คนยุโรปคนใดที่เป็นคนรัสเซียใกล้ชิดทางพันธุกรรม พันธุศาสตร์ของชาวยูเครนรัสเซีย เบลารุสและตาตาร์ ชาวสลาฟและคอเคเชียน ชาวยิว ชาวฟินน์ และประชากรกลุ่มอื่นๆ
เพนตากอนยอมรับการทดลองกับวัสดุชีวภาพของพลเมืองรัสเซีย
ข้อสันนิษฐานเกี่ยวกับสันทรายเกี่ยวกับการพัฒนาอาวุธชีวภาพที่เป็นไปได้โดยชาวอเมริกันได้รับการยืนยันอย่างหนักแน่น เพนตากอนยอมรับความจริงของการรวบรวมวัสดุชีวภาพจากพลเมืองรัสเซีย
โบ ดาวนีย์ โฆษกเพนตากอนระบุว่า ศูนย์วิจัยโมเลกุลของกลุ่มแพทย์ทางอากาศที่ 59 ของกองทัพอากาศสหรัฐฯ กำลังทำการศึกษาเกี่ยวกับกระดูกและกล้ามเนื้อเพื่อระบุตัวบ่งชี้ทางชีวภาพต่างๆ ที่เกี่ยวข้องกับการบาดเจ็บ จำเป็นต้องใช้ตัวอย่างที่มาจากรัสเซียเท่านั้น เนื่องจากชุดแรกมาจากรัสเซีย และตอนนี้จำเป็นต้องใช้ตัวอย่างที่เหมือนกันเพื่อการควบคุม
จำได้ว่ากองทัพอากาศสหรัฐตั้งใจที่จะซื้อตัวอย่างโมเลกุล RNA 12 ตัวอย่างและของเหลวไขข้อ (ข้อต่อ) 27 ตัวอย่างของชาวรัสเซีย ประกาศดังกล่าวถูกโพสต์บนพอร์ทัลการจัดซื้อจัดจ้างของรัฐบาลสหรัฐฯ ในขณะเดียวกัน สัญญาเน้นย้ำว่าในบรรดาผู้ที่อาศัยอยู่ในรัสเซีย ลูกค้ามีความสนใจเฉพาะชาวยุโรปเท่านั้น และผู้อพยพ เช่น จากยูเครนจะไม่ได้รับการพิจารณา
ประเด็นของการรวบรวมวัสดุชีวภาพจากรัสเซียเริ่มมีการพูดคุยกันในสังคมหลังจากที่วลาดิเมียร์ปูตินแถลงต่อสาธารณะเกี่ยวกับเรื่องนี้ ตามที่เขาพูด วัสดุชีวภาพถูกรวบรวม "สำหรับกลุ่มชาติพันธุ์ต่างๆ และผู้คนที่อาศัยอยู่ในที่ต่างกัน จุดทางภูมิศาสตร์ สหพันธรัฐรัสเซีย". “คำถามคือทำไมพวกเขาถึงทำเช่นนี้” ประธานาธิบดีถามวาทศิลป์ในที่ประชุมกับนักเคลื่อนไหวด้านสิทธิมนุษยชน
ต่อมา Dmitry Peskov อธิบายว่าเป็นข้อมูลที่ได้รับผ่านบริการพิเศษ
คำพูดของปูตินกระตุ้นการเยาะเย้ยกัดกร่อนจากส่วนหนึ่งของสังคมรัสเซีย “ชาวอินคาโบราณก็กลัวกลอุบายสกปรกเช่นกัน ดังนั้นผู้ปกครองสูงสุดจึงมีสาวใช้พิเศษ ซึ่งมีหน้าที่กินผม น้ำลาย ตัดเล็บ และวัสดุชีวภาพอื่น ๆ ที่เหลือจากมหาราช เพื่อป้องกันไม่ให้ขยะตกลงไปอย่างไร้ความปรานี มือ” อาจารย์อันเดรย์อ้างจากสื่อ Nikulin
อย่างไรก็ตาม ประเทศกำลังเตรียมรับมือกับภัยคุกคามใหม่ในระดับกฎหมาย คาดว่าในเดือนธันวาคมกฎหมายว่าด้วยการคุ้มครองวัสดุชีวภาพจะถูกส่งไปยัง State Duma “วันนี้เรามีห้องปฏิบัติการหลายแห่งที่ทำการวิจัยทางคลินิกซึ่งเป็นห้องปฏิบัติการต่างประเทศ เช่น Invitro เราให้พวกเขาเข้าไปใกล้ชิดที่สุด” Gennady Onishchenko รองหัวหน้าคนแรกของ State Duma Committee on Education and Science อธิบายถึงปัญหา
