Vannmiljø.
Fordeling av organismer etter livsmiljø
I ferd med lang historisk utvikling levende materie og dannelsen av stadig mer avanserte former for levende vesener, organismer, som mestrer nye habitater, ble fordelt på jorden i henhold til dens mineralskjell (hydrosfære, litosfære, atmosfære) og tilpasset tilværelsen under strengt definerte forhold.
Det første livsmediet var vann. Det var i den livet oppsto. Etter hvert som den historiske utviklingen skred frem, begynte mange organismer å befolke land-luft-miljøet. Som et resultat dukket det opp landplanter og dyr, som raskt utviklet seg og tilpasset seg nye levekår.
I prosessen med å fungere av levende materie på land, ble overflatelagene til litosfæren gradvis forvandlet til jord, til en slags, som V.I. Vernadsky sa det, en bioinert kropp av planeten. Både vannlevende og terrestriske organismer begynte å befolke jorden, og skapte et spesifikt kompleks av innbyggerne.
På den moderne jorda skilles fire livsmiljøer klart - vannlevende, land-luft-, jord- og levende organismer - som er betydelig forskjellige i deres forhold. La oss se på hver av dem.
Generelle egenskaper. Livets vannmiljø, hydrosfæren, opptar opptil 71 % av klodens areal. Når det gjelder volum, er vannreservene på jorden estimert til 1370 millioner kubikkmeter. km, som er 1/800 av klodens volum. Hovedmengden vann, mer enn 98%, er konsentrert i hav og hav, 1,24% er representert av isen i polarområdene; i ferskvann i elver, innsjøer og sumper overstiger ikke vannmengden 0,45 %.
Vannmiljøet er hjemsted for rundt 150 000 dyrearter (omtrent 7 % av det totale antallet på kloden) og 10 000 plantearter (8 %). Til tross for det faktum at representanter for det store flertallet av grupper av planter og dyr forble i vannmiljøet (i deres "vugge"), er antallet av deres arter mye mindre enn for landlevende. Dette betyr at utviklingen på land skjedde mye raskere.
Den mest mangfoldige og rike planten og dyreverden hav og hav i de ekvatoriale og tropiske områdene (spesielt Stillehavet og Atlanterhavet). Sør og nord for disse beltene blir den kvalitative sammensetningen av organismer gradvis utarmet. I området til den østindiske skjærgården er det rundt 40 000 dyrearter, og i Laptevhavet er det bare 400. Dessuten er hoveddelen av organismene i verdenshavet konsentrert i et relativt lite område med havkystene i den tempererte sonen og blant mangrovene i tropiske land. I store vannområder langt fra kysten er det ørkenområder, praktisk talt blottet for liv.
Andelen elver, innsjøer og sumper i biosfæren er ubetydelig sammenlignet med hav og hav. Likevel skaper de tilførsel av ferskvann som er nødvendig for et stort antall planter og dyr, så vel som for mennesker.
Vannmiljøet har sterk innflytelse på innbyggerne. I sin tur påvirker det levende stoffet i hydrosfæren habitatet, behandler det, involverer det i syklusen av stoffer. Det anslås at vannet i hav og hav, elver og innsjøer brytes ned og gjenopprettes i den biotiske syklusen i løpet av 2 millioner år, det vil si at alt har gått gjennom den levende materie på planeten mer enn tusen ganger*. Dermed er den moderne hydrosfæren et produkt av den vitale aktiviteten til levende materie, ikke bare av moderne, men også fra tidligere geologiske epoker.
Et karakteristisk trekk ved vannmiljøet er dets mobilitet selv i stillestående vannmasser, for ikke å nevne rennende, raskt rennende elver og bekker. Havene og havene opplever flo og fjære, kraftige strømmer og stormer; I innsjøer beveger vannet seg under påvirkning av vind og temperatur. Vannets bevegelse sikrer tilførsel av akvatiske organismer med oksygen og næringsstoffer og fører til en utjevning (reduksjon) i temperatur gjennom hele reservoaret.
Innbyggerne i reservoarene har utviklet passende tilpasninger til miljøets mobilitet. For eksempel, i rennende reservoarer er det såkalte "begroende" planter som er godt festet til undervannsobjekter - grønne alger (Cladophora) med et spor av skudd, kiselalger (Diatomeae), vannmoser (Fontinalis), som danner et tett dekke selv på steiner i raske elverifler .
Dyr tilpasset seg også mobiliteten i vannmiljøet. Hos fisk som lever i raskt rennende elver er kroppen nesten rund i tverrsnitt (ørret, ørekyte). De beveger seg vanligvis mot strømmen. Virvelløse dyr av rennende vannforekomster holder seg vanligvis på bunnen, kroppen deres er flatet ut i dorso-ventral retning, mange har forskjellige fikseringsorganer på den ventrale siden, slik at de kan feste seg til undervannsobjekter. I havet oppleves den sterkeste påvirkningen av bevegelige vannmasser av organismer i tidevanns- og surfesonene. På steinete strender i bølgene er det vanlige havkaker (Balanus, Chthamalus), gastropoder (Patella Haliotis) og noen arter av krepsdyr som gjemmer seg i sprekkene i kysten.
I livet til vannlevende organismer på tempererte breddegrader spiller den vertikale bevegelsen av vann i stående reservoarer en viktig rolle. Vannet i dem er tydelig delt inn i tre lag: den øvre epilimnion, hvis temperatur opplever skarpe sesongsvingninger; temperaturhopplag - metalimnion (termoklin), hvor det observeres en skarp temperaturforskjell; bunn dypt lag, hypolimnion, hvor temperaturen endres noe gjennom året.
Om sommeren er de varmeste vannlagene plassert ved overflaten, og de kaldeste ligger på bunnen. Denne lag-for-lag-fordelingen av temperaturer i et reservoar kalles direkte lagdeling. Om vinteren, med en nedgang i temperaturen, observeres en omvendt lagdeling: kaldt overflatevann med temperaturer under 4 ° C ligger over relativt varme. Dette fenomenet kalles temperaturdikotomi. Den er spesielt uttalt i de fleste av våre innsjøer sommer og vinter. Som et resultat av temperaturdikotomi dannes en tetthetsstratifisering av vann i et reservoar, dets vertikale sirkulasjon blir forstyrret og en periode med midlertidig stagnasjon begynner.
Om våren blir overflatevann, på grunn av oppvarming til 4 °C, tettere og synker dypere, og varmere vann stiger opp fra dypet for å ta plass. Som et resultat av slik vertikal sirkulasjon oppstår homotermi i reservoaret, det vil si at temperaturen til hele vannmassen i noen tid utjevnes. Med en ytterligere temperaturøkning blir de øvre vannlagene mindre og mindre tette og synker ikke lenger - sommerstagnasjon setter inn.
Om høsten avkjøles overflatelaget, blir tettere og synker dypere, og fortrenger varmere vann til overflaten. Dette skjer før utbruddet av høsthomotermi. Når overflatevann avkjøles under 4 °C, blir de igjen mindre tett og blir igjen på overflaten. Som et resultat stopper vannsirkulasjonen og vinterstagnasjon oppstår.
Organismer i vannforekomster på tempererte breddegrader er godt tilpasset sesongmessige vertikale bevegelser av vannlag, til vår- og høsthomotermi og til sommer- og vinterstagnasjon (fig. 13).
I innsjøer på tropiske breddegrader synker aldri overflatevannstemperaturen under 4 °C og temperaturgradienten i dem kommer tydelig til uttrykk ned til de dypeste lagene. Blanding av vann skjer som regel her uregelmessig på den kaldeste tiden av året.
Spesielle forhold for livet utvikler seg ikke bare i vannsøylen, men også i bunnen av reservoaret, siden det ikke er lufting i jorda og mineralforbindelser vaskes ut av dem. Derfor har de ikke fruktbarhet og tjener bare som et mer eller mindre fast substrat for vannlevende organismer, og utfører hovedsakelig en mekanisk-dynamisk funksjon. I denne forbindelse får størrelsen på jordpartikler, tettheten av deres kontakt med hverandre og motstand mot utvasking av strømmer den største miljømessige betydningen.
Abiotiske faktorer i vannmiljøet. Vann som levemiljø har spesielle fysiske og kjemiske egenskaper.
Temperaturregimet i hydrosfæren er fundamentalt forskjellig fra det i andre miljøer. Temperatursvingningene i verdenshavet er relativt små: den laveste er omtrent -2 °C, og den høyeste er omtrent 36 °C. Amplituden av oscillasjoner her faller derfor innenfor 38 °C. Med dybden synker temperaturen på vannet i havene. Selv i tropiske områder på 1000 m dyp overstiger den ikke 4–5°C. På dypet av alle hav er det et lag med kaldt vann (fra -1,87 til + 2°C).
I ferske innlandsvannforekomster med tempererte breddegrader varierer temperaturen på overflatelagene av vann fra – 0,9 til +25 ° C, i dypere vann er den 4–5 ° C. Unntaket er termiske kilder, hvor temperaturen på overflatelaget noen ganger når 85–93 °C.
Termodynamiske egenskaper ved vannmiljøet, som høy spesifikk varmekapasitet, høy varmeledningsevne og ekspansjon ved frysing, skaper spesielt gunstige forhold for liv. Disse forholdene er også sikret av den høye latente fusjonsvarmen av vann, som et resultat av at temperaturen under isen om vinteren aldri er under frysepunktet (for ferskvann ca. 0 ° C). Siden vann har størst tetthet ved 4°C, og når det fryser utvider det seg, om vinteren dannes det bare is på toppen, men hovedtykkelsen fryser ikke.
Siden temperaturregimet til vannforekomster er preget av stor stabilitet, er organismene som bor i det preget av en relativ konstant kroppstemperatur og har et smalt spekter av tilpasningsevne til svingninger i miljøtemperaturen. Selv mindre avvik i det termiske regimet kan føre til betydelige endringer i livet til dyr og planter. Et eksempel er den "biologiske eksplosjonen" av lotus (Nelumbium caspium) i den nordligste delen av habitatet - i Volga-deltaet. I lang tid bebodd denne eksotiske planten bare en liten bukt. Bak siste tiåret Arealet med lotuskratt har økt nesten 20 ganger og okkuperer nå over 1500 hektar vannområde. Denne raske spredningen av lotusen forklares av det generelle fallet i nivået av Det kaspiske hav, som ble ledsaget av dannelsen av mange små innsjøer og elvemunninger ved munningen av Volga. I de varme sommermånedene ble vannet her varmere enn før, noe som bidro til veksten av lotuskratt.
Vann er også preget av betydelig tetthet (i denne forbindelse er det 800 ganger større enn luftmediet) og viskositet. Disse funksjonene påvirker plantene ved at deres mekaniske vev utvikler seg veldig svakt eller ikke i det hele tatt, så stilkene deres er veldig elastiske og bøyes lett. De fleste vannplanter er preget av oppdrift og evne til å være suspendert i vannsøylen. De stiger til overflaten og faller så igjen. Hos mange vannlevende dyr er integumentet rikelig smurt med slim, noe som reduserer friksjonen under bevegelse, og kroppen får en strømlinjeformet form.
Organismer i vannmiljøet er fordelt i hele dets tykkelse (i havbunner ble dyr funnet på over 10 000 m dyp). Naturligvis opplever de forskjellige press på forskjellige dyp. Dyphavsdyr er tilpasset høyt trykk (opptil 1000 atm), mens innbyggerne i overflatelagene ikke er utsatt for det. I gjennomsnitt, i vannsøylen, for hver 10 m dybde, øker trykket med 1 atm. Alle hydrobionter er tilpasset denne faktoren og er følgelig delt inn i dypvann og de som lever på grunne dyp.
