유 전적으로 러시아인이 더 가까운 유럽인. 러시아 우크라이나인, 벨로루시인, 타타르인, 슬라브인, 백인, 유태인, 핀란드인 및 기타 인구의 유전학
국방부는 러시아 시민의 생체 재료 실험을 인정했습니다.
미국인에 의한 생물 무기 개발 가능성에 대한 종말론적 가정은 강력한 확인을 받았습니다. 미 국방부는 러시아 시민으로부터 생체 재료를 수집한 사실을 인정했다.
보 다우니 미 국방부 대변인에 따르면 미 공군 59항공 의료 그룹의 분자 연구 센터는 외상과 관련된 다양한 바이오마커를 식별하기 위해 근골격계 연구를 수행하고 있다. 러시아 원산지 샘플은 첫 번째 배치가 러시아에서 왔기 때문에 필요하며 이제 제어를 위해 동일한 샘플이 필요합니다.
미 공군은 러시아인의 RNA 분자 샘플 12개와 윤활액(관절액) 샘플 27개를 구매할 계획임을 상기하십시오. 이 발표는 미국 정부 조달 포털에 게시되었습니다. 동시에 계약은 러시아의 모든 거주자 중 고객이 유럽인에게만 관심이 있으며 예를 들어 우크라이나에서 온 이민자는 고려하지 않을 것이라고 강조합니다.
러시아인으로부터 생체 재료를 수집하는 문제는 블라디미르 푸틴의 공개 성명 이후 사회에서 논의되기 시작했습니다. 그에 따르면 생체 재료는 "다른 인종 그룹과 다른 지역에 사는 사람들을 위해" 수집됩니다. 지리적 포인트 러시아 연방". "문제는 그들이 왜 이러는 거지?" 대통령은 인권 운동가들과의 회의에서 수사적으로 물었다.
나중에 Dmitry Peskov는 특별 서비스를 통해 얻은 정보에 관한 것이라고 설명했습니다.
푸틴의 발언은 러시아 사회 일부에서 신랄한 조롱을 불러일으켰다. “고대 잉카인들도 그런 더러운 속임수를 두려워했기 때문에 최고 통치자는 쓰레기가 불친절하게 떨어지는 것을 방지하기 위해 대왕이 남긴 머리카락, 침, 손톱 깎기 및 기타 생체 물질을 먹는 임무를 맡은 특별한 하녀까지 두었습니다. 손”이라고 Nikulin 미디어는 Andrey 선생님을 인용했습니다.
그럼에도 불구하고 국가는 입법 차원에서 새로운 위협에 대응할 준비를 하고 있다. 12월에 생체 재료 보호에 관한 법률이 State Duma에 제출될 것으로 예상됩니다. "오늘날 우리는 임상 연구를 수행하는 많은 실험실이 있습니다. 예를 들어 Invitro와 같은 외국 실험실입니다. 우리는 그들을 가장 친밀하게 만들었습니다. "라고 State Duma 교육 및 과학위원회의 제 1 부국장 인 Gennady Onishchenko가 문제를 설명했습니다.
"Invitro"에서 그들은 생체 재료 수출에 대한 관여를 부인했습니다. 차례로 러시아 과학 아카데미 "SP"의 일반 유전학 연구소는이 기관이 생체 재료 수집에 관여했다고보고했습니다. 연구소 직원 Ilya Zakharov-Gezekhus는 "우리 연구소의 과학 책임자 인 Nikolai Yankovsky 학자가 방금 생체 재료 수집 프로그램을 조직하고 이끌었습니다. "라고 말했습니다. Yankovsky "SP"에 직접 연락하는 것은 불가능했습니다.
상위 연구원러시아 과학 아카데미 발달 생물학 연구소의 유전학 실험실 Alexey Kulikov는 그의 미국 동료를 정당화하는 경향이 있습니다.
미국인들이 무엇을 하고 있었는지 이해해야 합니다. 그들은 근골격계에 영향을 받은 환자에서 유전자가 어떻게 작용하는지 살펴보고 윤활액의 구성을 살펴보았습니다. 그들은 처음에 슬라브 국가의 아픈 대표자로부터 동유럽 어딘가에서 생체 재료를 구입했기 때문에 러시아, 우크라이나, 벨로루시 등 슬라브 국가 대표의 근골격계에 문제가 없는 사람들의 통제 샘플도 필요합니다.
"SP": - 음, 어떻게? 입찰에는 우크라이나의 생체 재료가 고려되지 않을 것이라고 명시적으로 명시되어 있습니다. 러시아인이 필요해...
그것은 여전히 어떤 의도와도 관련이 없습니다. 유전자 무기는 모두 넌센스입니다. 이것은 비현실적입니다. 세상의 사람들은 너무 다형성이 있기 때문입니다. 매우 다양합니다. 어떤 사람에게는 효과가 있고 다른 사람에게는 효과가 없는 것을 생각해내는 것은 어렵습니다. 나는 여기에서 생각한다 우리 대화하는 중이 야실험 조건에 대해서만. 먹다 과학적 경험그리고 통제가 있습니다. 통제 물질은 같은 지역에서 나온 것이어야 합니다.
"SP":-군대가 이러한 "과학적 연구"를 수행하지 않았다면 국방부는 ...
과학 연구는 군대에서도 수행할 수 있습니다. 또한 법 집행 기관 대표가 해결하는 특정 의료 업무도 있습니다. 아마도 우리는 조종사의 낮은 이동성에 대해 이야기하고 있습니다. 그들은 반드시 오랫동안앉다. 따라서 근골격계에 문제가 없는 것이 근본적으로 중요하다. 따라서 미국인들은 이러한 질병과 이를 담당하는 특정 유전자에 관심이 있습니다.
차례로 러시아 과학 아카데미 일반 유전학 연구소의 실험실 책임자 인 Sergey Kiselev는 유전 무기를 만드는 것이 너무 위험하고 사람을 죽이는 것이 더 쉽다고 믿습니다. 전통적인 방법.
러시아의 생체 재료가 이전되었으며 이전 중입니다. 적어도 Onishchenko가 말했듯이 임상 시험의 틀 내에서. 지난 25년 동안 수십 개의 외국 회사가 러시아에서 임상 약물 시험을 수행해 왔기 때문입니다. 약물이 어떻게 작용하는지 이해하려면 생체 재료가 필요합니다.
물론 이러한 샘플은 위의 목적과 다른 목적을 위해 사용될 수 있습니다. 그것은 여전히 국가 생물학적 샘플로 남아 있기 때문입니다. 가장 중요한 것은 샘플을 사용하여 얻은 정보가 나중에 어떻게 사용되는지입니다. 유전 정보는 개인적인 이득을 포함하여 다양한 목적으로 사용될 수 있습니다.
"SP": - 뭐?
오늘날의 기술을 통해 각 사람의 게놈을 자세히 확인할 수 있습니다. 즉, CIA 건물 식당에있는 숟가락에서 DNA 샘플을 채취하면 정보관이 입사 지원시 쓴 것처럼 실제로 미네소타 주에서 왔는지 아니면 동부 시베리아에서 왔는지 확인할 수 있습니다. . 즉, 사람을 장소에 매우 정확하게 묶을 수 있습니다.
그러나이를 위해서는 먼저 만들어야합니다 유전자 지도영토. 전 세계 많은 국가의 관심있는 정보 기관이 오랫동안 그러한 영토지도를 만들어 왔다고 생각합니다. 사람을 식별할 수 있도록 다른 상황.
예를 들어, 위험한 직업을 가진 사람들. 그들 중 하나가 산산조각 나면 유전지도를 사용하여 그가 누구인지, 어디에서 왔는지 확인하고 그에게 경의를 표하는 것이 가능합니다. 즉, 국내․ 그러나이 정보가 제 3 자에게 들어가면 그들의 이익을 위해 사용될 수 있습니다.
"SP": - V 이 경우우리는 유전적이고 민족적인 무기를 만들 확률에 관심이 있습니다 ...
제 생각에는 그런 무기를 만드는 것은 무의미합니다. 첫째, 사람을 죽이기 위해서는 호흡, 혈액 순환 등과 같은 생명 기능에 영향을 미칠 필요가 있습니다. 그것들은 거의 없으며 모든 사람에게 동일합니다. 민족적으로 중요한 중요한 기능을 찾는 것은 거의 불가능합니다.
둘째, 미국인은 누구입니까? 그들은 명목상 국가가 없습니다. 러시아인, 중국인, 앵글로색슨 족을 포함하여 모두 거기에서 섞였습니다. 따라서 러시아인에 대한 무기가 만들어지면 일부 미국인은 러시아인이거나 그 후손이기 때문에 그 행동에 빠질 것입니다.
따라서 지구 전체에 사람들이 더 많이 퍼지고 게놈이 혼합될수록 유전자 무기를 만들 가능성이 줄어듭니다. 수익성이 없습니다. 다른 방법으로 "담그는" 것이 더 쉽고 저렴하며 효율적입니다.
"SP": -그럼에도 불구하고 펜타곤은 그러한 연구를 수행하고 있으며 오늘 그 대표는 이것을 직접 인정했습니다 ...
입찰 조건을 살펴보았습니다. 이 공군 기지는 RNA 샘플을 요청했습니다. 그러나 RNA는 아유전적 물질입니다. 그것은 인간 게놈과 세포의 기능적 상태 사이의 중개자입니다. RNA는 한 번에 각 개별 세포의 전문화를 반영합니다. 즉, 게놈 - 신체의 각 세포의 DNA는 항상 동일합니다. 그리고 각 세포의 RNA는 다릅니다. 매분 게놈이 작동하는 방식을 보여주기 때문입니다. 따라서 엄지발가락에서 채취한 RNA 샘플은 다른 장기의 RNA와 다를 것입니다. 미국인들은 RNA가 정확히 어디에 필요한지 밝히지 않았습니다.