ใน "Invitro" พวกเขาปฏิเสธความเกี่ยวข้องในการส่งออกวัสดุชีวภาพ ในทางกลับกันสถาบันพันธุศาสตร์ทั่วไปของ Russian Academy of Sciences "SP" รายงานว่าสถาบันนี้มีส่วนร่วมในการรวบรวมวัสดุชีวภาพ “ผู้อำนวยการด้านวิทยาศาสตร์ของสถาบันของเรา นักวิชาการ Nikolai Yankovsky เพิ่งจัดและเป็นหัวหน้าโครงการรวบรวมวัสดุชีวภาพ” Ilya Zakharov-Gezehus พนักงานของสถาบันกล่าว ไม่สามารถติดต่อ Yankovsky "SP" ได้ในทันที
อาวุโส นักวิจัยห้องปฏิบัติการพันธุศาสตร์ของสถาบันชีววิทยาพัฒนาการของ Russian Academy of Sciences Alexey Kulikov มีแนวโน้มที่จะพิสูจน์เพื่อนร่วมงานชาวอเมริกันของเขา
คุณต้องเข้าใจว่าคนอเมริกันกำลังทำอะไร พวกเขาพิจารณาว่ายีนทำงานอย่างไรในผู้ป่วยที่มีระบบกล้ามเนื้อและกระดูกได้รับผลกระทบ และดูที่องค์ประกอบของน้ำไขข้อ เริ่มแรกพวกเขาซื้อวัสดุชีวภาพที่ไหนสักแห่งในยุโรปตะวันออกจากตัวแทนผู้ป่วยของประเทศสลาฟ ดังนั้นพวกเขาจึงต้องการตัวอย่างควบคุมของคนที่ปกติดีกับระบบกล้ามเนื้อและกระดูกจากตัวแทนของประเทศสลาฟ: ชาวรัสเซีย ชาวยูเครน ชาวเบลารุส เป็นต้น
"SP": - ดีอย่างไร การประกวดราคาระบุไว้อย่างชัดเจนว่าวัสดุชีวภาพจากยูเครนจะไม่ได้รับการพิจารณา พวกเขาต้องการชาวรัสเซีย...
ยังไม่มีเจตนาใดๆทั้งสิ้น อาวุธทางพันธุกรรมล้วนเป็นเรื่องไร้สาระ สิ่งนี้ไม่สมจริงเพราะผู้คนในโลกมีความหลากหลายมากเกินไป - มีความหลากหลายมาก เป็นการยากที่จะคิดหาสิ่งที่ได้ผลสำหรับบางคนและไม่ได้ผลสำหรับคนอื่น ฉันคิดว่าที่นี่ เรากำลังพูดถึงเกี่ยวกับเงื่อนไขของการทดลอง กิน ประสบการณ์ทางวิทยาศาสตร์และมีการควบคุม วัสดุควบคุมต้องมาจากภูมิภาคเดียวกัน
"SP": - ทุกอย่างเป็นเช่นนั้นหาก "การศึกษาทางวิทยาศาสตร์" ไม่ได้ดำเนินการโดยกองทัพเพนตากอน ...
การวิจัยทางวิทยาศาสตร์สามารถทำได้โดยกองทัพ นอกจากนี้เรายังมีงานด้านการแพทย์เฉพาะที่แก้ไขโดยตัวแทนของหน่วยงานบังคับใช้กฎหมาย บางทีเรากำลังพูดถึงความคล่องตัวต่ำของนักบิน พวกเขาต้อง เป็นเวลานานนั่ง. ดังนั้นจึงเป็นสิ่งสำคัญโดยพื้นฐานที่จะไม่มีปัญหากับระบบกล้ามเนื้อและกระดูก ดังนั้นชาวอเมริกันจึงสนใจโรคเหล่านี้และยีนใดที่รับผิดชอบในเรื่องนี้
ในทางกลับกันหัวหน้าห้องปฏิบัติการของสถาบันพันธุศาสตร์ทั่วไปของ Russian Academy of Sciences, Sergey Kiselev เชื่อว่าการสร้างอาวุธทางพันธุกรรมนั้นเสี่ยงเกินไปและง่ายกว่าที่จะฆ่าคน วิธีการแบบดั้งเดิม.