Vanngjennomsiktighet og dets lysregime har stor innflytelse på vannlevende organismer. Dette påvirker spesielt distribusjonen av fotosyntetiske planter. I gjørmete reservoarer lever de kun i overflatelaget, og der det er større gjennomsiktighet trenger de ned til betydelige dyp. En viss turbiditet av vann skapes av et stort antall partikler suspendert i det, noe som begrenser penetrasjonen av sollys. Turbiditet i vann kan være forårsaket av partikler av mineralske stoffer (leire, silt) og små organismer. Gjennomsiktigheten til vannet avtar også om sommeren med den raske veksten av vannvegetasjon og massereproduksjonen av små organismer suspendert i overflatelagene. Lysregimet til reservoarene avhenger også av sesongen. I nord, på tempererte breddegrader, når reservoarene fryser og isen på toppen fortsatt er dekket med snø, er gjennomtrengningen av lys inn i vannsøylen sterkt begrenset.
Lysregimet bestemmes også av den naturlige reduksjonen i lys med dybden på grunn av at vann absorberer sollys. I dette tilfellet absorberes stråler med forskjellige bølgelengder annerledes: røde absorberes raskest, mens blågrønne trenger inn til betydelige dybder. Havet blir mørkere med dybden. Fargen på miljøet endres, og beveger seg gradvis fra grønnaktig til grønn, deretter til blå, blå, blåfiolett, og gir plass til konstant mørke. Følgelig, med dybde, erstattes grønne alger (Chlorophyta) med brune (Phaeophyta) og røde (Rhodophyta), hvis pigmenter er tilpasset for å fange sollys med forskjellige bølgelengder. Fargen på dyr endres også naturlig med dybden. I overflaten lever vanligvis lyse lag med vann, klare og forskjellig fargede dyr, mens dyphavsarter er blottet for pigmenter. I havets skumringssone lever dyr som er farget med en rødlig fargetone, som hjelper dem å gjemme seg for fiender, siden den røde fargen i blåfiolette stråler oppfattes som svart.
Vannets saltholdighet spiller en viktig rolle i livet til vannlevende organismer. Som du vet, er vann et utmerket løsningsmiddel for mange mineralforbindelser. Som et resultat har naturlige reservoarer en viss kjemisk sammensetning. Høyeste verdi har karbonater, sulfater, klorider. Mengden oppløste salter per 1 liter vann i ferskvannsforekomster overstiger ikke 0,5 g (vanligvis mindre); i hav og hav når den 35 g (tabell 6).
Tabell 6.Distribusjon av basiske salter i ulike reservoarer (ifølge R. Dazho, 1975)
Kalsium spiller en viktig rolle i livet til ferskvannsdyr. Bløtdyr, krepsdyr og andre virvelløse dyr bruker det til å bygge skjell og eksoskjelettet. Men ferskvannsforekomster, avhengig av en rekke omstendigheter (tilstedeværelsen av visse løselige salter i jorda i reservoaret, i jorda og jorda på breddene, i vannet i innstrømmende elver og bekker) varierer sterkt både i sammensetning og i konsentrasjonen av salter oppløst i dem. Sjøvannet er mer stabilt i denne forbindelse. Nesten alle kjente elementer ble funnet i dem. Men når det gjelder viktighet, tar bordsalt først plassen, deretter magnesiumklorid og sulfat og kaliumklorid.
Ferskvannsplanter og dyr lever i et hypotont miljø, det vil si et miljø der konsentrasjonen av oppløste stoffer er lavere enn i kroppsvæsker og vev. På grunn av forskjellen i osmotisk trykk utenfor og inne i kroppen, trenger vann hele tiden inn i kroppen, og ferskvannshydrobionter blir tvunget til å fjerne det intensivt. I denne forbindelse er deres osmoreguleringsprosesser godt uttrykt. Konsentrasjonen av salter i kroppsvæskene og vevet til mange marine organismer er isotonisk med konsentrasjonen av oppløste salter i vannet rundt. Derfor er deres osmoregulatoriske funksjoner ikke utviklet i samme grad som hos ferskvannsdyr. Vanskeligheter med osmoregulering er en av grunnene til at mange marine planter og spesielt dyr ikke var i stand til å befolke ferskvannsforekomster og viste seg, med unntak av visse representanter, å være typiske marine innbyggere (coelenterata - Coelenterata, pigghuder - Echinodermata, pogonophora - Pogonophora , svamper - Spongia, kappedyr – Tunicata). På det samme Insekter lever praktisk talt ikke i hav og hav, mens ferskvannsbassenger er rikelig befolket av dem. Typisk marine og typisk ferskvannsarter tolererer ikke betydelige endringer i saltholdighet i vannet. Alle av dem er stenohaline organismer. Det er relativt få euryhaline dyr av ferskvann og marin opprinnelse. De finnes vanligvis, og i betydelige mengder, i brakkvann. Disse er ferskvannsgjedde (Stizostedion lucioperca), brasme (Abramis brama), gjedde (Esox lucius) og multefamilien (Mugilidae).
I ferskvann er planter festet på bunnen av reservoaret vanlige. Ofte er deres fotosyntetiske overflate plassert over vannet. Disse er cattails (Typha), siv (Scirpus), pilspisser (Sagittaria), vannliljer (Nymphaea), eggkapsler (Nuphar). I andre er de fotosyntetiske organene nedsenket i vann. Disse inkluderer pondweed (Potamogeton), urut (Myriophyllum) og elodea (Elodea). Noen høyere ferskvannsplanter er rotløse. De flyter enten fritt eller vokser over gjenstander under vann eller alger festet til bakken.
Mens oksygen ikke spiller en vesentlig rolle i luftmiljøet, er det den viktigste miljøfaktoren i vannmiljøet. Innholdet i vann er omvendt proporsjonalt med temperaturen. Med synkende temperatur øker oppløseligheten av oksygen, som andre gasser. Akkumuleringen av oksygen oppløst i vann skjer som et resultat av dets inntreden fra atmosfæren, så vel som på grunn av den fotosyntetiske aktiviteten til grønne planter. Når vann blandes, som er typisk for rennende reservoarer og spesielt for hurtigstrømmende elver og bekker, øker også oksygeninnholdet.
Ulike dyr har ulike behov for oksygen. For eksempel er ørret (Salmo trutta) og ørekyte (Phoxinus phoxinus) svært følsomme for mangelen og lever derfor kun i raskt rennende, kaldt og godt blandet vann. Mort (Rutilus rutilus), ruffe (Acerina cernua), karpe (Cyprinus carpio), karpe (Carassius carassius) er upretensiøse i denne forbindelse, og larvene til chironomid-mygg (Chironomidae) og tubifex-ormer (Tubifex) lever på stort dyp. hvor det ikke er oksygen i det hele tatt eller svært lite. Vanninsekter og bløtdyr (Pulmonata) kan også leve i vannmasser med lavt oksygennivå. Imidlertid stiger de systematisk til overflaten og lagrer frisk luft i noen tid.
Karbondioksid er omtrent 35 ganger mer løselig i vann enn oksygen. Det er nesten 700 ganger mer av det i vann enn i atmosfæren der det kommer fra. I tillegg er karbonater og bikarbonater av alkali- og jordalkalimetaller en kilde til karbondioksid i vann. Karbondioksid inneholdt i vann sikrer fotosyntese av vannplanter og tar del i dannelsen av kalkholdige skjelettstrukturer hos virvelløse dyr.
Konsentrasjonen av hydrogenioner (pH) er av stor betydning i livet til vannlevende organismer. Ferskvannsbassenger med en pH på 3,7–4,7 anses som sure, 6,95–7,3 anses som nøytrale, og de med en pH større enn 7,8 er alkaliske. I ferskvannsmasser opplever pH til og med daglige svingninger. Sjøvann er mer alkalisk og pH endres mye mindre enn ferskvann. pH synker med dybden.
Konsentrasjonen av hydrogenioner spiller en stor rolle i distribusjonen av vannlevende organismer. Ved en pH på mindre enn 7,5 vokser gresshoppe (Isoetes) og burberry (Sparganium); ved 7,7–8,8, dvs. i et alkalisk miljø, utvikles mange typer tjern og elodea. I det sure vannet i sumper dominerer sphagnummoser (Sphagnum), men elasmobranch bløtdyr av slekten Unio er fraværende; andre bløtdyr er sjeldne, men skjelljordstengler (Testacea) er rikelig. De fleste ferskvannsfisk tåler en pH-verdi mellom 5 og 9. Er pH-verdien mindre enn 5 er det en massiv død av fisk, og over 10 dør all fisk og andre dyr.
Økologiske grupper av hydrobioner. Vannsøylen - pelagisk (pelagos - hav) er bebodd av pelagiske organismer som aktivt kan svømme eller oppholde seg (flyte) i visse lag. I samsvar med dette er pelagiske organismer delt inn i to grupper - nekton og plankton. Bunnbeboere utgjør den tredje økologiske gruppen av organismer - benthos.
Nekton (nekios–· flytende)– Dette er en samling av pelagiske aktivt bevegelige dyr som ikke har direkte forbindelse med bunnen. Dette er hovedsakelig store dyr som kan overvinne lange avstander og sterke vannstrømmer. De er preget av en strømlinjeformet kroppsform og velutviklede bevegelsesorganer. Typiske nektoniske organismer er fisk, blekksprut, pinnipeds og hval. I ferskvann inkluderer nekton i tillegg til fisk amfibier og aktivt bevegelige insekter. Mange marine fisker kan bevege seg gjennom vannet med stor hastighet. Noen blekksprut (Oegopsida) svømmer veldig raskt, opptil 45–50 km/t, seilfisk (Istiopharidae) når hastigheter på opptil 100–10 km/t, og sverdfisk (Xiphias glabius) når hastigheter på opptil 130 km/t.
Plankton– sveve, vandrende)– Dette er et sett med pelagiske organismer som ikke har evnen til raske aktive bevegelser. Planktoniske organismer kan ikke motstå strømmer. Dette er hovedsakelig små dyr - dyreplankton og planter - planteplankton. Planktonet inkluderer med jevne mellomrom larvene til mange dyr som flyter i vannsøylen.
Planktoniske organismer er lokalisert enten på overflaten av vannet, eller på dypet, eller til og med i bunnlaget. De første danner en spesiell gruppe - neuston. Organismer, hvis kropp er plassert i vann, og en del av disse er over overflaten, kalles pleuston. Dette er sifonophorer (Siphonophora), andemat (Lemna), etc.
Planteplankton har veldig viktig i livet til vannforekomster, siden det er hovedprodusenten av organisk materiale. Det inkluderer først og fremst kiselalger (Diatomeae) og grønnalger (Chlorophyta), planteflagellater (Phytomastigina), peridineae (Peridineae) og kokkolitoforider (Coccolitophoridae). I de nordlige farvannene i verdenshavet dominerer kiselalger, og i tropiske og subtropiske farvann dominerer pansrede flagellater. I ferskvann er det i tillegg til kiselalger vanlig med grønn- og blågrønnalger (Suanophyta).
Dyreplankton og bakterier finnes på alle dyp. Marint dyreplankton er dominert av små krepsdyr (Copepoda, Amphipoda, Euphausiacea) og protozoer (Foraminifera, Radiolaria, Tintinnoidea). Mer store representanter artene er pteropoder (Pteropoda), maneter (Scyphozoa) og svømmende ctenophora (Ctenophora), salper (Salpae) og noen ormer (Alciopidae, Tomopteridae). I ferskvann er dårlig svømmende, relativt store krepsdyr (Daphnia, Cyclopoidea, Ostracoda, Simocephalus; Fig. 14), mange hjuldyr (Rotatoria) og protozoer vanlige.
Det største artsmangfoldet oppnås av plankton i tropiske farvann.