두 번째로 요청한 것은 관절낭의 활액이었습니다. 더욱이 입찰서에는 RNA를 채취한 사람이 아닌 다른 기증자로부터 액체를 채취할 수 있다고 명시돼 있었다. 즉, 미국인들은 RNA와 활액 사이의 연결이 필요하지 않습니다. 서로 관련이 없는 서로 다른 연구를 위해 샘플을 채취한 것 같습니다.
"SP": -그러나 이것은 명확성을 높이지 못했습니다 ...
그곳에서 돈을 세탁했을 수도 있습니다. 또는 학생들은 입찰서를 작성하는 방법을 배웠습니다.
그러나 이고르 니쿨린(Igor Nikulin) 유엔 생물무기위원회 전 위원은 펜타곤의 공격적인 계획에 대해 의심의 여지가 없습니다.
물론 이것은 군사 목적으로 연구를 수행하려는 미국인의 시도입니다. 펜타곤은 인류를 이롭게 하기 위해 고안된 자선 단체나 인도주의 단체가 아닙니다. 오히려 반대로. 여기에서 선택적으로 작용하는 바이러스를 사용할 수 있습니다.
이러한 바이러스는 오늘날에도 여전히 존재합니다. 예를 들어, 에볼라, 라사, 마르부르크. 그들이 일으키는 출혈열은 주로 Negroid 종족, 조류 독감-Mongoloid, Indo-Europeans의 SARS에 작용합니다.
"SP": - 경쟁 조건에서 어떤 이유로 우크라이나는 예외입니다 ...
우크라이나에서는 미국 프로그램이 5-7년 전에 수행되었습니다. 그리고 지금 그들은 전염병만 있습니다. 홍역, 풍진, 결핵, 파상풍, 콜레라 등 그리고 미국인들은 이에 대한 백신을 제공합니다. 매우 편안합니다.
지난 10년 동안 미국은 수백억 달러를 지출하고 전 세계에 400개 이상의 실험실을 설립하여 새로운 유형의 생물학 무기와 백신을 개발했습니다. 약 40개의 실험실이 여러 국가에 있습니다. 구소련. 이들은 우크라이나, 몰도바, 조지아, 아르메니아, 아제르바이잔, 우즈베키스탄, 키르기스스탄 및 카자흐스탄입니다. 이것이 통제된 혼돈입니다.
"SP": -하지만 우리 러시아 유전 학자들은 하나 또는 다른 민족 집단을 겨냥한 무기 개발 가능성에 대해 회의적입니다 ...
그들에게 남은 일은 무엇입니까? 그들 중 많은 사람들이 보조금을 받고 있습니다. 내가 보조금을 받았다면 나는 또한 침묵했을 것입니다. 그러나 미국인들이 저에게 보조금을 제공하지 않을 것이기 때문에 저는 이 주제에 대해 자유롭게 말할 수 있습니다.
뉴스 평가하기
파트너 뉴스:본질적으로 모든 사람의 유전자 코드는 각 부모로부터 물려받은 모든 유전 정보를 저장하는 23 쌍의 염색체를 갖도록 배열됩니다.
염색체의 형성은 감수분열 시 발생하며, 교차 과정에서 각각 무작위로 모계 염색체에서 절반, 부계 염색체에서 절반을 가져오며, 특정 유전자는 어머니로부터 물려받게 되고 어떤 유전자는 아버지로부터 물려받게 됩니다. 알려지지 않은 모든 것은 우연히 결정됩니다.
이 복권에는 남자 염색체 Y 하나만이 참여하지 않고, 마치 릴레이 배턴처럼 아버지에게서 아들에게로 온전히 전달된다. 여성에게는 이 Y 염색체가 전혀 없다는 점을 분명히 하겠습니다.
각 후속 세대에서 유전자좌라고 하는 Y 염색체의 특정 영역에서 돌연변이가 발생하며, 이는 모든 후속 세대에 다음 세대로 전달됩니다. 남성 명사.
이러한 돌연변이 덕분에 속의 재구성이 가능해졌습니다. Y 염색체에는 약 400개 정도의 유전자좌가 있지만 비교 일배체형 분석 및 속 재구성에 사용되는 유전자좌는 약 100개에 불과합니다.
소위 유전자좌 또는 STR 마커라고도 하는 7~42개의 탠덤 반복이 있습니다. 전반적인 그림그것은 각 개인에게 고유합니다. 특정 수의 세대가 지나면 돌연변이가 발생하고 탠덤 반복의 수가 위 또는 아래로 변경되므로 공통 트리에서 돌연변이가 많을수록 일배체형 그룹의 공통 조상이 더 오래된 것을 볼 수 있습니다.
하플로그룹 자체는 유전 정보를 가지고 있지 않습니다. 유전 정보는 염색체의 처음 22쌍인 상염색체에 있습니다. 유럽의 유전적 구성요소의 분포를 볼 수 있습니다. Haplogroups는 현대 민족 형성의 새벽에 지나간 날의 표식일 뿐입니다.
러시아인들 사이에서 가장 흔한 하플로그룹은 무엇입니까?
사람들 |
인간 |
||||||||
|---|---|---|---|---|---|---|---|---|---|
동부, 서부 및 남부 슬라브. |
|||||||||
| 러시아인(북쪽) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | |
| 러시아인(센터) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | |
| 러시아인(남쪽) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 3 | |
| 러시아인 (모두위대한 러시아인) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| 벨로루시인 | 574 | 52 | 10 | 3 | 16 | 10 | 3 | 러시아인, 슬라브인, 인도유럽인 및 하플로그룹 R1a, R1b, N1c, I1 및 I2약 8-9천년 전인 고대에는 인도-유럽 어족의 토대를 마련한 언어 집단이 있었습니다. 첫 단계이들은 haplogroup R1a 및 R1b일 가능성이 높습니다. 인도-유럽 어족에는 인도-이란(남아시아), 슬라브, 발트( 동유럽), 켈트족(서유럽), 독일인(중부, 북유럽).아마도 그들은 또한 약 7,000년 전에 이주로 인해 유라시아의 다른 지역으로 갔고 일부는 남쪽과 동쪽(R1a-Z93)으로 이동하여 인도-이란 민족과 언어 (대부분 투르크 민족의 민족 발생에 참여), 일부는 유럽 영토에 남아 있었고 슬라브와 러시아인특히 (R1a-Z283, R1b-L51). 다른 형성 단계에서 이미 고대에 이주 흐름의 교차점이 있었기 때문에 모든 유럽 인종 그룹에 많은 수의 하플로 그룹이 존재했습니다. 슬라브어는 한때 통일된 Balto-Slavic 언어 그룹(아마 후기 Corded Ware의 고고학 문화)에서 나왔습니다. 언어 학자 Starostin의 계산에 따르면 이것은 약 3.3 천년 전에 일어났습니다. 기원전 5세기부터의 시대 서기 4~5세기 조건부 Proto-Slavic, tk로 간주될 수 있습니다. Balts와 Slavs는 이미 분리되었지만 아직 Slavs 자체는 없었으며 조금 후에 AD 4-6 세기에 나타날 것입니다. 슬라브 형성의 초기 단계에서 약 80%는 하플로그룹 R1a-Z280 및 I2a-M423이었습니다. Balts 형성의 초기 단계에서 약 80%는 N1c-L1025 및 R1a-Z92 하플로그룹이었습니다. Balts와 Slavs의 이주의 영향과 교차점은 처음부터였습니다. 왜냐하면 많은 측면에서이 구분은 임의적이며 일반적으로 세부 사항없이 주요 추세만을 반영하기 때문입니다. 이란 언어는 인도 유럽어이며 그 연대는 다음과 같습니다. 기원전 2 천년부터 가장 오래된 것입니다. 기원전 4세기부터 기원전 4세기까지. 서기 9세기까지, 그리고 서기 9세기부터 새로운 것까지. 지금까지. 즉, 중앙 아시아에서 인도와이란으로 인도 유럽 언어를 사용하는 일부 부족이 떠난 후에 가장 오래된이란 언어가 나타납니다. 그들의 주요 haplogroup은 아마도 R1a-Z93, J2a, G2a3이었을 것입니다. 서부이란 언어 그룹은 기원전 5 세기 경에 나중에 나타났습니다. 따라서 Indo-Aryans, Celts, Germans 및 Slavs는 학술 과학에서 Indo-Europeans가되었습니다. 다양한 그룹. 이것은 절대적으로 맞습니다. 유전적 측면에서 인도유럽인의 이질성은 Y-하플로그룹과 상염색체 모두에서 두드러진다. Indo-Iranians는 BMAC의 서아시아 유전 적 영향으로 더 큰 특징이 있습니다. Indian Vedas에 따르면 북쪽 (중앙 아시아에서)에서 인도 (남아시아)로 온 것은 Indo-Aryans였으며 인도 Vedas의 기초를 형성 한 것은 찬송가와 전설이었습니다. 그리고 계속해서 언어학에 대해 살펴보겠습니다. 이 러시아어(및 관련 발트 언어, 예를 들어 한때 존재했던 발토 슬라브어 언어 커뮤니티의 일부인 리투아니아어)는 켈트어, 게르만어 및 기타 언어와 함께 산스크리트어에 상대적으로 가깝기 때문입니다. 큰 인도 유럽 가족의 . 그러나 유전 계획에서 Indo-Aryans는 이미 서아시아 인이 더 많았으며 인도에 접근함에 따라 Vedoid 영향도 강화되었습니다. 그래서 그것은 분명해졌습니다. 하플로그룹 R1a DNA 계보에서 이것은 Slavs의 일부, Turks의 일부 및 Indo-Aryans의 일부에 대한 일반적인 haplogroup입니다 (당연히 환경에 다른 haplogroup의 대표자가 있었기 때문에). 하플로그룹 R1a1러시아 평원을 따라 이주하는 동안 그들은 Mordovians (Erzya 및 Moksha)와 같은 Finno-Ugric 민족의 일부가되었습니다. 지파의 일부(for 하플로그룹 R1a1이것은 Z93의 하위 분기입니다) 이주하는 동안 그들은 약 3500년 전, 즉 기원전 2천년 중반에 이 인도 유럽어를 인도와 이란으로 가져왔습니다. 인도에서는 위대한 파니니의 노력으로 기원전 1천년 중반에 산스크리트어로 개종했고, 페르시아-이란에서는 아리아어가 이란어 그룹의 기초가 되었으며, 그 중 가장 오래된 것은 기원전 2000년경으로 거슬러 올라갑니다. 다음 데이터가 확인되었습니다. DNA 계보언어학은 여기에서 상관 관계가 있습니다. 큰 부분 하플로그룹 R1a1-Z93고대에도 그들은 투르크 민족 집단에 합류했으며 오늘날 여러 가지 방식으로 투르크족의 이주를 표시합니다. 