen arbeidsgener som er viktige for å forstå både prosessene med vekst og knolldannelse, og prosessene ved potetsykdom. Det forventes at denne kunnskapen vil forbedre motstanden til et av de viktigste matvarene mot patogener.
En logisk utvikling av funksjonell genomikk er proteomikk. Dette nye vitenskapsfeltet studerer proteomet, som vanligvis refererer til det komplette settet med proteiner i en celle på et gitt tidspunkt. Dette settet med proteiner, som gjenspeiler den funksjonelle tilstanden til genomet, endres hele tiden, mens genomet forblir uendret.
Studiet av proteiner har lenge vært brukt til å gjøre vurderinger om aktiviteten til plantegenomer. Som kjent er enzymer som finnes i alle planter forskjellige i individuelle arter og varianter i sekvensen av aminosyrer. Slike enzymer, med samme funksjon, men forskjellige sekvenser av individuelle aminosyrer, kalles isoenzymer. De har forskjellige fysisk-kjemiske og immunologiske egenskaper (molekylvekt, ladning), som kan påvises ved hjelp av kromatografi eller elektroforese. I mange år har disse metodene vært vellykket brukt for å studere såkalt genetisk polymorfisme, det vil si forskjeller mellom organismer, varianter, populasjoner, arter, spesielt hvete og relaterte former for korn. Men nylig, på grunn av den raske utviklingen av DNA-analysemetoder, inkludert sekvensering, har studiet av proteinpolymorfisme blitt erstattet av studiet av DNA-polymorfisme. Direkte studier av spektra av lagringsproteiner (prolaminer, gliadiner, etc.), som bestemmer de grunnleggende ernæringsmessige egenskapene til korn, er imidlertid fortsatt en viktig og pålitelig metode for genetisk analyse, seleksjon og frøproduksjon av landbruksplanter.
Kunnskap om gener, mekanismene for deres uttrykk og regulering er ekstremt viktig for utvikling av bioteknologi og produksjon av transgene planter. Det er kjent at imponerende suksesser på dette området forårsaker tvetydige reaksjoner fra miljø- og medisinske miljøer. Imidlertid er det et område med plantebioteknologi der denne frykten, om ikke helt grunnløs, så i alle fall virker ubetydelig. Vi snakker om å lage transgene industrianlegg som ikke brukes som matvarer. India høstet nylig sin første avling av transgen bomull som er motstandsdyktig mot en rekke sykdommer. Det er informasjon om innføring av spesielle gener som koder for pigmentproteiner i bomullsgenomet og produksjon av bomullsfibre som ikke krever kunstig farging. En annen industriell avling som kan være gjenstand for effektiv genteknologi er lin. Bruken som et alternativ til bomull for tekstilråvarer har nylig blitt diskutert. Dette problemet er ekstremt viktig for landet vårt, som har mistet sine egne kilder til bomullsråvarer.
UTSIKTER FOR Å STUDERE PLANTEGENOMER
Det er åpenbart at strukturelle studier av plantegenomer vil være basert på tilnærminger og metoder for komparativ genomikk ved å bruke resultatene av dechiffrering av genomene til Arabidopsis og ris som hovedmateriale. En betydelig rolle i utviklingen av komparativ plantegenomikk vil uten tvil bli spilt av informasjonen som før eller siden vil bli gitt ved total (grov) sekvensering av genomene til andre planter. I dette tilfellet vil sammenlignende plantegenomikk være basert på å etablere genetiske forhold mellom individuelle loki og kromosomer som tilhører ulike genom. Vi vil ikke snakke så mye om den generelle genomikken til planter, men om den selektive genomikken til individuelle kromosomale loki. Dermed ble det nylig vist at genet som er ansvarlig for vernalisering er lokalisert i VRn-AI-lokuset til kromosom 5A av heksaploid hvete og Hd-6-lokuset til kromosom 3 av ris.
Utviklingen av disse studiene vil være en kraftig drivkraft for identifikasjon, isolering og sekvensering av mange funksjonelt viktige plantegener, spesielt gener som er ansvarlige for sykdomsresistens, tørkeresistens og tilpasningsevne til ulike vekstforhold. Funksjonell genomikk, basert på masseidentifikasjon (screening) av gener som fungerer i planter, vil bli stadig mer brukt.
Vi kan forutse ytterligere forbedringer i kromosomteknologier, først og fremst mikrodisseksjonsmetoden. Bruken utvider dramatisk mulighetene for genomisk forskning uten å kreve store kostnader, for eksempel total genomsekvensering. Metoden for å lokalisere individuelle gener på plantekromosomer ved bruk av hybridisering vil bli mer utbredt. på stedet. For øyeblikket er bruken begrenset av det enorme antallet repeterende sekvenser i plantegenomet, og muligens av særegenhetene ved den strukturelle organiseringen av plantekromosomer.