Grupper av planktoniske organismer er differensiert etter størrelse. Nannoplankton (nannos - dverg) er de minste algene og bakteriene; mikroplankton (mikro - små) - de fleste alger, protozoer, hjuldyr; mesoplankton (mesos - midten) - copepoder og cladocerans, reker og en rekke dyr og planter, ikke mer enn 1 cm i lengde; makroplankton (makroer – store) – maneter, mysider, reker og andre organismer større enn 1 cm; megaloplankton (megalos – enormt) – veldig stort, over 1 m, dyr. For eksempel når svømmektenophoren (Cestus veneris) en lengde på 1,5 m, og cyanea-maneten (Suapea) har en klokke med en diameter på opptil 2 m og tentakler 30 m lange.

Planktoniske organismer er en viktig næringskomponent for mange vannlevende dyr (inkludert giganter som bardehvaler - Mystacoceti), spesielt med tanke på at de, og spesielt planteplankton, er preget av sesongmessige utbrudd av massereproduksjon (vannoppblomstring).
Benthos– dybde)– et sett med organismer som lever på bunnen (på bakken og i bakken) av vannforekomster. Den er delt inn i phytobenthos og zoobenthos. Hovedsakelig representert av festede eller sakte bevegelige dyr, samt gravende dyr. Bare på grunt vann består den av organismer som syntetiserer organisk materiale (produsenter), forbruker (forbrukere) og ødelegger (nedbrytere) det. På store dyp, hvor lys ikke trenger inn, er phytobenthos (produsenter) fraværende.
Bunnorganismer er forskjellige i livsstil - mobile, stillesittende og immobile; ved fôringsmetode - fotosyntetisk, kjøttetende, planteetende, detrivorous; etter størrelse – makro-, meso-mikrobentos.
Havets fytobentos inkluderer hovedsakelig bakterier og alger (kiselalger, grønn, brun, rød). Langs kysten er det også blomstrende planter: Zostera, Phyllospadix, Rup-pia. Den rikeste phytobenthos er i steinete og steinete områder av bunnen. Langs kysten danner tare (Laminaria) og fucus (Fucus) noen ganger en biomasse på opptil 30 kg per 1 kvm. m. På myk jord, hvor planter ikke kan feste seg godt, utvikler phytobenthos seg hovedsakelig på steder som er beskyttet mot bølger.
Ferskvannsfytobenzos er representert av bakterier, kiselalger og grønnalger. Kystplanter er rikelig, plassert innover land fra kysten i klart definerte belter. I den første sonen vokser halvt nedsenkede planter (siv, siv, starr og stang). Den andre sonen er okkupert av nedsenkede planter med flytende blader (vannliljer, andemat, vannliljer). Den tredje sonen er dominert av nedsenkede planter - damveis, elodea, etc.
I henhold til deres livsstil kan alle vannplanter deles inn i to økologiske hovedgrupper: hydrofytter - planter som er nedsenket i vann bare med sin nedre del og vanligvis rot i bakken, og hydatofytter - planter som er helt nedsenket i vann, men noen ganger flyte på overflaten eller ha flytende blader.
Den marine zoobenthosen er dominert av foraminiferer, svamper, coelenterates, nemerteans, polychaete ormer, sipunculider, mosdyr, brachiopoder, bløtdyr, ascidianer og fisk. Bunnformer er mest tallrike på grunt vann, hvor deres totale biomasse ofte når titalls kilo per kvadratmeter. m. Med dybden synker antallet benthos kraftig og på store dyp utgjør milligram per 1 kvm. m.
I ferskvann er det mindre dyrebentos enn i hav og hav, og artssammensetningen er mer ensartet. Dette er hovedsakelig protozoer, noen svamper, cilierte og oligochaete ormer, igler, mosdyr, bløtdyr og insektlarver.
Økologisk plastisitet av akvatiske organismer. Vannorganismer har mindre økologisk plastisitet enn terrestriske, siden vann er et mer stabilt miljø og dets abiotiske faktorer gjennomgår relativt små svingninger. Marine planter og dyr er minst plastisk. De er svært følsomme for endringer i saltholdighet og temperatur i vannet. Således kan madrepore-koraller ikke tåle selv svak avsalting av vann og lever bare i havet, dessuten på fast grunn ved en temperatur ikke lavere enn 20 ° C. Dette er typiske stenobionter. Det finnes imidlertid arter med økt økologisk plastisitet. For eksempel er rhizomet Cyphoderia ampulla en typisk eurybiont. Den lever i hav og ferskvann, i varme dammer og kalde innsjøer.
Ferskvannsdyr og -planter er som regel mye mer plastiske enn marine, siden ferskvann som livsmiljø er mer varierende. De mest fleksible er brakkvannsinnbyggerne. De er tilpasset både høye konsentrasjoner av oppløste salter og betydelig avsalting. Det er imidlertid et relativt lite antall arter, siden miljøfaktorer gjennomgår betydelige endringer i brakkvann.
Bredden av økologisk plastisitet til akvatiske organismer vurderes ikke bare i forhold til hele komplekset av faktorer (eury- og stanobiontisitet), men også til enhver av dem. Kystplanter og dyr, i motsetning til innbyggerne i åpne soner, er hovedsakelig eurytermiske og euryhaline organismer, siden nær kysten er temperaturforholdene og saltregimet ganske varierende (oppvarming av solen og relativt intens avkjøling, avsalting ved tilstrømning av vann fra bekker og elver, spesielt i regntiden osv.). En typisk stenotermisk art er lotus. Den vokser bare i godt oppvarmede grunne reservoarer. Av samme grunner viser innbyggerne i overflatelagene seg å være mer eurytermiske og euryhaline sammenlignet med dyphavsformer.
Økologisk plastisitet fungerer som en viktig regulator for spredning av organismer. Som regel er vannlevende organismer med høy økologisk plastisitet ganske utbredt. Dette gjelder for eksempel elodea. Imidlertid er krepsdyret saltlake (Artemia salina) diametralt i motsetning til det i denne forstand. Den lever i små vannmasser med veldig salt vann. Dette er en typisk stenohaline-representant med smal økologisk plastisitet. Men i forhold til andre faktorer er den veldig plastisk og finnes derfor overalt i saltvannsforekomster.
Økologisk plastisitet avhenger av organismens alder og utviklingsfase. Dermed den marine gastropoden Littorina i voksen alder daglig ved lavvann lang tid gjør uten vann, og larvene fører en rent planktonisk livsstil og tåler ikke uttørking.
Tilpasningsegenskaper til vannplanter.Økologien til vannplanter, som nevnt, er veldig spesifikk og skiller seg kraftig fra økologien til de fleste landlevende planteorganismer. Vannplanters evne til å absorbere fuktighet og mineralsalter direkte fra miljø gjenspeiles i deres morfologiske og fysiologiske organisering. Vannplanter er først og fremst preget av dårlig utvikling av ledende vev og rotsystemer. Sistnevnte tjener hovedsakelig for festing til undervannssubstratet og, i motsetning til landplanter, utfører ikke funksjonen mineralernæring og vannforsyning. I denne forbindelse er røttene til rotfestede vannplanter blottet for rothår. De lever av hele overflaten av kroppen. De kraftig utviklede rhizomene til noen av dem tjener til vegetativ forplantning og lagring av næringsstoffer. Dette er mange tjerner, vannliljer og eggekapsler.
Den høye tettheten av vann gjør det mulig for planter å bo i hele tykkelsen. For dette formålet har lavere planter som bor i forskjellige lag og fører en flytende livsstil spesielle vedheng som øker oppdriften og lar dem forbli suspendert. Mekanisk vev er dårlig utviklet hos høyere hydrofytter. I bladene, stilkene og røttene deres, som nevnt, er det luftbærende intercellulære hulrom. Dette øker lettheten og oppdriften til organer som er suspendert i vann og flyter på overflaten, og hjelper også til å vaske bort indre celler med vann med gasser og salter oppløst i det. Hydatofytter er generelt preget av en stor bladoverflate med et lite totalt plantevolum. Dette gir dem intens gassutveksling når det er mangel på oksygen og andre gasser oppløst i vann. Mange tjerntang (Potamogeton lusens, P. perfoliatus) har tynne og veldig lange stilker og blader, dekslene deres er lett gjennomtrengelige for oksygen. Andre planter har sterkt dissekerte blader (vannsmørblomst – Ranunculus aquatilis, urut – Myriophyllum spicatum, hornurt – Ceratophyllum dernersum).
En rekke vannplanter har utviklet seg heterophyly (ulike blader). For eksempel, i Salvinia, tjener nedsenkede blader som mineralnæring, mens flytende blader fungerer som organisk næring. I vannliljer og eggekapsler er de flytende og nedsenkede bladene betydelig forskjellige fra hverandre. Den øvre overflaten av de flytende bladene er tett og læraktig med et stort antall stomata. Dette fremmer bedre gassutveksling med luft. Det er ingen stomata på undersiden av flytende eller nedsenkede blader.
Et like viktig adaptivt trekk ved planter for å leve i et vannmiljø er at bladene nedsenket i vann vanligvis er veldig tynne. Klorofyll i dem er ofte lokalisert i cellene i epidermis. Dette fører til en økning i fotosyntesehastigheten under dårlige lysforhold. Slike anatomiske og morfologiske trekk kommer tydeligst til uttrykk i mange dammene (Potamogeton), elodea (Helodea canadensis), vannmoser (Riccia, Fontinalis) og Vallisneria spiralis.
Beskyttelsen av vannplanter mot utvasking av mineralsalter fra celler er sekresjon av slim fra spesielle celler og dannelse av endoderm i form av en ring av tykkere vegger.
Den relativt lave temperaturen i vannmiljøet forårsaker døden av vegetative deler av planter nedsenket i vann etter dannelsen av vinterknopper, samt erstatning av sommermøre. tynne blader tøffere og kortere vinter. Samtidig påvirker lav vanntemperatur de generative organene til vannplanter negativt, og dens høye tetthet gjør pollenoverføring vanskelig. Derfor formerer vannplanter seg intensivt med vegetative midler. Den seksuelle prosessen er undertrykt hos mange av dem. Ved å tilpasse seg egenskapene til vannmiljøet, bærer de fleste nedsenkede og flytende planter blomstrende stengler opp i luften og formerer seg seksuelt (pollen bæres av vind- og overflatestrømmer). De resulterende fruktene, frøene og andre rudimenter fordeles også av overflatestrømmer (hydrokory).
Vannholdige planter inkluderer ikke bare vannplanter, men også mange kystplanter. Fruktene deres er svært flytende og kan forbli i vann i lang tid uten å miste spiringen. Vann transporterer fruktene og frøene til chastukha (Alisma plantago-aquatica), pilspiss (Sagittaria sagittifolia), sageweed (Butomusumbellatus), damweed og andre planter. Fruktene til mange sarg (Sagekh) er innelukket i særegne luftsekker og bæres også av vannstrømmer. Det antas at til og med kokospalmer spredte seg over skjærgårdene til de tropiske øyene i Stillehavet takket være oppdriften til fruktene deres - kokosnøtter. Langs Vakhsh-elven, langs kanalene, spredte gumai-ugressen (Sorgnum halepense) seg på samme måte.
Tilpasningsegenskaper til vannlevende dyr. Dyrenes tilpasning til vannmiljøet er enda mer mangfoldig enn planters. De har anatomiske, morfologiske, fysiologiske, atferdsmessige og andre adaptive egenskaper. Selv bare å liste dem er vanskelig. Derfor vil vi i generelle termer bare nevne de mest karakteristiske av dem.
Dyr som lever i vannsøylen har for det første tilpasninger som øker oppdriften og lar dem motstå bevegelse av vann og strømmer. Bunnorganismer utvikler tvert imot tilpasninger som hindrer dem i å stige opp i vannsøylen, det vil si at de reduserer oppdriften og lar dem holde seg på bunnen selv i raskt rennende vann.