이는 고대의 관점에서 놀라운 일이 아닙니다 하플로그룹 R1a1반면 대표자들은 하플로그룹 R1a1-Z280 Finno-Ugric 부족의 일부 였지만 Slavic 식민지 개척자들이 정착하는 동안 그들 중 많은 사람들이 Slavs에 의해 동화되었지만 지금도 예를 들어 Erzya는 여전히 지배적 인 haplogroup입니다. R1a1-Z280. 이 모든 새로운 데이터는 우리에게 제공할 수 있었습니다. DNA 계보, 특히 선사 시대에 현대 러시아 평원과 중앙 아시아 영토에서 haplogroup 항공사의 대략적인 이동 날짜. 따라서 과학자들은 모든 슬라브, 켈트, 독일인 등에게 언어학의 관점에서 볼 때 사실 인 인도 유럽인의 이름을 부여했습니다. 이 인도 유럽인들은 어디에서 왔습니까? 사실 인도유럽어족은 인도와 이란으로 이주하기 훨씬 이전부터 러시아 평야 전역과 멀리 남쪽으로는 발칸 반도까지, 멀리 서쪽으로는 피레네 산맥까지 있었다. 나중에이 언어는이란과 인도 모두 남아시아로 퍼졌습니다. 그러나 유전적 측면에서 상관관계는 훨씬 작습니다. "현재 과학계에서 유일하게 정당화되고 받아들여지는 것은 인도-이란 언어를 사용하는 부족과 민족과 관련해서만 "아리아인"이라는 용어를 사용하는 것입니다." 그렇다면 인도-유럽 흐름은 서쪽, 유럽 또는 그 반대로 동쪽으로 어느 방향으로 갔습니까? 일부 추정에 따르면 인도유럽어족의 나이는 약 8500년입니다. 인도 유럽인의 조상 고향은 아직 결정되지 않았지만 버전 중 하나에 따르면 남부 또는 북부의 흑해 지역이 될 수 있습니다. 인도에서는 우리가 이미 알고 있듯이 약 3500년 전, 아마도 중앙아시아의 영토에서 인도-아리아어가 도입되었으며, 아리아인 자체는 R1a1-L657, G2a, J2a, J2b, H 등 서부 및 남부 유럽의 Haplogroup R1a167개 마커 일배체형 분석 하플로그룹 R1a1모든 유럽 국가에서 서유럽 방향으로 R1a1 조상의 대략적인 이동 경로를 결정할 수 있습니다. 그리고 계산에 따르면 북쪽의 아이슬란드에서 남쪽의 그리스에 이르기까지 거의 유럽 전역에서 하플로그룹 R1a1의 공통 조상은 약 7000년 전이었습니다!즉, 후손들은 릴레이 경주처럼 자신의 일배 체형을 대대로 자신의 후손에게 물려 주었고, 같은 역사적 장소에서 이주하는 과정에서 흩어졌습니다. 아마도 우랄이나 흑해 저지대로 판명되었습니다. . 현대 지도에서 이들은 주로 동부 및 중부 유럽(폴란드, 벨로루시, 우크라이나, 러시아)에 있는 국가입니다. 그러나 하플로그룹의 더 오래된 일배체형의 범위는 R1a1동쪽으로 - 시베리아로 이어집니다. 그리고 가장 오래되고 가장 변이된 일배체형으로 표시되는 첫 번째 조상의 수명은 75000년 전입니다. 그 당시에는 슬라브인도, 독일인도, 켈트인도 없었습니다. 중부 및 동부 유럽R1a1의 공통 조상인 폴란드는 약 5000년 전에 살았습니다(주로 R1a1-M458 및 Z280의 하위 분기군). 러시아-우크라이나어의 경우-4500년 전, 이는 계산 정확도 내에서 거의 일치합니다.그리고 4세대라고 해도 그런 용어에는 차이가 없습니다. 현대 폴란드에서 하플로그룹 R1a1평균 56%, 일부 지역에서는 최대 62%입니다. 나머지는 대부분 서유럽 하플로그룹 R1b(12%), 스칸디나비아 하플로그룹 I1(17%) 및 발트해 하플로그룹 N1c1 (8%). 체코와 슬로바키아에는 4200년 전에 공통의 원시 슬라브 조상이 살았습니다. 러시아인과 우크라이나인보다 조금 적습니다. 즉, 우리는 현대 폴란드, 체코, 슬로바키아, 우크라이나, 벨로루시, 러시아의 영토에 정착하는 것에 대해 이야기하고 있습니다. 모두 불과 몇 세대 만에 4 천년 이상 전에 말입니다. 고고학에서 그러한 연대 측정의 정확성은 전혀 생각할 수 없는 일입니다. 체코와 슬로바키아의 후예 하플로그룹 R1a1약 40%. 나머지는 대부분 서유럽 R1b(22-28%) 스칸디나비아 I1그리고 발칸 하플로그룹 I2a(누적 18%) 현대 헝가리 영토에서 공통 조상 R1a1은 5000년 전에 살았습니다. 이제 haplogroup R1a1의 자손 중 최대 4분의 1이 있습니다. 나머지는 주로 서유럽 haplogroup R1b(20%)와 결합된 스칸디나비아 I1 및 Balkan I2(총 26%) haplogroup을 가지고 있습니다. 헝가리인이 Finno-Ugric 언어 그룹의 언어를 사용한다는 점을 감안할 때 가장 일반적인 haplogroup은 다음과 같습니다. N1c1마자르족의 고대 헝가리 부자 무덤에서 하플로그룹을 가진 남성의 유해가 주로 발견됩니다. N1c1, 제국 형성에 참여한 부족의 첫 번째 지도자였습니다. 리투아니아와 라트비아에서는 공통조상이 4800년 깊이까지 재구성된다. 기본적으로 현재 Z92, Z280 및 M458 하위 클래드가 있습니다. 리투아니아인들 사이에서 가장 흔한 것은 Baltic haplogroup N1c1로 47%에 이릅니다. 일반적으로 리투아니아와 라트비아는 haplogroup N1c1의 South Baltic subclade L1025가 특징입니다. 일반적으로 상황은 분명합니다. 나는 아이슬란드, 네덜란드, 덴마크, 스위스, 벨기에, 리투아니아, 프랑스, 이탈리아, 루마니아, 알바니아, 몬테네그로, 슬로베니아, 크로아티아, 스페인, 그리스, 불가리아, 몰도바와 같은 유럽 국가에서 공통 조상이 5000- 5500년 전에는 더 정확할 수 없습니다. 이것은 공통 조상입니다. 하플로그룹 R1a나열된 모든 국가에 대해. 약 7500년 전 인도-유럽인의 공통 조상인 위에 표시된 발칸 지역을 제외하고 말하자면 유럽의 공통 조상입니다. 통신사 점유율 하플로그룹 R1a1다음 국가에서는 네덜란드와 이탈리아 4%, 알바니아 9%, 그리스 8-11%(테살로니키 최대 14%), 불가리아와 헤르체고비나 12-15%, 덴마크 14-17%, 세르비아, 보스니아와 마케도니아에서 15-25%, 스위스에서 3%, 루마니아와 헝가리에서 20%, 아이슬란드에서 23%, 몰도바에서 22-39%, 크로아티아에서 29-34%, 슬로베니아에서 30-37%(16 발칸 반도 전체에서 %), 동시에 에스토니아에서 32-37%, 리투아니아에서 34-38%, 라트비아에서 41%, 벨로루시에서 40%, 우크라이나에서 45-54%입니다. 동유럽 러시아에서 하플로그룹 R1a, 이미 언급했듯이 발트해 연안의 높은 점유율로 인해 평균 47 % 하플로그룹 N1c1러시아의 북쪽과 북서쪽, 그러나 남쪽과 러시아의 중앙에서는 haplogroup R1a의 다른 하위 분기의 비율이 55%에 이릅니다. 터키인과 하플로그룹 R1a1조상의 일배 체형은 모든 곳에서 다르며 지역마다 고유 한 하위 분기가 특징입니다. Altai 및 기타 Turks의 사람들은 또한 haplogroup R1a1의 높은 비율을 가지고 있으며 Bashkirs는 subclade Z2123의 40 %에 도달합니다. 이것은 Z93의 자식 계통이며 일반적으로 투르크족이라고 할 수 있으며 인도-이란인의 이주와 관련이 없습니다.오늘은 많은 수의 하플로그룹 R1a1중앙 아시아의 투르크 인구 중 Sayano-Altai 지역에 있습니다. 63%에 도달하는 키르기즈인들 사이에서. 러시아인이나이란 인이라고 부를 수 없습니다. 모든 이름이 밝혀졌습니다. 하플로그룹 R1a1하나의 이름-최소한 과장이지만 대부분-무지. 하플로그룹은 민족 집단이 아니며, 보인자의 언어 및 민족 소속이 기록되지 않습니다. Haplogroups는 또한 유전자와 직접적인 관련이 없습니다. Türks는 주로 Z93의 다양한 하위 클레이드가 특징이지만 Volga 지역에는 R1a1-Z280도 있으며 Volga Finns에서 Volga Türks로 이전되었을 수 있습니다. Haplogroup R1a1-Z93은 또한 Ashkenazi 유대인의 하위 그룹 인 Levites의 경우 중간 빈도의 아랍인에게 일반적입니다 (하위 클레이드 CTS6는 후자에서 확인되었습니다). 이 계통은 이미 초기 단계에서 이 민족의 민족 발생에 참여했습니다. 최초 배포 지역 하플로그룹 R1a1유럽에서 - 이것은 아마도 동유럽의 영토이고 아마도 흑해 저지대 일 것입니다. 그 전에는 아마도 아시아, 아마도 남아시아나 중국 북부였을 것입니다. 백인 R1a1 일배 체형아르메니아. haplogroup의 공통 조상의 나이 R1a1- 6500년 전. R1a1-Z282도 있지만 기본적으로 R1a1-Z93의 하위군이기도 합니다.소아시아, 아나톨리아 반도. 중동, 유럽, 아시아를 잇는 역사적인 교차로. 그것은 "인도 유럽 고향"의 첫 번째 또는 두 번째 후보였습니다. 그러나 하플로그룹 R1a1의 공통 조상은 약 6500년 전에 그곳에 살았다. 일배체형으로 판단할 때 이 조상의 집은 실질적으로 아나톨리아에 있을 수 있거나 원래의 인도-유럽인이 운반자였음이 분명합니다. 하플로그룹 R1b. 그러나 일반 일배체형 데이터베이스에서 터키인 개인의 대표성이 낮을 가능성이 높습니다. 따라서 아르메니아인과 아나톨리아인은 모두 조상이 같거나 조상이 시간이 매우 가깝고 몇 세대 내에 있습니다. 이것은 Z93 및 Z282 *의 하위 클레이드입니다. 아나톨리아에 있는 R1a1-Z93 하플로그룹의 공통 조상보다 4500년 전이 기원전 3천년의 마지막 분기에 소아시아에 히타이트가 출현한 시기와 잘 일치한다는 점에 유의해야 합니다. 비록 많은 R1a1-Z93 이미 우리 시대에 투르크 민족이 한반도로 이주한 후에 거기에 선이 나타날 수 있습니다. 알렉세이 조린 *** | |
이 복권에는 남성 염색체 하나만 Y가 참여하지 않고 마치 지휘봉처럼 아버지에게서 아들에게 완전히 전달됩니다. 여성에게는 이 Y 염색체가 전혀 없다는 점을 분명히 하겠습니다.