I overskuelig fremtid vil også kromosomteknologier få stor betydning for den evolusjonære genomikken til planter. Disse teknologiene, som er relativt rimelige, gjør det mulig å raskt vurdere intra- og interspesifikk variabilitet og studere komplekse allopolyploide genomer av tetraploid og heksaploid hvete og triticale; analysere evolusjonære prosesser på kromosomnivå; undersøke dannelsen av syntetiske genomer og introduksjonen (introgresjon) av fremmed genetisk materiale; identifisere genetiske forhold mellom individuelle kromosomer av forskjellige arter.
Studiet av plantekaryotype ved bruk av klassiske cytogenetiske metoder, beriket med molekylærbiologisk analyse og datateknologi, vil bli brukt til å karakterisere genomet. Dette er spesielt viktig for å studere stabiliteten og variasjonen til karyotypen på nivået av ikke bare individuelle organismer, men også populasjoner, varianter og arter. Til slutt er det vanskelig å forestille seg hvordan man kan estimere antall og spektra av kromosomale omorganiseringer (aberrasjoner, broer) uten bruk av differensielle fargingsmetoder. Slike studier er ekstremt lovende for overvåking av miljøet basert på tilstanden til plantegenomet.
I det moderne Russland er det usannsynlig at direkte sekvensering av plantegenomer vil bli utført. Slikt arbeid, som krever store investeringer, er uholdbart for vår nåværende økonomi. I mellomtiden er informasjon om strukturen til genomene til Arabidopsis og ris, oppnådd av verdensvitenskap og tilgjengelig i internasjonale databanker, tilstrekkelig for utvikling av innenlandsk plantegenomikk. Det er mulig å forutse en utvidelse av forskningen på plantegenomer basert på komparative genomiske tilnærminger for å løse spesifikke problemer med avl og avlingsproduksjon, samt å studere opprinnelsen til ulike plantearter av økonomisk betydning.
Det kan antas at i innenlandsk avlspraksis og plantedyrking vil genomiske tilnærminger som genetisk typing (RELF, RAPD, AFLP-analyser, etc.), som er ganske rimelige for vårt budsjett, bli mye brukt. Parallelt med direkte metoder for å bestemme DNA-polymorfisme, vil tilnærminger basert på studiet av proteinpolymorfisme, primært lagringsproteiner av korn, bli brukt for å løse problemer med genetikk og planteavl. Kromosomteknologier vil bli mye brukt. De er relativt rimelige, og utviklingen krever ganske moderate investeringer. Innenfor kromosomforskning er ikke innenlandsvitenskapen dårligere enn verden.
Det bør understrekes at vår vitenskap har gitt et betydelig bidrag til dannelsen og utviklingen av plantegenomikk [,].
Den grunnleggende rollen ble spilt av N.I. Vavilov (1887-1943).
Innen molekylærbiologi og plantegenomikk er det banebrytende bidraget til A.N. Belozersky (1905-1972).
Innenfor kromosomforskning er det nødvendig å merke seg arbeidet til den fremragende genetikeren S.G. Navashin (1857-1930), som først oppdaget satellittkromosomer i planter og beviste at det er mulig å skille individuelle kromosomer ut fra egenskapene til deres morfologi.
En annen klassiker innen russisk vitenskap G.A. Levitsky (1878-1942) beskrev i detalj kromosomene til rug, hvete, bygg, erter og sukkerroer, introduserte begrepet "karyotype" i vitenskapen og utviklet læren om det.
Moderne spesialister, som er avhengige av prestasjonene til verdensvitenskapen, kan gi et betydelig bidrag til den videre utviklingen av plantegenetikk og genomikk.
Forfatteren uttrykker sin dyptfølte takknemlighet til akademiker Yu.P. Altukhov for kritisk diskusjon av artikkelen og verdifulle råd.Arbeidet til teamet ledet av forfatteren av artikkelen ble støttet av Russian Foundation for Basic Research (stipend nr. 99-04-48832; 00-04-49036; 00-04-81086), programmet til presidenten for den russiske føderasjonen for støtte til vitenskapelige skoler (tilskudd nr. 00-115 -97833 og NSh-1794.2003.4) og programmet til det russiske vitenskapsakademiet "Molekylærgenetiske og kromosomale markører i utviklingen av moderne metoder for seleksjon og frø produksjon."
LITTERATUR
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Introduksjon til plantegenomikk // Molekylærbiologi. 2001. T. 35. s. 339-348. 2. Penn E. Bonanza for Plant Genomics // Vitenskap. 1998. V. 282. S. 652-654. 3. Plantegenomikk // Proc. Natl. Acad. Sci. USA. 1998. V. 95. S. 1962-2032. 4. Kartel N.A. og så videre. Genetikk. Encyklopedisk ordbok. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Genomdifferensiering i Aegilops. 1. Distribusjon av svært repeterende DNA-sekvenser på kromosomer av diploide arter // Genom. 1996. V. 39. S. 293-306. Historie om kromosomanalyse // Biol. membraner. 2001. T. 18. s. 164-172.