I små former som lever i vannsøylen, observeres en reduksjon i skjelettformasjoner. Hos protozoer (Rhizopoda, Radiolaria) er skjellene porøse, og flintryggene på skjelettet er hule innvendig. Den spesifikke tettheten til maneter (Scyphozoa) og ctenophora (Ctenophora) reduseres på grunn av tilstedeværelsen av vann i vevet. En økning i oppdrift oppnås også ved akkumulering av fettdråper i kroppen (nattlys - Noctiluca, radiolarians - Radiolaria). Større ansamlinger av fett er også observert i noen krepsdyr (Cladocera, Copepoda), fisk og hvaler. Den spesifikke tettheten til kroppen reduseres også av gassbobler i protoplasmaet til testate amøber og luftkamre i skjellene til bløtdyr. Mange fisker har svømmeblærer fylt med gass. Sifonoforene Physalia og Velella utvikler kraftige lufthulrom.
Dyr som passivt svømmer i vannsøylen er ikke bare preget av en reduksjon i vekt, men også av en økning i det spesifikke overflatearealet av kroppen. Faktum er at jo større viskositeten til mediet og jo høyere det spesifikke overflatearealet til kroppen er, jo saktere synker det ned i vann. Som et resultat blir dyrets kropp flatet og alle slags ryggrader, utvekster og vedheng dannes på den. Dette er karakteristisk for mange radiolarier (Chalengeridae, Aulacantha), flagellater (Leptodiscus, Craspedotella) og foraminifera (Globigerina, Orbulina). Siden viskositeten til vannet avtar med økende temperatur og øker med økende saltholdighet, er tilpasninger til økt friksjon mest uttalt ved høye temperaturer og lave saltholdigheter. For eksempel er det flagellerte Ceratium fra Det indiske hav bevæpnet med lengre, hornlignende vedheng enn de som finnes i det kalde vannet i det østlige Atlanterhavet.
Aktiv svømming hos dyr utføres ved hjelp av flimmerhår, flageller og kroppsbøyning. Dette er hvordan protozoer, cilierte ormer og hjuldyr beveger seg.
Blant vannlevende dyr er reaktiv svømming vanlig på grunn av energien til den utstøpte vannstrømmen. Dette er typisk for protozoer, maneter, øyenstikkerlarver og noen muslinger. Den reaktive bevegelsesmåten når sin høyeste perfeksjon hos blekksprut. Noen blekksprut utvikler en hastighet på 40–50 km/t når de kaster ut vann. Større dyr utvikler spesialiserte lemmer (svømmebein hos insekter, krepsdyr; finner, svømmeføtter). Kroppen til slike dyr er dekket med slim og har en strømlinjeformet form.
En stor gruppe dyr, for det meste ferskvann, bruker en overflatefilm av vann (overflatespenning) for å bevege seg rundt. For eksempel løper spinnbiller (Gyrinidae) og vannstrider (Gerridae, Veliidae) fritt på den. Små Hydrophilidae biller beveger seg langs den nedre overflaten av filmen, og damsnegler (Limnaea) og mygglarver er suspendert fra den. Alle av dem har en rekke funksjoner i strukturen til lemmer, og integumentene deres blir ikke fuktet av vann.
Bare i vannmiljøet finner man ubevegelige dyr som fører en tilknyttet livsstil. De er preget av en særegen kroppsform, liten oppdrift (kroppens tetthet er større enn tettheten til vann) og spesielle enheter for feste til underlaget. Noen fester seg til bakken, andre kryper langs den eller fører en gravende livsstil, noen slår seg ned på undervannsobjekter, spesielt bunnen av skip.
Av dyrene som er festet til bakken, er de mest typiske svamper, mange coelenterater, spesielt hydroider (Hydroidea) og korallpolypper (Anthozoa), crinoider (Crinoidea), muslinger (Bivalvia), havkaker (Cirripedia), etc.
Blant gravende dyr er det spesielt mange ormer, insektlarver og bløtdyr. Visse fisk (piggfisk - Cobitis taenia, flyndre - Pleuronectidae, rokker - Rajidae) og lampreylarver (Petromyzones) tilbringer betydelig tid i bakken. Overfloden av disse dyrene og deres artsmangfold avhenger av typen jord (stein, sand, leire, silt). Det er vanligvis færre av dem på steinete jord enn på gjørmete jord. Virvelløse dyr, som koloniserer gjørmete jord i store mengder, skaper optimale forhold for livet til en rekke større bunndyrrovdyr.
De fleste vannlevende dyr er poikiloterme, og kroppstemperaturen deres avhenger av temperaturen i miljøet. Hos homeotermiske pattedyr (pinnipeds, hvaler) dannes et tykt lag av subkutant fett, som utfører en termisk isolasjonsfunksjon.
For vannlevende dyr er miljøbelastningen viktig. I denne forbindelse er det stenobathiske dyr, som ikke tåler store trykksvingninger, og eurybatiske dyr, som lever både ved høyt og lavt trykk. Holothurianere (Elpidia, Myriotrochus) lever på dybder fra 100 til 9000 m, og mange arter av Storthyngura-kreps, pogonophora, crinoider er lokalisert på dybder fra 3000 til 10 000 m. Slike dyphavsdyr har spesifikke organisatoriske trekk: en økning i kroppsstørrelse ; forsvinning eller dårlig utvikling av det kalkholdige skjelettet; ofte – reduksjon av synsorganene; styrking av utviklingen av taktile reseptorer; mangel på kroppspigmentering eller omvendt mørk farge.
Opprettholdelse av et visst osmotisk trykk og ionisk tilstand av løsninger i dyrekroppen er sikret av komplekse mekanismer for vann-saltmetabolisme. Imidlertid er de fleste vannlevende organismer poikilosmotiske, det vil si at det osmotiske trykket i kroppen deres avhenger av konsentrasjonen av oppløste salter i vannet rundt. Bare virveldyr, høyere krepsdyr, insekter og deres larver er homoiosmotiske - de opprettholder konstant osmotisk trykk i kroppen, uavhengig av saltholdigheten i vannet.
Marine virvelløse dyr har generelt ikke mekanismer for vann-saltmetabolisme: anatomisk er de lukket for vann, men osmotisk er de åpne. Det vil imidlertid være feil å si at de absolutt ikke har noen mekanismer som kontrollerer vann-saltmetabolismen.

De er rett og slett ufullkomne, og dette forklares med at saltholdigheten i sjøvann er nær saltholdigheten til kroppsjuicene. Tross alt, i ferskvannshydrobioner er saltholdigheten og iontilstanden til mineralstoffer i kroppsjuicene som regel høyere enn i det omkringliggende vannet. Derfor er deres osmoregulatoriske mekanismer godt uttrykt. Den vanligste måten å opprettholde konstant osmotisk trykk på er å regelmessig fjerne vann som kommer inn i kroppen ved hjelp av pulserende vakuoler og utskillelsesorganer. Hos andre dyr utvikles ugjennomtrengelige dekker av kitin eller kåte formasjoner for disse formålene. Noen mennesker produserer slim på overflaten av kroppen.
Vanskeligheten med å regulere osmotisk trykk i ferskvannsorganismer forklarer deres artsfattigdom sammenlignet med sjøinnbyggere.
La oss bruke eksemplet med fisk for å se hvordan osmoregulering av dyr skjer i sjø og ferskvann. Ferskvannsfisk fjerner overflødig vann gjennom det intensive arbeidet med ekskresjonssystemet, og absorberer salter gjennom gjellefilamentene. Marin fisk, tvert imot, blir tvunget til å fylle på vannreservene sine og drikker derfor sjøvann, og overskuddssaltene som følger med fjernes fra kroppen gjennom gjellefilamentene (fig. 15).
Endre forhold i vannmiljøet forårsaker visse atferdsreaksjoner hos organismer. Vertikale migrasjoner av dyr er assosiert med endringer i belysning, temperatur, saltholdighet, gassregime og andre faktorer. I hav og hav deltar millioner av tonn vannlevende organismer i slike migrasjoner (senker ned i dypet, stiger til overflaten). Under horisontale migrasjoner kan vannlevende dyr reise hundrevis og tusenvis av kilometer. Dette er gyte-, overvintrings- og fôringsvandringene til mange fisker og akvatiske pattedyr.
Biofiltre og deres økologiske rolle. En av de spesifikke egenskapene til vannmiljøet er tilstedeværelsen i det av et stort antall små partikler av organisk materiale - detritus, dannet av døende planter og dyr. Enorme masser av disse partiklene legger seg på bakterier og, takket være gassen som frigjøres som følge av bakterieprosessen, blir de konstant suspendert i vannsøylen.
Detritus er en mat av høy kvalitet for mange vannlevende organismer, så noen av dem, de såkalte biofiltrene, har tilpasset seg for å få det ved hjelp av spesifikke mikroporøse strukturer. Disse strukturene filtrerer så å si vannet, og holder på partikler suspendert i det. Denne fôringsmetoden kalles filtrering. En annen gruppe dyr avleirer detritus på overflaten av enten sin egen kropp eller på spesielle fangstinnretninger. Denne metoden kalles sedimentering. Ofte lever den samme organismen ved både filtrering og sedimentering.
Biofiltreringsdyr (elasmobranch bløtdyr, fastsittende pigghuder og polychaete annelids, mosdyr, ascidianer, planktoniske krepsdyr og mange andre) spiller en stor rolle i den biologiske rensingen av vannforekomster. For eksempel, en koloni med blåskjell (Mytilus) per 1 kvm. m passerer gjennom mantelhulen opp til 250 kubikkmeter. m vann per dag, filtrering og utfelling av suspenderte partikler. Det nesten mikroskopiske krepsdyret Calanus (Calanoida) renser opptil 1,5 liter vann per dag. Hvis vi tar i betraktning det enorme antallet av disse krepsdyrene, virker arbeidet de gjør med biologisk rensing av vannforekomster virkelig enormt.
I ferskvann er aktive biofiltre perlebygg (Unioninae), tannløse blåskjell (Anodontinae), sebramuslinger (Dreissena), dafnia (Daphnia) og andre virvelløse dyr. Deres betydning som et slags biologisk "rensesystem" av vannforekomster er så stor at det er nesten umulig å overvurdere det.
Soneinndeling av vannmiljøet. Det akvatiske bomiljøet er preget av klart definert horisontal og spesielt vertikal sonering. Alle hydrobionter er strengt begrenset til å leve i visse soner som er forskjellige i forskjellige livsbetingelser.
I verdenshavet kalles vannsøylen pelagisk, og bunnen er bunndyr. Følgelig skilles også økologiske grupper av organismer som lever i vannsøylen (pelagisk) og på bunnen (bunnfisk).
Bunnen, avhengig av dybden av dens forekomst fra vannoverflaten, er delt inn i sublitoral (et område med gradvis nedgang til en dybde på 200 m), bathyal (bratt skråning), abyssal (havbunn med en gjennomsnittlig dybde på 3–6 km), ultra-abyssal (bunnen av oseaniske depresjoner som ligger på en dybde på 6 til 10 km). Litoralsonen utmerker seg også - kanten av kysten, som periodisk oversvømmes under høyvann (fig. 16).
Det åpne vannet i verdenshavet (pelagialt) er også delt inn i vertikale soner som tilsvarer de bentiske sonene: epipelagisk, bathypelagisk, abyssopelagisk.
De littorale og sublitorale sonene er rikest befolket av planter og dyr. Det er mye sollys, lavtrykk og betydelige temperatursvingninger. Innbyggerne i det avgrunns- og ultra-avgrunnsdypet lever ved en konstant temperatur, i mørket, og opplever et enormt trykk, og når flere hundre atmosfærer i havdypet.