각 후속 세대에서 유전자좌라고 하는 Y 염색체의 특정 영역에서 돌연변이가 발생하며, 이는 남성 성별에 따라 모든 후속 세대에 전달됩니다. 이러한 돌연변이 덕분에 속의 재구성이 가능해졌습니다. Y염색체에는 약 1000개 정도의 유전자좌가 존재하는데 일배체형의 비교 분석과 속의 재구성에 사용되는 유전자좌는 100개 조금 넘습니다.
소위 유전자좌(또는 STR 마커라고도 함)에는 7~42개의 탠덤 반복이 있으며, 전체 패턴은 사람마다 고유합니다. 특정 수의 세대가 지나면 돌연변이가 발생하고 탠덤 반복의 수가 위 또는 아래로 변경되므로 공통 트리에서 돌연변이가 많을수록 일배체형 그룹의 공통 조상이 더 오래된 것을 볼 수 있습니다.
하플로그룹 자체는 유전 정보를 가지고 있지 않습니다. 유전 정보는 염색체의 처음 22쌍인 상염색체에 있습니다. 유럽의 유전적 구성요소의 분포를 볼 수 있습니다. Haplogroups는 현대 민족 형성의 새벽에 지나간 날의 표식일 뿐입니다.
러시아인들 사이에서 가장 흔한 하플로그룹은 무엇입니까?
| 사람들 | 수량, 인간 | R1a1, | R1b1, | 나는1, | 나는2, | N1c1, | E1b1b1, | J2, | G2a, |
|---|---|---|---|---|---|---|---|---|---|
| 동부, 서부 및 남부 슬라브. | |||||||||
| 러시아인(북쪽) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | 1 |
| 러시아인(센터) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | 1 |
| 러시아인(남쪽) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 4 | 3 |
| 러시아인 (모두위대한 러시아인) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| 벨로루시인 | 574 | 52 | 10 | 3 | 16 | 10 | 3 | 2 | 2 |
| 우크라이나인 | 93 | 54 | 2 | 5 | 16 | 8 | 8 | 6 | 3 |
| 러시아인(우크라이나인과 벨로루시인과 함께) | 1874 | 48 | 7 | 4 | 13 | 16 | 4 | 3 | 3 |
| 폴란드인 | 233 | 56 | 16 | 7 | 10 | 8 | 4 | 3 | 2 |
| 슬로바키아인 | 70 | 47 | 17 | 6 | 11 | 3 | 9 | 4 | 1 |
| 체코인 | 53 | 38 | 19 | 11 | 12 | 3 | 8 | 6 | 5 |
| 슬로베니아인 | 70 | 37 | 21 | 12 | 20 | 0 | 7 | 3 | 2 |
| 크로아티아인 | 108 | 24 | 10 | 6 | 39 | 1 | 10 | 6 | 2 |
| 세르비아인 | 113 | 16 | 11 | 6 | 29 | 1 | 20 | 7 | 1 |
| 불가리아인 | 89 | 15 | 11 | 5 | 20 | 0 | 21 | 11 | 5 |
| Balts, Finns, Germans, Greeks 등 | |||||||||
| 리투아니아인 | 164 | 34 | 5 | 5 | 5 | 44 | 1 | 0 | 0 |
| 라트비아인 | 113 | 39 | 10 | 4 | 3 | 42 | 0 | 0 | 0 |
| 핀란드(동쪽) | 306 | 6 | 3 | 19 | 0 | 71 | 0 | 0 | 0 |
| 핀란드(서쪽) | 230 | 9 | 5 | 40 | 0 | 41 | 0 | 0 | 0 |
| 스웨덴 | 160 | 16 | 24 | 36 | 3 | 11 | 3 | 3 | 1 |
| 독일인 | 98 | 8 | 48 | 25 | 0 | 1 | 5 | 4 | 3 |
| 독일인(바이에른) | 80 | 15 | 48 | 16 | 4 | 0 | 8 | 6 | 5 |
| 영어 | 172 | 5 | 67 | 14 | 6 | 0.1 | 3 | 3 | 1 |
| 아일랜드의 | 257 | 1 | 81 | 6 | 5 | 0 | 2 | 1 | 1 |
| 이탈리아인 | 99 | 2 | 44 | 3 | 4 | 0 | 13 | 18 | 8 |
| 루마니아인 | 45 | 20 | 18 | 2 | 18 | 0 | 7 | 13 | 7 |
| 오세트인 | 359 | 1 | 7 | 0 | 0 | 1 | 16 | 67 | |
| 아르메니아인 | 112 | 2 | 26 | 0 | 4 | 0 | 6 | 20 | 10 |
| 그리스인 | 116 | 4 | 14 | 3 | 10 | 0 | 21 | 23 | 5 |
| 터키인 | 103 | 7 | 17 | 1 | 5 | 4 | 10 | 24 | 12 |
특히 주목할 만한 것은 러시아인들 사이에서 가장 흔한 4개의 하플로그룹입니다.
R1a1 47.0%, N1c1 20.0%, I2 10.6%, I1 6.2%
말하는 간단한 말로: 유전적 구성 러시아인 Y-염색체의 곧은 남성 라인을 따라 다음과 같습니다.
동유럽인 - 47%
발트 3국 - 20%
그리고 구석기 시대 이후 원래 유럽인의 두 하플로 그룹
스칸디나비아인 - 6%
발칸 반도 - 11%
이름은 조건부이며 영토 최대값에 따라 지정됩니다. 유럽 사람하플로그룹 R1a1, N1c1, I1 및 I2에 대한 서브클레이드. 근본적인 점은 200년의 타타르-몽골 멍에 이후 몽골의 후예가 없었다는 것이다. 어느 쪽이든 떠났지만 그러한 관계에서 매우 적은 수의 직접적인 유전 상속인이 있습니다. 이 말로는 묻고 싶지 않아 역사적 출처 Rus의 몽골 인에 대해, 그러나 러시아인에 대한 Mongol-Tatars의 유전 적 영향에주의를 기울이기 위해서만 있습니다. 거기에 없거나 중요하지 않습니다. 그건 그렇고, Bulgar Tatars의 게놈에는 많은 수의 보균자가 있습니다. 하프로그룹 R1a1(약 30%) 및 N1c1(약 20%), 그러나 그들은 대부분 비유럽인입니다.
또 다른 중요한 점, 남부 러시아인은 오차 범위 내에서 우크라이나인과 다르지 않으며 우세한 하플로그룹 R1a1 중 하나를 갖는 북부 러시아인도 하플로그룹 N1c1의 비율이 더 높습니다. 그러나 N1c1 일배체형의 비율은 러시아인에서 평균 20%입니다.
황제. 니콜라스 2세
올덴부르크 대공 가문의 최초 알려진 조상은 1091년 연대기에 언급된 레리가우 백작 에길마르(1108년 사망)였습니다.
Nicholas II는 haplogroup의 운반자로 밝혀졌습니다. R1b1a2-Holstein-Gottorp 왕조의 서유럽 라인 대표. 이 독일 왕조는 다음에서 가장 흔한 말단 싹둑 U106이 특징입니다. 유럽 북서부게르만 부족의 정착지에서. 완전 전형적이진 않네요 러시아인 DNA 마커이지만 러시아인 사이에 존재하는 것은 독일인과 슬라브인 사이의 초기 접촉과도 연관될 수 있습니다.