มีการถ่ายโอนวัสดุชีวภาพจากรัสเซียและกำลังถ่ายโอน อย่างน้อยก็ตามที่ Onishchenko กล่าว ภายในกรอบของการทดลองทางคลินิก เนื่องจากในช่วง 25 ปีที่ผ่านมา บริษัทต่างชาติหลายสิบแห่งได้ทำการทดลองยาทางคลินิกในรัสเซีย จำเป็นต้องใช้วัสดุชีวภาพเพื่อทำความเข้าใจว่ายาทำงานอย่างไร
แน่นอนว่าตัวอย่างดังกล่าวสามารถใช้ได้ทั้งเพื่อวัตถุประสงค์ข้างต้นและเพื่อวัตถุประสงค์อื่นๆ เพราะมันยังคงเป็นตัวอย่างทางชีววิทยาของชาติ สิ่งสำคัญคือข้อมูลที่ได้รับจากตัวอย่างจะถูกนำมาใช้อย่างไร ข้อมูลทางพันธุกรรมสามารถนำมาใช้เพื่อวัตถุประสงค์ที่หลากหลาย รวมถึงเพื่อผลประโยชน์ส่วนบุคคล
"SP": - อะไรนะ?
เทคโนโลยีในปัจจุบันช่วยให้เราสามารถกำหนดจีโนมของแต่ละคนได้อย่างละเอียด นั่นคือการเก็บตัวอย่าง DNA จากช้อนในห้องอาหารของอาคาร CIA คุณสามารถระบุได้ว่าเจ้าหน้าที่ข่าวกรองมาจากรัฐมินนิโซตาจริง ๆ ตามที่เขียนไว้ตอนสมัครงานหรือมาจากไซบีเรียตะวันออก . นั่นคือคุณสามารถเชื่อมโยงบุคคลเข้ากับสถานที่ได้อย่างแม่นยำ
แต่สำหรับสิ่งนี้คุณต้องสร้างก่อน แผนที่พันธุกรรมดินแดน ฉันคิดว่าหน่วยข่าวกรองที่สนใจของหลายประเทศทั่วโลกได้สร้างแผนที่ดินแดนดังกล่าวมาเป็นเวลานาน เพื่อให้สามารถระบุตัวบุคคลใน สถานการณ์ที่แตกต่างกัน.
ตัวอย่างเช่นคนที่มีอาชีพที่เป็นอันตราย หากหนึ่งในนั้นถูกฉีกออกเป็นชิ้นๆ ก็เป็นไปได้โดยใช้แผนที่พันธุกรรมเพื่อระบุว่าเขาเป็นใคร มาจากไหน และแสดงความเคารพต่อเขา นั่นคือภายในประเทศ แต่ถ้าข้อมูลนี้ไปถึงบุคคลที่สามก็สามารถใช้เพื่อประโยชน์ของพวกเขาได้
"SP": - V กรณีนี้เราสนใจในความน่าจะเป็นของการสร้างอาวุธพันธุกรรมชาติพันธุ์ ...
ในความคิดของฉัน การสร้างอาวุธแบบนี้ไม่มีจุดหมาย ประการแรก ในการจะฆ่าคน จำเป็นต้องมีอิทธิพลต่อการทำงานที่สำคัญ เช่น การหายใจ การไหลเวียนโลหิต ฯลฯ มีไม่กี่อย่างและเหมือนกันสำหรับทุกคน การค้นหาหน้าที่สำคัญทางเชื้อชาตินั้นแทบจะเป็นไปไม่ได้เลย
และประการที่สอง ใครคือชาวอเมริกัน? พวกเขาไม่มีชาติที่มียศฐาบรรดาศักดิ์ ทุกคนปะปนกันที่นั่น รวมทั้งรัสเซีย จีน แองโกล-แซกซอน ดังนั้น หากมีการสร้างอาวุธเพื่อต่อต้านรัสเซีย ชาวอเมริกันบางส่วนจะตกอยู่ภายใต้การกระทำของมัน เนื่องจากพวกเขาเป็นชาวรัสเซียหรือไม่ก็ลูกหลานของพวกเขา
ดังนั้นยิ่งการแพร่กระจายของผู้คนทั่วโลก การผสมจีโนมมากเท่าไหร่ โอกาสที่จะสร้างอาวุธทางพันธุกรรมก็จะยิ่งน้อยลงเท่านั้น มันไม่เกิดประโยชน์ ง่ายกว่า ถูกกว่า และมีประสิทธิภาพมากกว่าในการ "แช่" ในอีกทางหนึ่ง
"SP": - อย่างไรก็ตามเพนตากอนดำเนินการศึกษาดังกล่าวและวันนี้ตัวแทนยอมรับโดยตรง ...