En lignende, men mindre klart definert sonering er også karakteristisk for ferskvannsforekomster i innlandet.
| Parameternavn | Betydning |
| Artikkel emne: | Vannmiljø. |
| Rubrikk (tematisk kategori) | Økologi |
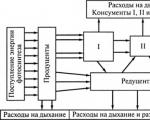
Vann er livets første medium: liv oppsto i det og de fleste organismegruppene ble dannet. Alle innbyggere i vannmiljøet kalles hydrobionter. Et karakteristisk trekk ved vannmiljøer er bevegelsen av vann, som manifesterer seg i formen strømmer(vannoverføring i en retning) og uro(avvik av vannpartikler fra deres opprinnelige posisjon og påfølgende retur til den). Golfstrømmen fører 2,5 millioner m^3 vann per år, som er 25 ganger mer enn alle elvene på jorden til sammen. I tillegg oppstår tidevannssvingninger i havnivået under påvirkning av tiltrekningen av Månen og Solen.
I tillegg til bevegelsen av vann til nummeret viktige egenskaper av vannmiljøet inkluderer tetthet og viskositet, spektralitet, oppløst oksygen og mineralinnhold.
Tetthet og viskositet bestemme først og fremst betingelsene for bevegelse av vannlevende organismer. Jo høyere tetthet av vannet er, jo mer støttende blir det, jo lettere er det å oppholde seg i det. En annen betydning av tetthet er dens press på kroppen. Med en utdyping på 10,3 m i ferskvann og 9,986 m i sjøvann øker trykket med 1 atm. Når viskositeten øker, øker motstanden mot aktiv bevegelse av organismer. Tettheten av levende vev er høyere enn tettheten til ferskvann og sjøvann; derfor har vannlevende organismer i utviklingsprosessen utviklet forskjellige strukturer som øker oppdriften deres - en generell økning i den relative overflaten av kroppen på grunn av en reduksjon i størrelse; utflating; utvikling av ulike utvekster (bust); reduksjon i kroppstetthet på grunn av skjelettreduksjon; opphopning av fett og tilstedeværelsen av en svømmeblære. Vann, i motsetning til luft, har en større oppdriftskraft, og derfor er den maksimale størrelsen på vannlevende organismer mindre begrenset.
Termiske egenskaper vann skiller seg betydelig fra de termiske egenskapene til luft. Den høye spesifikke varmekapasiteten til vann (500 ganger høyere) og termisk ledningsevne (30 ganger høyere) bestemmer en konstant og relativt jevn temperaturfordeling i vannmiljøet. Temperatursvingninger i vann er ikke like skarpe som i luft. Temperatur påvirker hastigheten til ulike prosesser.
Lys- og lysmodus. Solen lyser opp overflatene på land og hav med samme intensitet, men vannets evne til å absorbere og spre seg er ganske høy, noe som begrenser dybden av lysinntrengning i havet. Dessuten absorberes ikke stråler med forskjellige bølgelengder likt: røde blir spredt nesten umiddelbart, mens blå og grønne går dypere. Sonen der fotosyntesehastigheten overstiger respirasjonshastigheten kalles vanligvis eufotiske sone. Den nedre grensen ved hvilken fotosyntesen balanseres ved respirasjon kalles vanligvis kompensasjonspunkt.
Åpenhet vann avhenger av innholdet av suspenderte partikler i det. Gjennomsiktighet er preget av den maksimale dybden der en spesielt senket hvit skive med en diameter på 30 cm fortsatt er synlig. klart vann i Sargassohavet (skiven er synlig på en dybde på 66 m), i Stillehavet (60 m), indiske hav(50 m). I grunt hav er gjennomsiktigheten 2-15 m, i elver 1-1,5 m.
Oksygen- nødvendig for å puste. I vann er fordelingen av oppløst oksygen utsatt for kraftige svingninger. Om natten er oksygeninnholdet i vann mindre. Respirasjon av vannlevende organismer skjer enten gjennom overflaten av kroppen eller gjennom spesielle organer (lunger, gjeller, luftrør).
Mineraler. Sjøvann inneholder hovedsakelig natrium, magnesium, klor og sulfationer. Friske kalsiumioner og karbonationer.
Miljøklassifisering vannlevende organismer. Mer enn 150 tusen arter av dyr og rundt 10 tusen arter av planter lever i vannet. De viktigste biotopene til vannlevende organismer er: vannsøylen ( pelagisk) og bunnen av reservoarene ( benthal). Det er pelagiske og bunnlevende organismer. Den pelagiske sonen er delt inn i grupper: plankton(et sett med organismer som ikke er i stand til aktiv bevegelse og beveger seg med vannstrømmer) og nekton(store dyr hvis motoriske aktivitet er tilstrekkelig til å overvinne vannstrømmer). Benthos- et sett med organismer som bor på bunnen.
Vannmiljø. - konsept og typer. Klassifisering og funksjoner i kategorien "Aquatic environment." 2017, 2018.
Habitat, forhold og levesett Praktisk anvendelse av paleontologi i geologi § I stratigrafi (bruksgrunnlaget er loven om evolusjonens irreversibilitet). § I paleogeografi er trofiske eller matforbindelser (gresk trofe - mat, ernæring) de viktigste i... .
Et trådløst miljø betyr ikke et helt trådløst nettverk. Vanligvis kommuniserer trådløse komponenter med et nettverk som bruker kabel som overføringsmedium. Slike nettverk kalles hybrid. Følgende typer trådløse nettverk finnes: LAN,...
-
Et økologisk system (økosystem) er et romlig definert sett som består av et fellesskap av levende organismer (biocenose), deres habitat (biotop) og et system av forbindelser som utveksler materie og energi mellom dem. Det er akvatiske og terrestriske naturlige... .
Du kjenner allerede begreper som "habitat" og "bomiljø". Du må lære å skille dem. Hva er "bomiljø"?
Bomiljøet er en del av naturen med et spesielt sett av faktorer, for å eksistere der ulike systematiske grupper av organismer har utviklet lignende tilpasninger.
Det er fire hovedmiljøer for liv på jorden: akvatiske, bakke-luft, jord og levende organismer.
Vannmiljø
Det akvatiske levemiljøet er preget av høy tetthet, spesiell temperatur, lys, gass og saltregimer. Organismer som lever i vannmiljøer kalles hydrobionter(fra gresk hydro- vann, bios- livet).
Temperaturregime i vannmiljøet
I vann endres temperaturen mindre enn på land på grunn av den høye spesifikke varmekapasiteten og varmeledningsevnen til vannet. En økning i lufttemperaturen på 10 °C fører til en økning i vanntemperaturen på 1 °C. Med dybden synker temperaturen gradvis. På store dyp er temperaturregimet relativt konstant (ikke høyere enn +4 °C). I de øvre lagene observeres daglige og sesongmessige svingninger (fra 0 til +36 °C). Siden temperaturen i vannmiljøet varierer innenfor et smalt område, krever de fleste vannlevende organismer en stabil temperatur. Selv små temperaturavvik forårsaket for eksempel av virksomheter som slipper ut varmt vann er skadelig for dem. Avløpsvann. Hydrobionter som kan eksistere under store temperatursvingninger finnes bare i små vannmasser. På grunn av det lille volumet av vann i disse reservoarene, observeres betydelige daglige og sesongmessige temperaturendringer.
Lysregime i vannmiljøet
Det er mindre lys i vann enn i luft. Noen av solens stråler reflekteres fra overflaten, og noen absorberes av vannsøylen.
En dag under vann er kortere enn en dag på land. Om sommeren, på en dybde på 30 m, er det 5 timer, og på en dybde på 40 m - 15 minutter. Den raske reduksjonen av lys med dybden er assosiert med dets absorpsjon av vann.
Grensen for fotosyntesesonen i havet er på en dybde på ca 200 m. I elver varierer den fra 1,0 til 1,5 m og avhenger av vannets gjennomsiktighet. Vannets klarhet i elver og innsjøer er sterkt redusert på grunn av forurensning fra suspenderte partikler. På en dybde på mer enn 1500 m er det praktisk talt ikke noe lys.
Gassregimet i vannmiljøet
I vannmiljøet er oksygeninnholdet 20-30 ganger mindre enn i luft, så det er en begrensende faktor. Oksygen kommer inn i vann på grunn av fotosyntese av vannplanter og luftoksygens evne til å løse seg opp i vann. Når vann røres, øker oksygeninnholdet i det. De øvre vannlagene er rikere på oksygen enn de nedre lagene. Ved oksygenmangel oppstår døden (massedød av vannlevende organismer). Vinterfrys oppstår når vannmasser er dekket med is. Sommer - når oppløseligheten av oksygen avtar på grunn av høy vanntemperatur. Årsaken kan også være en økning i konsentrasjonen av giftige gasser (metan, hydrogensulfid) som dannes ved nedbrytning av døde organismer uten tilgang på oksygen. På grunn av variasjonen i oksygenkonsentrasjonen er de fleste vannlevende organismer eurybionter i forhold til den. Men det finnes også stenobionter (ørret-, planaria-, maiflue- og caddisfluelarver) som ikke tåler oksygenmangel. De er indikatorer på vannrenhet. Karbondioksid oppløses i vann 35 ganger bedre enn oksygen, og konsentrasjonen i det er 700 ganger høyere enn i luft. CO2 akkumuleres i vann på grunn av respirasjon av vannlevende organismer og nedbryting av organiske rester. Karbondioksid gir fotosyntese og brukes i dannelsen av kalkholdige skjeletter av virvelløse dyr.
Saltregimet i vannmiljøet
Vannets saltholdighet spiller en viktig rolle i livet til vannlevende organismer. Basert på saltinnhold er naturlig vann delt inn i grupper presentert i tabellen:
I verdenshavet er saltholdigheten i gjennomsnitt 35 g/l. Høyest saltinnhold er i saltsjøer (opptil 370 g/l). Typiske innbyggere i ferskvann og saltvann er stenobionter. De tåler ikke fluktuasjoner i saltholdighet i vannet. Det er relativt få eurybionts (brasmer, gjedde, gjedde, ål, pinnebak, laks osv.). De kan leve i både ferskvann og saltvann.
Tilpasninger av planter til livet i vann
Alle planter i vannmiljøet kalles hydrofytter(fra gresk hydro- vann, phyton- anlegg). Bare alger lever i saltvann. Kroppen deres er ikke delt inn i vev og organer. Algene tilpasset seg endringer i sammensetningen av solspekteret avhengig av dybden ved å endre sammensetningen av pigmentene deres. Når man beveger seg fra de øvre vannlagene til de dype, endres fargen på algene i sekvensen: grønn - brun - rød (de dypeste algene).
Grønnalger inneholder grønne, oransje og gule pigmenter. De er i stand til fotosyntese under tilstrekkelig høy intensitet sollys. Derfor lever grønnalger i små ferskvannsforekomster eller i grunt hav. Disse inkluderer: spirogyra, ulotrix, ulva, etc. Brunalger inneholder i tillegg til grønne brune og gule pigmenter. De er i stand til å fange mindre intens solstråling på en dybde på 40-100 m. Representanter for brunalger er fucus og tare, som bare lever i havet. Rødalger (porfyr, phyllophora) kan leve på dyp på mer enn 200 m. I tillegg til grønne har de røde og blå pigmenter som kan fange selv svakt lys på store dyp.
I ferskvann er mekanisk vev dårlig utviklet i stilkene til høyere planter. Hvis du for eksempel fjerner en hvit vannlilje eller en gul vannlilje fra vannet, henger stilkene deres og er ikke i stand til å støtte blomstene i oppreist stilling. De er avhengige av vann på grunn av dens høye tetthet. En tilpasning til mangel på oksygen i vann er tilstedeværelsen av aerenchyma (luftbærende vev) i planteorganer. Mineraler finnes i vann, så det ledende og rotsystemet er dårlig utviklet. Røtter kan være helt fraværende (andmat, elodea, dammat) eller tjene til å forankre dem i underlaget (cattail, pilspiss, chastukha). Det er ingen rothår på røttene. Bladene er ofte tynne og lange eller kraftig dissekert. Mesofyll er ikke differensiert. Stomata til flytende blader er på oversiden, mens de til blader nedsenket i vann er fraværende. Noen planter kjennetegnes ved å ha blader av ulik form (heterofilt) avhengig av hvor de finnes. Vannliljer og pilspisser har forskjellige bladformer i vann og i luft.