자연 왕자. 루리코비치
Vladimir Monomakh와 "Monomachis"라고하는 그의 후손은 haplogroup에 속합니다. N1c1-L550, South Baltic 지역 (subclade L1025)과 Fennoscandia (subclades Y7795, Y9454, Y17113, Y17415, Y4338)에 널리 퍼져 있습니다. Rurik 왕조는 말단 절단 Y10931이 특징입니다.
역사가들이 Olgoviches라고 부르는 사람들 중 일부는 (봉건 투쟁에서 Vladimir Monomakh의 주요 라이벌 인 Oleg Svyatoslavich의 이름을 따서 명명되었으며 모든 출처에서 알 수 있듯이 그의 사촌) Monomashich 가족의 Rurikovichs와 관련이 없습니다 (직접 남성) 선). 이들은 Yuri Tarussky의 후손입니다.
러시아인, 슬라브인, 인도유럽인 및 하플로그룹 R1a, R1b, N1c, I1 및 I2
약 8-9 천년 전 고대에는 인도 유럽어 계열의 토대를 마련한 언어 그룹이있었습니다 (초기 단계에서는 haplogroups R1a 및 R1b 일 가능성이 큽니다). Indo-European 가족에는 Indo-Iranians (남아시아), Slavs 및 Balts (동유럽), Celts (서유럽), Germans (중부, 북유럽)와 같은 언어 그룹이 포함됩니다. 아마도 그들은 또한 약 7,000년 전에 이주로 인해 유라시아의 다른 지역으로 갔고 일부는 남쪽과 동쪽(R1a-Z93)으로 이동하여 인도-이란 민족과 언어 (대부분 투르크 민족의 민족 발생에 참여), 일부는 유럽 영토에 남아 있었고 슬라브와 러시아인특히 (R1a-Z283, R1b-L51). 다른 형성 단계에서 이미 고대에 이주 흐름의 교차점이 있었기 때문에 모든 유럽 인종 그룹에 많은 수의 하플로 그룹이 존재했습니다.슬라브어는 한때 통일된 Balto-Slavic 언어 그룹(아마 후기 Corded Ware의 고고학 문화)에서 나왔습니다. 언어 학자 Starostin의 계산에 따르면 이것은 약 3.3 천년 전에 일어났습니다. 기원전 5세기부터의 시대 서기 4~5세기 조건부 Proto-Slavic, tk로 간주될 수 있습니다. Balts와 Slavs는 이미 분리되었지만 아직 Slavs 자체는 없었으며 조금 후에 AD 4-6 세기에 나타날 것입니다. 슬라브 형성의 초기 단계에서 약 80%는 하플로그룹 R1a-Z280 및 I2a-M423이었습니다. Balts 형성의 초기 단계에서 약 80%는 N1c-L1025 및 R1a-Z92 하플로그룹이었습니다. Balts와 Slavs의 이주의 영향과 교차점은 처음부터였습니다. 왜냐하면 많은 측면에서이 구분은 임의적이며 일반적으로 세부 사항없이 주요 추세만을 반영하기 때문입니다.
이란 언어는 인도 유럽어이며 그 연대는 다음과 같습니다. 기원전 2 천년부터 가장 오래된 것입니다. 기원전 4세기부터 기원전 4세기까지. 서기 9세기까지, 그리고 서기 9세기부터 새로운 것까지. 지금까지. 즉, 중앙 아시아에서 인도와이란으로 인도 유럽 언어를 사용하는 일부 부족이 떠난 후에 가장 오래된이란 언어가 나타납니다. 그들의 주요 haplogroup은 아마도 R1a-Z93, J2a, G2a3이었을 것입니다. 서부이란 언어 그룹은 기원전 5 세기 경에 나중에 나타났습니다.
따라서 Indo-Aryans, Celts, Germans 및 Slavs는 학술 과학에서 Indo-Europeans가되었으며, 이 용어는 그러한 방대하고 다양한 그룹에 가장 적합합니다. 이것은 절대적으로 맞습니다. 유전적 측면에서 인도유럽인의 이질성은 Y-하플로그룹과 상염색체 모두에서 두드러진다. Indo-Iranians는 BMAC의 서아시아 유전 적 영향으로 더 큰 특징이 있습니다.
Indian Vedas에 따르면 북쪽 (중앙 아시아에서)에서 인도 (남아시아)로 온 것은 Indo-Aryans였으며 인도 Vedas의 기초를 형성 한 것은 찬송가와 전설이었습니다. 그리고 계속해서 언어학에 대해 살펴보겠습니다. 이 러시아어(및 관련 발트 언어, 예를 들어 한때 존재했던 발토 슬라브어 언어 커뮤니티의 일부인 리투아니아어)는 켈트어, 게르만어 및 기타 언어와 함께 산스크리트어에 상대적으로 가깝기 때문입니다. 큰 인도 유럽 가족의 . 그러나 유전 계획에서 Indo-Aryans는 이미 서아시아 인이 더 많았으며 인도에 접근함에 따라 Vedoid 영향도 강화되었습니다.
그래서 그것은 분명해졌습니다. 하플로그룹 R1a DNA 계보에서 이것은 Slavs의 일부, Turks의 일부 및 Indo-Aryans의 일부에 대한 일반적인 haplogroup입니다 (당연히 환경에 다른 haplogroup의 대표자가 있었기 때문에). 하플로그룹 R1a1러시아 평원을 따라 이주하는 동안 그들은 Mordovians (Erzya 및 Moksha)와 같은 Finno-Ugric 민족의 일부가되었습니다. 지파의 일부(for 하플로그룹 R1a1이것은 Z93의 하위 분기입니다) 이주하는 동안 그들은 약 3500년 전, 즉 기원전 2천년 중반에 이 인도 유럽어를 인도와 이란으로 가져왔습니다. 인도에서는 위대한 파니니의 노력으로 기원전 1천년 중반에 산스크리트어로 개종했고, 페르시아-이란에서는 아리아어가 이란어 그룹의 기초가 되었으며, 그 중 가장 오래된 것은 기원전 2000년경으로 거슬러 올라갑니다. 다음 데이터가 확인되었습니다. DNA 계보언어학은 여기에서 상관 관계가 있습니다.
큰 부분 하플로그룹 R1a1-Z93고대에도 그들은 투르크 민족 집단에 합류했으며 오늘날 여러 가지 방식으로 투르크족의 이주를 표시합니다. 이는 고대의 관점에서 놀라운 일이 아닙니다 하플로그룹 R1a1반면 대표자들은 하플로그룹 R1a1-Z280 Finno-Ugric 부족의 일부 였지만 Slavic 식민지 개척자들이 정착하는 동안 그들 중 많은 사람들이 Slavs에 의해 동화되었지만 지금도 예를 들어 Erzya는 여전히 지배적 인 haplogroup입니다. R1a1-Z280.
이 모든 새로운 데이터는 우리에게 제공할 수 있었습니다. DNA 계보, 특히 선사 시대에 현대 러시아 평원과 중앙 아시아 영토에서 haplogroup 항공사의 대략적인 이동 날짜.
따라서 과학자들은 모든 슬라브, 켈트, 독일인 등에게 언어학의 관점에서 볼 때 사실 인 인도 유럽인의 이름을 부여했습니다.
이 인도 유럽인들은 어디에서 왔습니까? 사실 인도유럽어족은 인도와 이란으로 이주하기 훨씬 이전부터 러시아 평야 전역과 멀리 남쪽으로는 발칸 반도까지, 멀리 서쪽으로는 피레네 산맥까지 있었다. 나중에이 언어는이란과 인도 모두 남아시아로 퍼졌습니다. 그러나 유전적 측면에서 상관관계는 훨씬 작습니다.
"현재 과학계에서 유일하게 정당화되고 받아들여지는 것은 인도-이란 언어를 사용하는 부족과 민족과 관련해서만 "아리아인"이라는 용어를 사용하는 것입니다."
그렇다면 인도-유럽 흐름은 서쪽, 유럽 또는 그 반대로 동쪽으로 어느 방향으로 갔습니까? 일부 추정에 따르면 인도유럽어족의 나이는 약 8500년입니다. 인도 유럽인의 조상 고향은 아직 결정되지 않았지만 버전 중 하나에 따르면 남부 또는 북부의 흑해 지역이 될 수 있습니다. 인도에서는 우리가 이미 알고 있듯이 약 3500년 전, 아마도 중앙아시아의 영토에서 인도-아리아어가 도입되었으며, 아리아인 자체는 R1a1-L657, G2a, J2a, J2b, H 등
서부 및 남부 유럽의 Haplogroup R1a1
67개 마커 일배체형 분석 하플로그룹 R1a1모든 유럽 국가에서 서유럽 방향으로 R1a1 조상의 대략적인 이동 경로를 결정할 수 있습니다. 그리고 계산에 따르면 북쪽의 아이슬란드에서 남쪽의 그리스에 이르기까지 거의 유럽 전역에서 하플로그룹 R1a1의 공통 조상은 약 7000년 전이었습니다! 즉, 후손들은 릴레이 경주처럼 자신의 일배 체형을 대대로 자신의 후손에게 물려 주었고, 같은 역사적 장소에서 이주하는 과정에서 흩어졌습니다. 아마도 우랄이나 흑해 저지대로 판명되었습니다. . 현대 지도에서 이들은 주로 동부 및 중부 유럽(폴란드, 벨로루시, 우크라이나, 러시아)에 있는 국가입니다. 그러나 하플로그룹의 더 오래된 일배체형의 범위는 R1a1동쪽으로 - 시베리아로 이어집니다. 그리고 가장 오래되고 가장 변이된 일배체형으로 표시되는 첫 번째 조상의 수명은 75000년 전입니다. 그 당시에는 슬라브인도, 독일인도, 켈트인도 없었습니다.방법의 단점
당신이 테스트를 했고 그것이 당신을 매우 기쁘게 했다면, 나는 서둘러 타르 국자를 가져왔습니다. 예, Y염색체는 아버지에게서 아들로 거의 변하지 않고 유전되지만 실제로는 유전적으로 유전됩니다. 유용한 정보그 안에는 없지만 다른 쌍의 염색체에는 훨씬 더 많은 유전자가 있습니다.