Pollen, frukt og frø fra vannplanter er tilpasset spredning med vann. De har korkutvekster eller sterke skjell som hindrer vann i å komme inn og råtne.
Tilpasninger av dyr til livet i vann
I vannmiljøet er dyreverdenen rikere enn planteverdenen. Takket være deres uavhengighet fra sollys, befolket dyrene hele tykkelsen av vannet. Basert på typen morfologiske og atferdsmessige tilpasninger er de delt inn i følgende økologiske grupper: plankton, nekton, benthos.
Plankton(fra gresk planktos- svevende, vandrende) - organismer som lever i vannsøylen og beveger seg under påvirkning av strømmen. Dette er små krepsdyr, coelenterater og larvene til noen virvelløse dyr. Alle deres tilpasninger er rettet mot å øke oppdriften til kroppen:
- økning i kroppsoverflate på grunn av utflating og forlengelse av formen, utvikling av utvekster og bust;
- reduksjon i kroppstetthet på grunn av reduksjon av skjelettet, tilstedeværelsen av fettdråper, luftbobler og slimhinner.
Nekton(fra gresk nektos- flytende) - organismer som lever i vannsøylen og fører en aktiv livsstil. Representanter for nekton er fisk, hvaler, pinnipeds og blekksprut. De er i stand til å motstå strømmen ved å tilpasse seg aktiv svømming og redusere kroppsfriksjonen. Aktiv svømming oppnås gjennom velutviklede muskler. I dette tilfellet kan energien til den avgitte vannstrømmen, kroppens bøyning, finner, svømmeføtter etc. brukes Tilpasning bidrar til å redusere kroppsfriksjonen: strømlinjeformet kroppsform, elastisitet i huden, tilstedeværelse av
hudskjell og slim.
Benthos(fra gresk bunndyr- dybde) - organismer som lever på bunnen av et reservoar eller i tykkelsen av bunnjorda.
Tilpasninger av bunnlevende organismer er rettet mot å redusere oppdrift:
- vekting av kroppen på grunn av skjell (bløtdyr), kitiniserte integumenter (kreps, krabber, hummer, hummer);
- fiksering på bunnen ved hjelp av fikseringsorganer (sugekopper i igler, kroker i larver) eller en avflatet kropp (rokker, flyndre). Noen representanter graver seg ned i bakken (polychaete ormer).
I innsjøer og dammer identifiseres en annen økologisk gruppe organismer - neuston. Neuston- organismer knyttet til overflatefilmen av vann og som lever permanent eller midlertidig på denne filmen eller opptil 5 cm i dybden fra overflaten. Kroppen deres blir ikke fuktet fordi dens tetthet er mindre enn vann. Spesialdesignede lemmer lar dem bevege seg langs overflaten av vannet uten å stupe (vannstrider, spinnende biller). En unik gruppe vannlevende organismer er også perifyton— organismer som danner en begroingsfilm på undervannsobjekter. Representanter for periphyton er: alger, bakterier, protister, krepsdyr, muslinger, oligochaete ormer, bryozoaner, svamper.
Det er fire hovedlivsmiljøer på planeten Jorden: akvatiske, land-luft, jord og levende organismer. I et vannmiljø er oksygen den begrensende faktoren. Basert på arten av deres tilpasninger, er akvatiske innbyggere delt inn i økologiske grupper: plankton, nekton og benthos.
Tetthet av vann- dette er en faktor som bestemmer forholdene for bevegelse av vannlevende organismer og trykk på forskjellige dyp. For destillert vann er tettheten 1 g/cm 3 ved 4 °C. Tettheten av naturlig vann som inneholder oppløste salter kan være større, opptil 1,35 g/cm 3 . Trykket øker med dybden med et gjennomsnitt på 1 × 10 5 Pa (1 atm) for hver 10 m.
På grunn av den skarpe trykkgradienten i vannforekomster er vannlevende organismer generelt mye mer eurybatiske sammenlignet med landorganismer. Noen arter, fordelt på forskjellige dyp, tåler trykk fra flere til hundrevis av atmosfærer. For eksempel lever holothurians av slekten Elpidia og ormer Priapulus caudatus fra kystsonen til ultra-abyssal sonen. Selv ferskvannsinnbyggere, som tøfler, suvoikaer, svømmebiller, etc., tåler opptil 6 × 10 7 Pa (600 atm) i eksperimenter.
Imidlertid er mange innbyggere i hav og hav relativt stenobatiske og begrenset til visse dyp. Stenobacy er oftest karakteristisk for grunne- og dyphavsarter. Bare kystsonen er bebodd av annelidene Arenicola og limpet bløtdyr (Patella). Mange fisker, for eksempel fra gruppen av sportsfiskere, blekksprut, krepsdyr, pogonophora, sjøstjerner, etc., finnes bare på store dyp ved et trykk på minst 4 10 7 - 5 10 7 Pa (400-500 atm).
Vannets tetthet gir muligheten til å lene seg på det, noe som er spesielt viktig for ikke-skjelettformer. Miljøtettheten fungerer som en betingelse for å flyte i vann, og mange vannlevende organismer er tilpasset spesifikt til denne livsstilen. Suspenderte organismer som flyter i vann kombineres til en spesiell økologisk gruppe av vannlevende organismer - plankton ("planktos" - svevende).
Ris. 39. Økning i den relative kroppsoverflaten til planktoniske organismer (ifølge S. A. Zernov, 1949):
A - stavformet:
1 - kiselalger Synedra;
2 - cyanobacterium Aphanizomenon;
3 - peridinalge Amphisolenia;
4 - Euglena acus;
5 - blekksprut Doratopsis vermicularis;
6 - copepod Setella;
7 - Porcellana larve (Decapoda)

B - dissekerte former:
1 - bløtdyr Glaucus atlanticus;
2 - orm Tomopetris euchaeta;
3 - Palinurus krepselarve;
4 - larve av breiflabbfisk Lophius;
5 - copepod Calocalanus pavo
Plankton inkluderer encellede og koloniale alger, protozoer, maneter, sifonoforer, ctenophorer, pteropoder og kjølfotbløtdyr, ulike små krepsdyr, larver av bunndyr, fiskeegg og yngel og mange andre (fig. 39). Planktoniske organismer har mange lignende tilpasninger som øker oppdriften og hindrer dem i å synke til bunnen. Slike tilpasninger inkluderer: 1) en generell økning i den relative overflaten av kroppen på grunn av reduksjon i størrelse, utflating, forlengelse, utvikling av tallrike fremspring eller bust, som øker friksjonen med vann; 2) en reduksjon i tetthet på grunn av reduksjon av skjelettet, akkumulering av fett, gassbobler osv. I kiselalger avsettes reservestoffer ikke i form av tung stivelse, men i form av fettdråper . Nattlyset Noctiluca utmerker seg ved en slik overflod av gassvakuoler og fettdråper i cellen at cytoplasmaet i den ser ut som tråder som bare smelter sammen rundt kjernen. Sifonoforer, en rekke maneter, planktoniske gastropoder osv. har også luftkammer.
Tang (fytoplankton) De flyter passivt i vann, men de fleste planktondyr er i stand til å svømme aktivt, men i begrenset grad. Planktoniske organismer kan ikke overvinne strømmer og transporteres av dem over lange avstander. Mange typer dyreplankton Imidlertid er de i stand til vertikale migrasjoner i vannsøylen i titalls og hundrevis av meter, både på grunn av aktiv bevegelse og ved å regulere oppdriften til kroppen. En spesiell type plankton er en økologisk gruppe Neuston ("nein" - svømme) - innbyggere i overflatefilmen av vann ved grensen til luften.
Vannets tetthet og viskositet påvirker i stor grad muligheten for aktiv svømming. Dyr som er i stand til å svømme raskt og overvinne strømmens kraft, er forent i en økologisk gruppe nekton ("nektos" - flytende). Representanter for nekton er fisk, blekksprut og delfiner. Rask bevegelse i vannsøylen er bare mulig hvis du har en strømlinjeformet kroppsform og høyt utviklede muskler. Den torpedoformede formen er utviklet hos alle gode svømmere, uavhengig av deres systematiske tilhørighet og bevegelsesmetode i vannet: reaktiv, på grunn av bøyning av kroppen, ved hjelp av lemmer.
Oksygen regime. I oksygenmettet vann overstiger ikke innholdet 10 ml per 1 liter, som er 21 ganger lavere enn i atmosfæren. Derfor er pusteforholdene til vannlevende organismer betydelig kompliserte. Oksygen kommer hovedsakelig inn i vann gjennom den fotosyntetiske aktiviteten til alger og diffusjon fra luften. Derfor er de øvre lagene av vannsøylen som regel rikere på denne gassen enn de nedre. Når temperaturen og saltholdigheten til vannet øker, synker konsentrasjonen av oksygen i det. I lag som er tett befolket av dyr og bakterier, kan det skapes en kraftig mangel på O 2 på grunn av økt forbruk. For eksempel, i verdenshavet er livsrike dybder fra 50 til 1000 m preget av en kraftig forringelse av lufting - den er 7-10 ganger lavere enn i overflatevann bebodd av planteplankton. Forhold nær bunnen av reservoarer kan være nær anaerobe.
Blant akvatiske innbyggere er det mange arter som tåler store svingninger i oksygeninnhold i vann, opp til nesten fullstendig fravær. (euryoxybionts - "oksy" - oksygen, "biont" - innbygger). Disse inkluderer for eksempel ferskvannsoligochaeten Tubifex tubifex og gastropoden Viviparus viviparus. Blant fisk kan karpe, suter og karpe tåle svært lav oksygenmetning i vann. Imidlertid en rekke typer stenoxybiont - de kan bare eksistere med tilstrekkelig høy oksygenmetning av vannet (regnbueørret, ørret, ørekyt, øyenvippeorm Planaria alpina, larver av maifluer, steinfluer, etc.). Mange arter er i stand til å falle inn i en inaktiv tilstand når det er mangel på oksygen - anoksybiose – og dermed oppleve en ugunstig periode.
Respirasjon av vannlevende organismer skjer enten gjennom overflaten av kroppen eller gjennom spesialiserte organer - gjeller, lunger, luftrør. I dette tilfellet kan integumentet tjene som et ekstra åndedrettsorgan. For eksempel forbruker loach-fisken i gjennomsnitt 63 % oksygen gjennom huden. Hvis gassutveksling skjer gjennom kroppens integumenter, er de veldig tynne. Pusten gjøres også lettere ved å øke overflaten. Dette oppnås under utviklingen av arter ved dannelse av forskjellige utvekster, utflating, forlengelse og en generell reduksjon i kroppsstørrelse. Noen arter, når det er mangel på oksygen, endrer aktivt størrelsen på luftveisoverflaten. Tubifex tubifex-ormer forlenger kroppen kraftig; hydra og sjøanemone - tentakler; pigghuder - ambulacrale ben. Mange fastsittende og stillesittende dyr fornyer vannet rundt seg, enten ved å skape en rettet strøm eller ved oscillerende bevegelser, noe som fremmer blandingen. Toskallede bløtdyr bruker flimmerhår som fôrer veggene i mantelhulen til dette formålet; krepsdyr - arbeidet til mage- eller thoraxbena. Igler, klokkemygglarver (blodormer) og mange oligochaeter svaier med kroppen og stikker opp av bakken.
Hos noen arter forekommer en kombinasjon av vann- og luftånding. Disse inkluderer lungefisk, siphonophores discophants, mange lunge bløtdyr, krepsdyr Gammarus lacustris, etc. Sekundære vannlevende dyr beholder vanligvis den atmosfæriske typen respirasjon da det er mer energimessig fordelaktig og derfor krever kontakt med luften, for eksempel pinnipeds, hvaler, vannbiller , mygglarver osv.