그리고 이 다른 22개는 매우 무작위적인 방식으로 섞이며 Y에는 그러한 혼합의 흔적이 남지 않습니다.
상상하다. 앵글로색슨 선원들은 흑인 국가를 점령했습니다. 여성은 그러한 여행에 참여하지 않으며 지역 주민들과 연락을 취해야 합니다. 옵션은 무엇입니까?
1) 앵글로 색슨 족은 흑인 여성의 자녀가 있지만 국적은 소년에게만 이전됩니다. 이 경우 Y 염색체는 유럽인에게 이전되지만 실제로 중요한 유럽 유전자의 비율은 감소합니다. 1 세대는 흑인의 절반이 될 것이고 그러한 경우 이전의 "귀족"은 Y가 이것에서 나올지라도 빠르게 해산 될 것입니다. 인종 그룹. 그다지 말이되지 않습니다. Finns와 Indians에게도 비슷한 일이 일어 났을 것입니다. Yakuts와 Finns는 N1c1 haplogroup 특성의 비율이 가장 높지만 유 전적으로 이들은 6 천년 이상 전에 분리 된 고유 한 역사를 가진 N1c1 haplogroup의 다른 하위 분기를 가진 완전히 다른 민족입니다. 그 반대의 경우도 마찬가지입니다. 인도인 - 높은 비율 하플로그룹 R1a1유 전적으로 그들은이 haplogroup, tk의 유럽 대표와 공통점이 거의 없습니다. 또한 6,000년 이상 전에 분리된 고유한 역사를 가진 다양한 하위 분기군입니다.
2) Indo-Aryans 만족 카스트 제도. 1세대도 반흑인이지만, 귀족이 그들끼리만 교배한다면 원래 유전학의 비율은 50% 정도 떠오를 것입니다. 그러나 실제로 결혼은 주로 현지 여성과 이루어지며 정복자의 원래 유전자 풀을 얻는 것은 더욱 불가능합니다. 그리고 이것은 지구의 역사에서 일어났습니다. 상류층인도인의 20%에서 72%는 하플로그룹 R1a1(평균 43%), 그러나 유 전적으로 그들은 같은 유럽 또는 투르크 대표와 공통점이 거의 없습니다 하플로그룹 R1a1, 그리고 다시 그 이유는 고유한 역사를 가진 다양한 하위 클래드입니다.
Y가 최대 95%까지 흔한 중앙 아프리카 국가인 카메룬에서도 비슷한 상황이 발생했을 것입니다. 하플로그룹 R1b-V88, 하지만 동시에 전형적인 인류학적 아프리카 흑인 인구 중 하나입니다.
마커와 하플로그룹의 존재는 국적을 결정하는 중요한 조건이지만 충분조건은 아니라고 결론지을 수 있다. 사람의 국가-영토 출신을 확인하기 위해 Family Tree DNA에는 Family Finder라는 상염색체 검사가 있습니다.
알렉세이 조린 
과학자들은 최근 인간의 유전자 코드를 해독하는 데 근접했습니다. 여러면에서 이것은 이전에 생각했던 것만 큼 동질적이지 않고 더 오래된 것으로 판명 된 러시아 민족의 역사를 새롭게 살펴볼 수있게했습니다.
수세기의 깊이에서
인간 게놈은 가변적인 것입니다. 인류의 진화 과정에서 하플로그룹은 한 번 이상 돌연변이를 겪었습니다. 오늘날 과학자들은 이미 특정 돌연변이가 발생한 대략적인 시간을 결정하는 방법을 배웠습니다. 그래서, 미국 유전학자이러한 돌연변이 중 하나가 약 4,500년 전에 중앙 러시아 평야에서 발생했음을 발견했습니다. 아버지와 다른 뉴클레오티드 세트를 가진 소년이 태어났습니다. 그는 부계 R1a 대신 발생한 유전 분류 R1a1이 지정되었습니다.
다른 많은 돌연변이와 달리 이 돌연변이는 실행 가능한 것으로 판명되었습니다. R1a1 속은 살아남았을 뿐만 아니라 유라시아 대륙의 상당 부분에 정착했습니다. 현재 러시아, 벨로루시, 우크라이나 남성 인구의 약 70%가 R1a1 하플로그룹의 보균자이며 오래된 러시아 도시에서는 이 수치가 80%에 이릅니다. 따라서 R1a1은 러시아 민족 그룹의 일종의 표식 역할을 합니다. 신석기 후기에 살았던 고대 소년의 피가 현대 러시아 대부분의 남성의 정맥에 흐르는 것으로 밝혀졌습니다.
R1a1 haplogroup이 탄생한 지 약 500년 후, 그 대표자들의 이주 흐름은 동쪽으로 - Urals를 넘어 남쪽으로 - Hindustan으로, 서쪽으로 - 현대 영토로 퍼졌습니다. 유럽 국가. 중앙 러시아 평야의 주민들이 원래 범위를 훨씬 넘어 섰다는 사실도 고고학자들에 의해 확인되었습니다. 기원전 1천년 알타이 매장지의 뼈 유적 분석. 이자형. 몽골족 외에도 뚜렷한 백인도 그곳에 살았다는 것을 보여주었습니다.
타타르어는 없다
대중 과학 간행물 The American Journal of Human Genetics의 문제 중 하나에서 러시아인의 유전자 풀에 대한 러시아-에스토니아 과학자 팀의 연구에 대한 기사가 게시되었습니다. 연구원들의 발견은 전혀 예상치 못한 것이었다. 첫째, 러시아 민족은 유전적 특성으로 인해 이질적입니다. 국가의 중부 및 남부 지역에 거주하는 러시아인의 한 부분은 이웃 슬라브 민족과 가깝고 다른 부분은 러시아 북부에 있으며 유 전적으로 Finno-Ugric 민족과 밀접한 관련이 있습니다.
다음 결론은 더 흥미 롭습니다. 과학자들은 러시아 게놈에서 악명 높은 아시아 요소를 감지하지 못했습니다. 러시아 인구에서 눈에 띄는 양의 타타르-몽골 유전자 세트는 없습니다. "러시아인을 긁으십시오-타타르를 찾을 것입니다"라는 안정된 표현이 잘못된 것으로 밝혀졌습니다.
러시아 과학 아카데미 일반 유전학 연구소의 게놈 지리 연구소장인 올레그 발라노프스키 교수는 러시아 유전자 풀을 "거의 완전히 유럽인"으로 간주하고 중앙 아시아 유전자 풀과의 차이점을 "정말 대단하다"고 말합니다. 두 개의 다른 세계라면.
Kurchatov Institute의 게놈 방향 책임자 인 학자 Konstantin Skryabin은 Balanovsky에 동의합니다. 그는 다음과 같이 말합니다. "우리는 몽골 멍에의 파괴적인 영향에 대한 이론을 반박하는 러시아 게놈에서 눈에 띄는 타타르 도입을 찾지 못했습니다." 또한 과학자에 따르면 시베리아 인은 유 전적으로 Old Believers와 동일합니다. 그들은 동일한 "러시아 게놈"을 가지고 있습니다.
연구원들은 또한 한편으로는 러시아인과 다른 한편으로는 이웃 슬라브 민족(우크라이나인, 벨로루시인, 폴란드인) 간의 유전자형의 약간의 차이에 주목합니다. 러시아 북부 주민의 남부와 서부 슬라브의 차이는 더 두드러집니다.
특수 마커
인류학자 바실리 데랴빈에 따르면, 러시아 유전자형도 명확한 생리학적 지표를 가지고 있습니다. 그중 하나는 회색, 파란색, 회색-파란색, 파란색과 같은 러시아인의 밝은 색조가 우세하다는 것입니다. 우리는 그들 중 45%를 가지고 있습니다. 서유럽덜 - 약 35%. 러시아인과 머리 불공평 중 많은 사람들. 인류학자에 따르면 천연 검은 머리색을 가진 러시아인은 5%를 넘지 않습니다. 서유럽에서 검은 머리를 만날 확률은 45%입니다.
대중적인 믿음과는 달리 러시아인들 사이에는 들창코가 많지 않습니다. 약 7%, 약 75%의 경우 코가 곧습니다. 또한 러시아인들 사이에서 epicanthus는 발견되지 않습니다. 눈 안쪽 모서리에있는 몽골족 대표의 전형적인 주름입니다.
러시아 인종 그룹은 유대인들 사이에서 I 및 II 혈액형이 우세한 것이 특징입니다. 예를 들어 그룹 IV가 더 일반적입니다. 생화학 연구에 따르면 러시아인과 다른 유럽인의 혈액에는 특수 유전자 PH-c가 있지만 몽골로이드에는 없습니다.
북부인이 더 가깝다
러시아 과학 아카데미 및 인류학 연구소의 분자 유전학 연구소. D.N. Anuchin Moscow State University는 러시아인의 유전자 풀에 대한 심층 연구를 수행했으며 그 동안 러시아인과 북부 이웃 인 Finns 간의 유전자형 차이가 확립되었습니다. 이는 기존 단위 30 개에 달했습니다. 그러나 러시아 민족과 전통적으로 우리나라 북부에 살았던 Finno-Ugric 민족 (Mordovians, Mari, Veps, Karelians, Komi-Zyryans, Izhors) 간의 유전 적 차이는 세 단위에 해당합니다.