Mangel på oksygen i vann fører noen ganger til katastrofale fenomener - Jeg dør, ledsaget av døden til mange vannlevende organismer. Vinteren fryser ofte forårsaket av dannelsen av is på overflaten av vannmasser og opphør av kontakt med luft; sommer- en økning i vanntemperatur og en resulterende reduksjon i oksygenløselighet.
Hyppig død av fisk og mange virvelløse dyr om vinteren er karakteristisk for for eksempel den nedre delen av elvebassenget Ob, hvis vann, som renner fra våtmarkene i det vestsibirske lavlandet, er ekstremt fattig på oppløst oksygen. Noen ganger oppstår døden i havet.
I tillegg til mangel på oksygen, kan døden være forårsaket av en økning i konsentrasjonen av giftige gasser i vann - metan, hydrogensulfid, CO 2, etc., dannet som følge av nedbrytning av organisk materiale i bunnen av reservoarene .
Salt regime.Å opprettholde vannbalansen til vannlevende organismer har sine egne spesifikasjoner. Hvis det for landdyr og planter er viktigst å forsyne kroppen med vann under forhold med mangel, så er det for hydrobioner ikke mindre viktig å opprettholde en viss mengde vann i kroppen når det er et overskudd av det i miljøet . For mye vann i cellene fører til endringer i osmotisk trykk og forstyrrelse av de viktigste vitale funksjonene.
Mest vannlevende liv poikilosmotisk: det osmotiske trykket i kroppen deres avhenger av saltholdigheten i vannet rundt. Derfor er den viktigste måten for vannlevende organismer å opprettholde saltbalansen på å unngå habitater med uegnet saltholdighet. Ferskvannsformer kan ikke eksistere i havet, og marine former tåler ikke avsalting. Hvis saltholdigheten i vannet er gjenstand for endringer, beveger dyr seg på jakt etter et gunstig miljø. For eksempel når overflatelagene i havet avsaltes etter kraftig regn, kommer radiolarier, sjøkrepsdyr Calanus og andre ned til en dybde på 100 m. Virveldyr, høyere krepsdyr, insekter og deres larver som lever i vann tilhører bl.a. homoiosmotisk arter, opprettholde konstant osmotisk trykk i kroppen uavhengig av konsentrasjonen av salter i vannet.
Hos ferskvannsarter er kroppsjuicer hypertoniske i forhold til vannet rundt. De er i fare for overdreven vanning hvis vannstrømmen ikke forhindres eller overflødig vann ikke fjernes fra kroppen. I protozoer oppnås dette ved arbeidet med utskillelsesvakuoler, i flercellede organismer - ved å fjerne vann gjennom utskillelsessystemet. Noen ciliater skiller ut en mengde vann som tilsvarer kroppsvolumet hvert 2-2,5 minutt. Cellen bruker mye energi på å "pumpe ut" overflødig vann. Med økende saltholdighet bremses arbeidet til vakuoler. Således, i Paramecium-tøfler, ved en saltholdighet på 2,5 %o, pulserer vakuolen med intervaller på 9 s, ved 5 %o - 18 s, ved 7,5 %o - 25 s. Ved en saltkonsentrasjon på 17,5 % o slutter vakuolen å virke, siden forskjellen i osmotisk trykk mellom cellen og det ytre miljøet forsvinner.
Hvis vann er hypertonisk i forhold til kroppsvæskene til vannlevende organismer, risikerer de dehydrering som følge av osmotiske tap. Beskyttelse mot dehydrering oppnås ved å øke konsentrasjonen av salter også i kroppen til vannlevende organismer. Dehydrering forhindres av vann-ugjennomtrengelige integumenter av homoiosmotiske organismer - pattedyr, fisk, høyere kreps, vannlevende insekter og deres larver.
Mange poikilosmotiske arter går over til en inaktiv tilstand - suspendert animasjon som et resultat av mangel på vann i kroppen med økende saltholdighet. Dette er karakteristisk for arter som lever i bassenger med sjøvann og i kystsonen: hjuldyr, flagellater, ciliater, noen krepsdyr, svartehavspolychaeten Nereis divesicolor, etc. Salt suspendert animasjon- et middel for å overleve ugunstige perioder under forhold med varierende saltholdighet i vannet.
Virkelig euryhalin Det er ikke mange arter blant akvatiske innbyggere som kan leve i aktiv tilstand i både ferskvann og saltvann. Dette er hovedsakelig arter som bor i elvemunninger, elvemunninger og andre brakkvannsforekomster.
Temperatur reservoarene er mer stabile enn på land. Det henger sammen med fysiske egenskaper vann, spesielt dets høye spesifikke varmekapasitet, på grunn av hvilken mottak eller frigjøring av en betydelig mengde varme ikke forårsaker for plutselige endringer i temperaturen. Fordampningen av vann fra overflaten av reservoarene, som forbruker omtrent 2263,8 J/g, forhindrer overoppheting av de nedre lagene, og dannelsen av is, som frigjør fusjonsvarmen (333,48 J/g), bremser nedkjølingen.
Amplituden av årlige temperatursvingninger i de øvre lagene av havet er ikke mer enn 10-15 °C, i kontinentale farvann - 30-35 °C. Dype vannlag er preget av konstant temperatur. I ekvatorialfarvann gjennomsnittlig årstemperatur overflatelag +(26-27) °C, i polare lag - ca 0 °C og under. I varme landbaserte kilder kan vanntemperaturen nærme seg +100 °C, og i undervannsgeysirer, ved høyt trykk på havbunnen, er det registrert temperaturer på +380 °C.
Dermed er det en ganske betydelig variasjon av temperaturforhold i reservoarene. Mellom de øvre vannlagene med sesongmessige temperatursvingninger uttrykt i dem og de nedre, hvor det termiske regimet er konstant, er det en sone med temperaturhopp, eller termoklin. Termoklinen er mer uttalt i varme hav, hvor temperaturforskjellen mellom ytre og dype vann er større.
På grunn av det mer stabile temperaturregimet til vann er stenotermi vanlig blant vannlevende organismer i mye større grad enn blant landbefolkningen. Eurytermiske arter finnes hovedsakelig i grunne kontinentale reservoarer og i kystsonen til hav med høye og tempererte breddegrader, hvor daglige og sesongmessige temperatursvingninger er betydelige.
Lysmodus. Det er mye mindre lys i vann enn i luft. Noen av strålene som faller inn på overflaten av et reservoar reflekteres i luften. Jo lavere Solens posisjon er, desto sterkere er refleksjonen, så dagen under vann er kortere enn på land. For eksempel en sommerdag nær øya Madeira på 30 m - 5 timers dyp, og på 40 meters dyp kun 15 minutter. Den raske reduksjonen i mengden lys med dybden er assosiert med dets absorpsjon av vann. Stråler med forskjellige bølgelengder absorberes forskjellig: røde forsvinner nær overflaten, mens blågrønne trenger mye dypere. Skumringen i havet som blir dypere med dybden er først grønn, deretter blå, indigo og blåfiolett, og til slutt viker den for konstant mørke. Følgelig erstatter grønne, brune og røde alger, spesialisert på å fange lys med forskjellige bølgelengder, hverandre med dybde.
Fargen på dyr endres like naturlig med dybden. Innbyggerne i de littorale og sublitorale sonene er mest klare og varierte farger. Mange dype organismer, som huleorganismer, har ikke pigmenter. I skumringssonen er rødfarging utbredt, som er komplementær til det blåfiolette lyset på disse dypene. Stråler med ekstra farge absorberes mest fullstendig av kroppen. Dette gjør at dyr kan gjemme seg for fiender, siden deres røde farge i blåfiolette stråler visuelt oppfattes som svart. Rød farge er typisk for slike dyr skumringstid, som havabbor, røde koraller, forskjellige krepsdyr, etc.
Hos noen arter som lever nær overflaten av vannforekomster, er øynene delt i to deler med ulike evner til brytning av stråler. Den ene halvparten av øyet ser i luften, den andre i vann. En slik "fireøydhet" er karakteristisk for spinnebiller, den amerikanske fisken Anableps tetraphthalmus og en av de tropiske artene av blenny Dialommus fuscus. Ved lavvann sitter denne fisken i fordypninger og avslører en del av hodet fra vannet (se fig. 26).
Absorpsjonen av lys er sterkere, jo lavere gjennomsiktighet av vannet er, som avhenger av antall partikler som er suspendert i det.
Gjennomsiktighet kjennetegnes ved den maksimale dybden der en spesielt senket hvit skive med en diameter på ca. 20 cm (Secchi-skive) fortsatt er synlig. Det klareste vannet er i Sargassohavet: skiven er synlig til en dybde på 66,5 m. I Stillehavet er Secchi-skiven synlig opptil 59 m, i Det indiske hav - opptil 50, i grunt hav - opp til 5-15 m. Gjennomsiktigheten til elver er i gjennomsnitt 1-1,5 m, og i de gjørmete elvene, for eksempel i sentralasiatiske Amu Darya og Syr Darya, bare noen få centimeter. Grensen til den fotosyntetiske sonen varierer derfor sterkt i ulike vannmasser. I det klareste vannet eufotiske sone, eller sone for fotosyntese, strekker seg til dybder som ikke overstiger 200 m, crepuskulær eller dysfotiske, sonen okkuperer dybder på opptil 1000-1500 m, og dypere, i afotisk sone trenger ikke sollys i det hele tatt.
Mengden lys i de øvre lagene av reservoarene varierer sterkt avhengig av områdets breddegrad og årstiden. Lange polare netter begrenser i stor grad tiden tilgjengelig for fotosyntese i arktiske og antarktiske bassenger, og isdekke gjør det vanskelig for lys å nå alle frosne vannmasser om vinteren.
I havets mørke dyp bruker organismer lys som sendes ut av levende ting som en kilde til visuell informasjon. Gløden til en levende organisme kalles bioluminescens. Lysende arter finnes i nesten alle klasser av vannlevende dyr fra protozoer til fisk, så vel som blant bakterier, lavere planter og sopp. Bioluminescens ser ut til å ha forekommet flere ganger i forskjellige grupper på ulike stadier utvikling.
Kjemien til bioluminescens er nå ganske godt forstått. Reaksjonene som brukes til å generere lys er varierte. Men i alle tilfeller er dette oksidasjon av kompleks organiske forbindelser (luciferiner) ved bruk av proteinkatalysatorer (luciferase). Luciferiner og luciferaser har forskjellige strukturer i forskjellige organismer. Under reaksjonen frigjøres overskuddsenergien til det eksiterte luciferinmolekylet i form av lyskvanter. Levende organismer sender ut lys i impulser, vanligvis som respons på stimuli som kommer fra det ytre miljøet.
Glød spiller kanskje ikke en spesiell økologisk rolle i livet til en art, men kan være et biprodukt av den vitale aktiviteten til celler, som for eksempel i bakterier eller lavere planter. Det får økologisk betydning bare hos dyr som har et tilstrekkelig utviklet nervesystem og visuelle organer. Hos mange arter får de selvlysende organene en svært kompleks struktur med et system av reflektorer og linser som forsterker stråling (fig. 40). En rekke fisk og blekksprut, som ikke er i stand til å generere lys, bruker symbiotiske bakterier som formerer seg i de spesielle organene til disse dyrene.