과학자들은 Finno-Ugric 민족과 러시아인의 유전 적 통일성에 대해서만 이야기하는 것이 아니라 그들의 공통 기원에 대해서도 이야기하고 있습니다. 더욱이, 이들 민족 집단의 Y-염색체의 특정 구조는 힌두스탄 민족과 대체로 동일합니다. 그러나 이것은 러시아 사람들의 유전 조상의 정착 방향을 고려할 때 놀라운 일이 아닙니다.
인간 하플로그룹은 직계 남성과 여성 계통을 통해 전염됩니다. 그러나 남성과 여성 모두의 유전학은 DNA의 상염색체에 저장된 정보에 책임이 있습니다. 상염색체는 재조합 과정인 교차 후 양 부모로부터 물려받은 처음 22쌍의 인간 염색체입니다. 따라서 유전 정보의 약 절반은 아버지와 어머니로부터 후손에게 전달됩니다.안에 이 연구 80,000개 이상의 상염색체 싹이 사용되며 기준점은 매우 고해상도, 대부분의 사람들의 유전적 수준에서 상대적으로 작은 영향도 포착할 수 있습니다. 비교 분석 데이터는 비교 분석유전적 요소. 유전자 계산기 자체는 GedMatch 서비스에 있으며 누구나 유전자 그래프에서 비교 위치를 찾을 수 있습니다. 이를 위해서는 FTDNA(23andMe)의 상염색체 검사 결과만 있으면 충분하다. 연구 종료 시 지도가 제공됩니다. 지리적 분포 MDLP World-22 프로젝트의 주요 상염색체 구성요소에 대한 빈도 최대값.
아래 그래프는 각 모집단에 대한 주요 구성 요소와 평균 백분율을 보여줍니다. 한 줄에 - 한 모집단에 대한 백분율 분석. 각 구분(수직 막대)은 10%에 해당하며, 상염색체 성분의 이름은 범례에서 위에서 아래로 왼쪽에서 오른쪽으로 동일한 순서입니다. 공통 유전학의 백분율 구성이 유사할수록 다른 사람들, 위 그래프의 그림이 더 유사해 보입니다. 그럼 시작하겠습니다...
독일인, 리투아니아인, 러시아인, 스웨덴인, 핀란드인 등의 유전학
이 그래프는 유럽인의 주요 유전적 구성요소를 보여주며 다양한 인구에서 동유럽 구성요소(북동유럽)의 감소와 정렬됩니다. 보시다시피, 모든 유럽인들은 유전적 측면에서 매우 다르며, 동일한 기원의 집합에 유전적 구성 요소가 있지만 그럼에도 불구하고 매우 다른 비율입니다. 일반적으로 모든 슬라브인과 발트인에게 가장 중요한 것 중 하나는 리투아니아인과 벨로루시인 사이에서 최대인 동유럽의 이 구성 요소입니다. 아마도 고고학적 "유선 도자기 문화" 시대부터 이들 국가의 영토가 이 구성 요소의 기원 중심이었을 것입니다. 리투아니아인의 경우 80% 이상, 이탈리아인의 경우 20%에 불과합니다.보라색은 Atlanto-Mediterranean 구성 요소를 나타내며 북동쪽에서 남서쪽으로 이동함에 따라 증가합니다. 따라서 핀란드인들 사이에서는 평균 15%, 이탈리아인들 사이에서는 40%에 이릅니다. 나머지 구성 요소는 덜 발음됩니다.
러시아 우크라이나인 벨로루시인의 유전학

이 그래프는 동부 슬라브를 보여줍니다 - 러시아인, 벨로루시인, 우크라이나인. 나열된 세 민족의 유전적 패턴의 유사성에 주의를 기울이고 오차 범위 내에서 거의 차이가 없습니다. 우크라이나인과 남부 러시아인은 서아시아 구성 요소가 약간 증가하고 북부 러시아인은 조건부로 Samoyedic이라고 불리는 시베리아 구성 요소와 유럽의 Mesolithic 구성 요소가 약 10 %로 증가하여 후자의 지표에 따르면 독일어를 사용하는 스칸디나비아 인구 인 스웨덴에 더 가까워집니다.
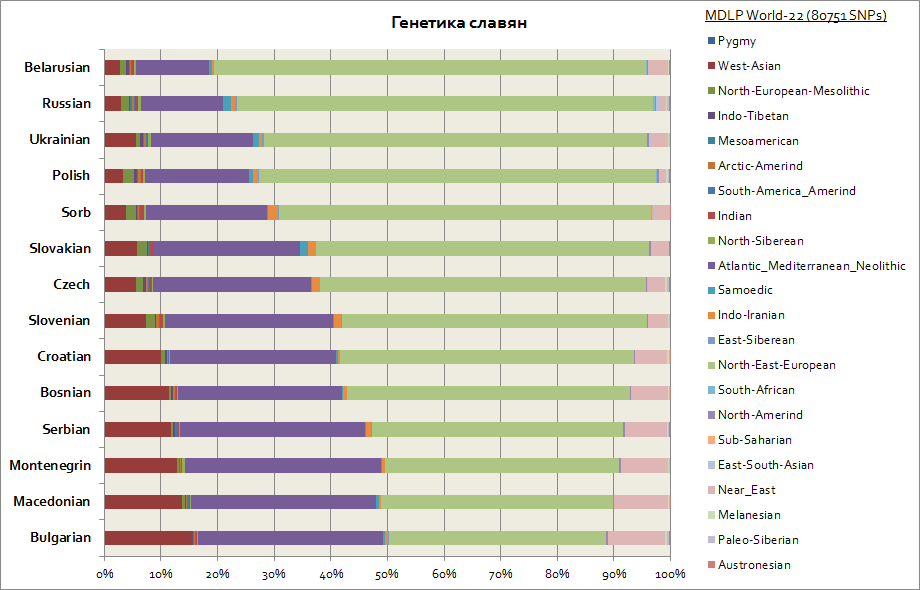
이 차트는 서부-폴란드인과 체코인, 남부인-세르비아인, 불가리아인, 마케도니아인 등을 포함한 모든 슬라브인을 묘사합니다.
모든 슬라브는 동유럽과 대서양-지중해의 두 가지 주요 구성 요소를 가지고 있습니다. 첫 번째는 벨로루시 인의 경우 최대이고 두 번째는 모든 남부 슬라브 인 - 세르비아 인, 마케도니아인, 불가리아 인입니다. 동유럽 구성요소는 슬라브인들 사이에서 더 주요한 기원이며, 아틀란토-지중해 구성요소는 슬라브인들이 발칸 반도로 이주함에 따라 더 많이 획득됩니다. 서부 우크라이나인과 슬로바키아인은 이웃에 비해 사모예딕 구성 요소가 약간 증가했습니다. 슬라브 민족- 벨로루시인, 체코인, 폴란드인; 이것은 아마도 Huns와 Ugrians가 중부 유럽으로 중세 이주한 유전 적 흔적 일 것입니다.
슬라브인, 러시아인, 타타르인, 독일인, 백인, 유대인 등의 유전학
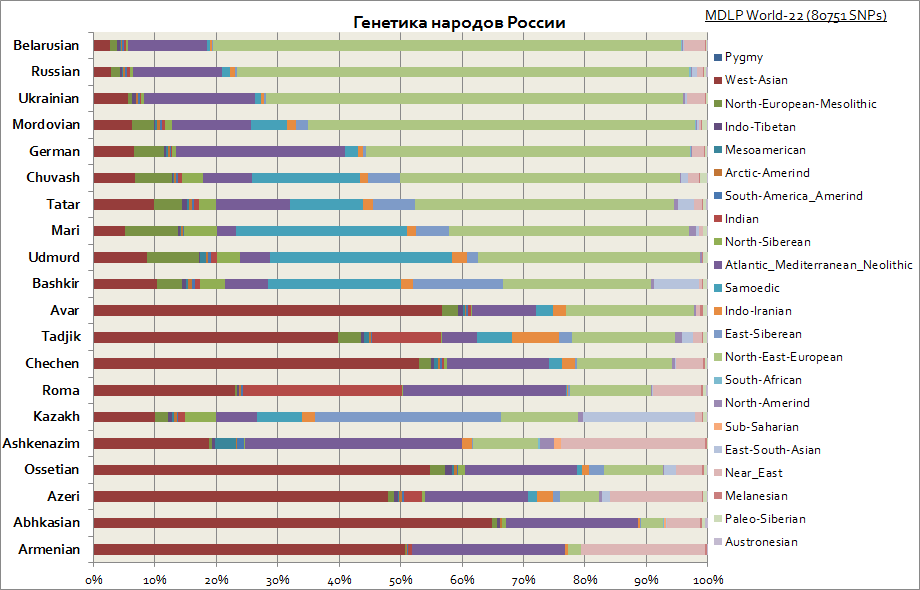
이 그래프는 러시아 민족의 다양한 기원을 보여줍니다. 볼 수 있듯이 슬라브 중 주요 구성 요소는 동유럽 구성 요소이며 볼가 지역 사람들 사이에서는 시베리아 구성 요소의 비율이 증가합니다. 백인의 경우 서아시아 구성 요소, 지중해 및 중동이 가장 특징적입니다.