Ris. 40. Luminescensorganer fra vannlevende dyr (ifølge S. A. Zernov, 1949):
1 - en dyphavs breiflabb med en lommelykt over den tannede munnen;
2 - fordeling av lysende organer i fisk av familien. Mystophidae;
3 - lysende organ til fisken Argyropelecus affinis:
a - pigment, b - reflektor, c - lysende kropp, d - linse
Bioluminescens har hovedsakelig en signalverdi i dyrenes liv. Lyssignaler kan tjene til orientering i en flokk, tiltrekke individer av det motsatte kjønn, lokke ofre, for kamuflasje eller distraksjon. Et lysglimt kan fungere som et forsvar mot et rovdyr ved å blende eller desorientere det. For eksempel frigjør dyphavsblekksprut, som flykter fra en fiende, en sky av lysende sekresjon, mens arter som lever i opplyste vann bruker mørk væske til dette formålet. I noen bunnormer - polychaetes - utvikles lysende organer i løpet av modningen av reproduktive produkter, og hunnene lyser lysere, og øynene er bedre utviklet hos menn. Hos rovfisk fra breiflabben blir den første strålen av ryggfinnen forskjøvet til overkjeven og omgjort til en fleksibel "stang" som på enden bærer et ormeformet "agn" - en kjertel fylt med slim med lysende bakterier. Ved å regulere blodstrømmen til kjertelen og dermed oksygentilførselen til bakterien, kan fisken frivillig få «agnet» til å gløde, etterligne ormens bevegelser og lokke inn byttedyr.
Spørsmål 1. Nevn hovedtrekkene i livet til organismer i vannmiljøet, i land-luftmiljøet og i jorda.
Karakteristikkene av livet til organismer i vannmiljøet, grunn-luftmiljøet og i jorda bestemmes av de fysiske og kjemiske egenskapene til disse livsmiljøene. Disse egenskapene har en betydelig innvirkning på virkningen av andre faktorer av livløs natur - de stabiliserer sesongmessige temperatursvingninger (vann og jord), endrer gradvis belysning (vann) eller eliminerer det fullstendig (jord), etc.
Vann er et tett medium sammenlignet med luft, har en flytekraft og er et godt løsemiddel. Derfor er mange organismer som lever i vann preget av dårlig utvikling av støttevev (vannplanter, protozoer, coelenterater, etc.), spesielle bevegelsesmetoder (sveving, jetfremdrift), pusteegenskaper og tilpasninger for å opprettholde et konstant osmotisk trykk i celler som danner kroppen deres.
Lufttettheten er mye lavere enn vanntettheten, så terrestriske organismer har høyt utviklet støttevev - det indre og ytre skjelettet.
Jord er det øverste laget av land som er transformert som et resultat av aktiviteten til levende vesener. Mellom jordpartiklene er det mange hulrom som kan fylles med vann eller luft. Derfor er jorda bebodd av både vannlevende og luftpustende organismer.
Spørsmål 2. Hvilke tilpasninger har organismer utviklet for å leve i et vannmiljø?
Vannmiljøet er tettere enn luften, noe som bestemmer tilpasninger til bevegelse i det.
Aktiv bevegelse i vann krever en strømlinjeformet kroppsform og velutviklede muskler (fisk, blekksprut - blekksprut, pattedyr - delfiner, sel).
Planktoniske organismer (flytende i vann) har tilpasninger som øker oppdriften deres, som å øke den relative overflaten av kroppen på grunn av tallrike fremspring og setae; reduksjon i tetthet på grunn av akkumulering av fett og gassbobler i kroppen (encellede alger, protozoer, maneter, små krepsdyr).
Organismer som lever i et vannmiljø er også preget av tilpasninger for å opprettholde vann-saltbalansen. Ferskvannsarter har tilpasninger for å fjerne overflødig vann fra kroppen. Dette betjenes for eksempel av ekskresjonsvakuoler i protozoer. I saltvann er det tvert imot nødvendig å beskytte kroppen mot dehydrering, som oppnås ved å øke konsentrasjonen av salter i kroppen.
En annen måte å opprettholde vann-saltbalansen på er å flytte til steder med et gunstig saltinnhold.
Og til slutt sikres bestandigheten i kroppens vann-saltmiljø av vann-ugjennomtrengelige integumenter (pattedyr, høyere kreps, vannlevende insekter og deres larver).
Planter trenger lysenergi fra solen for å leve, så vannplanter lever bare på de dypene der lyset kan trenge inn (vanligvis ikke mer enn 100 m). Med økende habitatdybde i planteceller endres sammensetningen av pigmenter som deltar i prosessen med fotosyntese, noe som gjør det mulig å fange opp deler av solspekteret som trenger inn i dypet.
Spørsmål 3. Hvordan unngår organismer negative effekter av lave temperaturer?
Ved lave temperaturer er det fare for at stoffskiftet stopper, så organismer har utviklet spesielle tilpasningsmekanismer for å stabilisere det.
Planter er minst tilpasset plutselige temperatursvingninger. Når temperaturen synker kraftig under 0 °C, kan vannet i vevene bli til is, noe som kan skade dem. Men planter er i stand til å motstå små negative temperaturer ved å binde frie vannmolekyler til komplekser som ikke er i stand til å danne iskrystaller (for eksempel ved å akkumulere opptil 20-30 % sukker eller fete oljer i cellene).
Med en gradvis nedgang i temperaturen under sesongmessige klimaendringer, begynner en hvileperiode i livet til mange planter, ledsaget av enten delvis eller fullstendig død av jordiske vegetative organer (urteaktige former), eller en midlertidig opphør eller nedbremsing av de viktigste fysiologiske prosessene - fotosyntese og transport av stoffer.
Hos dyr er den mest pålitelige beskyttelsen mot lave miljøtemperaturer varmblodighet, men ikke alle har det. Følgende måter å tilpasse dyr til lave temperaturer kan skilles: kjemisk, fysisk og atferdsmessig termoregulering.
Kjemisk termoregulering er assosiert med en økning i varmeproduksjon med synkende temperatur gjennom intensivering av redoksprosesser. Denne banen krever utgifter til en stor mengde energi, så dyr i tøffe klimatiske forhold trenger mer mat. Denne typen termoregulering utføres refleksivt.
Mange kaldblodige dyr er i stand til å vedlikeholde optimal temperatur kroppen på grunn av musklenes arbeid. For eksempel, i kjølig vær, varmer humler opp kroppen ved å skjelve til 32-33 °C, noe som gir dem muligheten til å ta av og mate.
Fysisk termoregulering er assosiert med tilstedeværelsen av spesielle kroppsbelegg hos dyr - fjær eller hår, som på grunn av deres struktur danner et luftgap mellom kroppen og miljøet, siden det er kjent at luft er en utmerket varmeisolator. I tillegg akkumulerer mange dyr som lever under tøffe klimatiske forhold subkutant fett, som også har varmeisolerende egenskaper.
Atferdsmessig termoregulering er assosiert med å bevege seg i rommet for å unngå temperaturer som er ugunstige for livet, skape tilfluktsrom, samle seg i grupper, endre aktivitet til forskjellige tider av døgnet eller året.
Spørsmål 4. Hva er hovedtrekkene til organismer som bruker kroppen til andre organismer som habitat?
Leveforholdene inne i en annen organisme er preget av større konstanthet sammenlignet med miljøforhold, derfor mister organismer som finner en plass i kroppen til planter eller dyr ofte fullstendig organene og systemene som er nødvendige for frittlevende arter (sanseorganer, bevegelsesorganer, fordøyelse , etc. ), men samtidig utvikler de enheter for å holde i vertens kropp (kroker, sugekopper, etc.) og effektiv reproduksjon.
Hvordan laste ned et gratis essay? . Og en link til dette essayet; Biosfære. Bomiljøer allerede i bokmerkene dine.Ytterligere essays om dette emnet
Abiotiske miljøfaktorer og deres påvirkning på levende organismer Formål: å avsløre egenskapene til abiotiske miljøfaktorer og vurdere deres påvirkning på levende organismer. Mål: å introdusere studentene til miljømessige miljøfaktorer; avsløre funksjonene til abiotiske faktorer, vurder påvirkningen av temperatur, lys og fuktighet på levende organismer; identifisere forskjellige grupper av levende organismer avhengig av påvirkningen av forskjellige abiotiske faktorer på dem; fullføre en praktisk oppgave for å identifisere grupper av organismer avhengig av den abiotiske faktoren. Utstyr: datapresentasjon, oppgaver
Spørsmål 1. Hvilke typer organismer spiller hovedrollen for å opprettholde næringssyklusen? Biogene elementer er de som finnes i et økosystem og er nødvendige for dets liv - makrotrofiske og mikrotrofiske. De er konstant assosiert, og blir en del av biomassen til økosystemet, noe som reduserer mengden de blir igjen i økosystemmiljøet. Hvis organismene ikke brytes ned på grunn av aktiviteten til nedbrytere (heterotrofe organismer), ville tilførselen av næringsstoffer være oppbrukt og økosystemets liv ville opphøre. Derfor kan det hevdes at det er reduksjonsmennene som spiller
Spørsmål 1. Hvilke tilpasninger til endringer i miljøtemperaturen har planter og dyr? Hvilestadiene til organismer - cyster, insektpupper, plantefrø - tåler temperaturendringer godt. Sporer av enkelte bakterier tåler temperatursvingninger fra -273 til +140 °C. Varmblodige dyr - fugler og pattedyr - holder konstant kroppstemperatur ved hjelp av høy level metabolisme, perfekt termoregulering og god varmeisolasjon. For eksempel noen hvaler og pinnipeds, på grunn av tilstedeværelsen av et tykt lag
Spørsmål 1. Hvilke epoker er jordens historie delt inn i? I jordens historie skilles det ut følgende epoker, hvis navn er av gresk opprinnelse: Katarchean (under den eldste), Archean (gammel), Proterozoic (primært liv), Paleozoic (gammelt liv), Mesozoic (midten) liv), kenozoisk (nytt liv). Spørsmål 2. Hvordan påvirket aktivitetene til levende organismer endringen i sammensetningen av planetens atmosfære? Den eldgamle atmosfæren inkluderte metan, ammoniakk, karbondioksid, hydrogen, vanndamp og andre uorganiske forbindelser. Som et resultat av livsaktiviteten til de første organismene i atmosfæren
1. Hva er det særegne ved fordelingen av liv i havet? Livet i verdenshavet er utbredt, men artssammensetningen og tettheten av planter og dyr i havvannet er ekstremt mangfoldig og ujevn. Levende organismer tilpasser seg livet under visse forhold dannet av en kombinasjon av ulike egenskaper ved vannmasser. 2. Hva bestemmer fordelingen av organismer i overflatelaget av vann? Fordelingen av organismer i overflatelaget avhenger av tilstedeværelsen av oksygen i vannet, overflod av næringsstoffer, saltholdighet, temperatur og tetthet
Khodchenkova Galina Mikhailovna Biologilærer, kommunal utdanningsinstitusjon "Zharkovskaya Secondary School No. 1" Tver-regionen Biologitime i 5. klasse "Organisk habitat" Hensikt med leksjonen: Å generalisere og utdype kunnskap om habitatene til levende organismer; å danne seg en idé om det organismiske habitatet, dets betingelser. Leksjonsmål: Pedagogisk: å utdype og utvide kunnskapen om habitatene til levende organismer; å avsløre trekk ved organismens habitat og trekk ved organismers tilpasningsevne til livsbetingelser i andre organismer. Utviklingsmessig: å utvikle selvstendig aktivitet; evnen til raskt å navigere i informasjonsflyten: finne det som trengs, forstå og bruke det som mottas
Spørsmål 1. Hva er virkningen av levende organismer på miljøet? Som et resultat av påvirkningen av levende organismer på miljøet, dets fysiske og Kjemiske egenskaper(gasssammensetning av luft og vann, struktur og egenskaper til jord og jevnt klima i området). Spørsmål 2. Hvilke typer effekter av levende organismer på miljøet kjenner du til? Typer av påvirkning av levende organismer på miljøet: 1) mekanisk (endringer i den mekaniske sammensetningen av jorda, filtrering av vann og luft, bevegelse av stoffer); 2) fysisk-kjemisk (endring i den kjemiske sammensetningen av vann, luft,