Finns, Ugrians, Udmurts, 헝가리인, Saami 등의 유전학
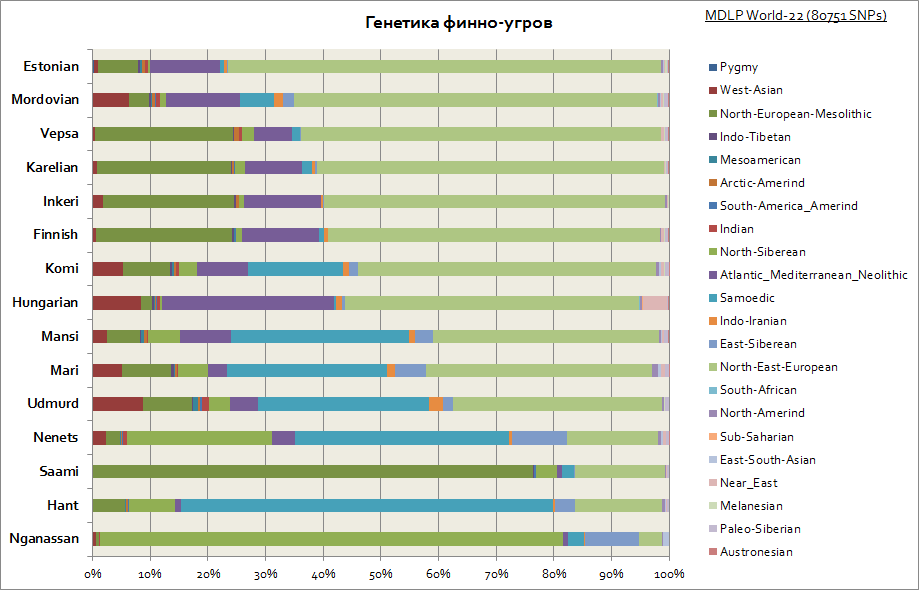
보시다시피 Finns, Vepsians 및 Karelians는 비슷한 특징이 있습니다. 유전적 기원슬라브와 함께. 그들은 또한 가장 큰 동유럽 구성 요소를 가지고 있으며, 이 지역의 시베리아 구성 요소가 증가함에 따라 Urals 및 Volga 지역에 가까워지면 감소합니다. 또한 모든 Finno-Ugric 민족은 유럽의 Mesolithic의 중요한 구성 요소를 가지고 있으며 Saami 중 거의 80 %에 도달하고 유럽의 인도 유럽 및 신석기 이전 인구와 관련이 있습니다. 카르파티아 지역과 중부 유럽의 다른 인구와 마찬가지로 전체 헝가리인에게 이러한 유전적 구성 요소 집합은 특징적입니다.
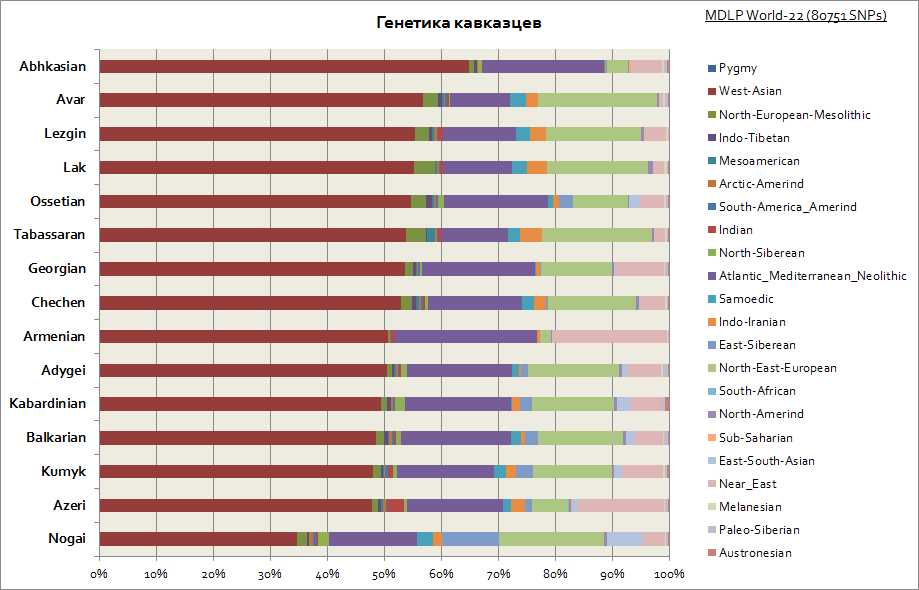
보시다시피 전체 코카서스는 상대적으로 유사한 유전 적 기원을 특징으로합니다. 이는 서아시아 구성 요소와 지중해의 상당 부분입니다. Nogais만이 약간 눈에 띕니다. 그들은 시베리아 구성 요소의 비율이 증가했습니다.

Ashkenazim과 Sephardim에서 볼 수 있듯이 서아시아, 대서양-지중해 및 중동 구성 요소의 빈도가 높습니다. 동시에 Ashkenazim은 아마도 Khazar 유산과 동유럽 구성 요소의 최대 30 % 증가로 인해 시베리아 구성 요소가 약간 증가했습니다. 남부 유럽의 국가.
특히 에티오피아 유대인과 인도 유대인만이 "회사"에서 제외됩니다. 전자는 사하라 사막 이남 아프리카의 비율이 높고(최대 40%) 후자는 조건부로 인도인이라고 불리는 남아시아 유전적 구성 요소의 비율이 높습니다(최대 50%).
Tatars, Bashkirs, Azerbaijanis, Chuvashs 등의 유전학
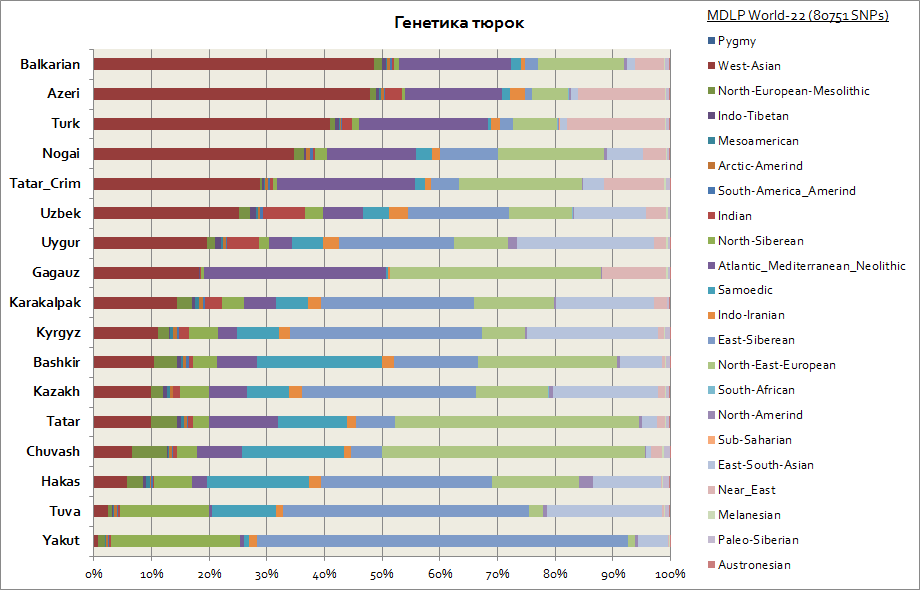
유전 적 측면에서 터키인은 유전 적 구성 요소가 크게 다르기 때문에 가장 이질적인 인종 그룹 중 하나로 밝혀졌습니다. 따라서 Turks의 주요 고향이 시베리아라는 점을 감안할 때 Yakuts, Tuvans, Khakasses와 같은 사람들은 동 시베리아 상 염색체 구성 요소를 30 ~ 65 %에 이르는 가장 큰 비율로 유지했습니다. 이 유전적 요소는 또한 키르기즈와 카자흐족의 주요 요소입니다. 나머지 구성 요소는 터키인을 거주 지역의 사람들에게 더 가깝게 만듭니다. 따라서 Yakuts 및 Tuvans의 경우 이들은 North Siberian 및 Samoyed 구성 요소입니다. 총 3개의 시베리아 구성 요소 Yakuts 중 최대 90%, Tuvans 중 최대 70%, 동남아시아 구성 요소의 20%로 증가하여 인구의 이주 흐름과 더 관련이 있습니다. 동아시아. Bashkirs의 경우 3개 시베리아 구성 요소의 비율은 최대 45%이며 동남아시아 구성 요소는 최대 10%입니다. Tatars는 평균 25~50%의 3가지 시베리아 유전적 구성 요소에 대한 데이터를 가지고 있습니다. 동시에 Bashkirs 중 Caucasoid 인구의 특징적인 구성 요소 비율은 최대 45 %, Tatars 중 평균 50 ~ 70 %입니다. 아제르바이잔인과 투르크인의 유전학은 오차 범위 내에서 실질적으로 다르지 않습니다. 평균 20%). 3개 시베리아 구성 요소의 비율은 아제르바이잔, 투르크, 발카르로 대표되며 3-7% 수준입니다.
결론
사람들의 유전학은 어족의 분포 또는 특정 인구에서 나타나는 편부모 마커(Y-DNA 및 mt-DNA 하플로그룹)의 비율과 직접적이고 중요한 상관관계가 없습니다. 가장 큰 상관 관계는 영토-지리적 원칙에 따라 추적할 수 있습니다. 따라서 일반적으로 특징적인 시베리아 구성 요소의 비율은 몽골로이드 종족동쪽에서 서쪽으로 점차 감소하고 그에 따라 코카서스 인종의 특징적인 구성 요소의 비율이 증가합니다. 우랄의 북쪽에서 라인을 따라 국경 지역에서 중앙 아시아대략적인 비율. 바이칼 호수 동쪽 지역에서는 큰 코카서스인종의 유전적 성분이 거의 나타나지 않는 반면, 페초라-볼가선 서쪽 지역에서는 큰 몽골인종의 시베리아인 성분이 사라지고 있다. .동유럽 유전 요소가 시베리아로 확산된 것은 이미 청동기 시대(안드로노프 서클의 문화)에 상당 부분 발생했지만, 축치 중 시베리아 극동의 개별 봉우리는 이미 러시아인의 이주와 관련이 있을 수 있습니다. 17 세기.
Negroid 종족의 특징 인 사하라 사막 이남 구성 요소의 몫은 아프리카 전역에 분포되어 있습니다. 지중해 남부와 아프리카 대륙의 북부 국경까지 적도 부분에서 최대에 도달하며 실제로는 외부에서 발견되지 않습니다. 밝은 배경은 아라비아 반도와 이란 고원의 남쪽 부분에 분포합니다.
유전적 요소의 지리학
알렉세이 조린
프로젝트