ฉันดูเงื่อนไขของการประกวดราคา ฐานทัพอากาศแห่งนี้ขอตัวอย่าง RNA แต่ RNA เป็นสารพันธุกรรมย่อย เป็นตัวกลางระหว่างจีโนมมนุษย์และสถานะการทำงานของเซลล์ RNA สะท้อนถึงความเชี่ยวชาญของแต่ละเซลล์ในคราวเดียวหรืออย่างอื่น นั่นคือจีโนม - ดีเอ็นเอในแต่ละเซลล์ของร่างกายจะเหมือนกันเสมอ และ RNA ในแต่ละเซลล์ก็แตกต่างกัน เพราะมันเป็นภาพจำลองการทำงานของจีโนมทุกนาที ดังนั้นตัวอย่าง RNA ที่ได้จากนิ้วหัวแม่เท้าจะแตกต่างจาก RNA จากอวัยวะอื่นๆ ชาวอเมริกันไม่ได้ระบุว่าพวกเขาต้องการ RNA ที่ไหน
สิ่งที่สองที่พวกเขาขอคือน้ำไขข้อจากแคปซูลข้อต่อ นอกจากนี้ เงื่อนไขการประกวดราคาระบุว่าของเหลวนั้นสามารถรับจากผู้บริจาครายอื่นได้ ไม่ใช่จากผู้ที่ได้รับ RNA นั่นคือชาวอเมริกันไม่ต้องการการเชื่อมต่อระหว่าง RNA และน้ำไขข้อ ดูเหมือนพวกเขาเอาตัวอย่างมาศึกษาคนละเรื่องกัน ไม่เกี่ยวกันเลย
"SP": - อย่างไรก็ตามสิ่งนี้ไม่ได้เพิ่มความชัดเจน ...
บางทีพวกเขาอาจฟอกเงินที่นั่น หรือนักเรียนได้เรียนรู้วิธีการเขียนประกวดราคา
แต่อดีตสมาชิกคณะกรรมาธิการว่าด้วยอาวุธชีวภาพแห่งสหประชาชาติ อิกอร์ นิคูลิน ไม่สงสัยใดๆ เกี่ยวกับแผนรุกของเพนตากอน
แน่นอนว่านี่เป็นความพยายามของชาวอเมริกันที่จะทำการวิจัยเพื่อจุดประสงค์ทางทหาร เพนตากอนไม่ใช่องค์กรการกุศลหรือเพื่อมนุษยธรรมที่ออกแบบมาเพื่อเป็นประโยชน์ต่อมวลมนุษยชาติ แต่ตรงกันข้าม ที่นี่เป็นไปได้ที่จะใช้ไวรัสที่จะทำหน้าที่คัดเลือก
ไวรัสดังกล่าวยังคงมีอยู่ในปัจจุบัน ตัวอย่างเช่น อีโบลา ลาสซา มาร์บวร์ก ไข้เลือดออกที่พวกเขาทำให้เกิดส่วนใหญ่ในเผ่าพันธุ์เนกรอยด์, ไข้หวัดนก - ในมองโกลอยด์, โรคซาร์สในอินโด - ยูโรเปียน
"SP": - ในเงื่อนไขของการแข่งขัน ด้วยเหตุผลบางประการ จึงมีข้อยกเว้นสำหรับยูเครน ...
ในยูเครน โครงการอเมริกันได้ดำเนินการเมื่อ 5-7 ปีที่แล้ว และตอนนี้พวกเขามีแต่โรคระบาด ไม่ว่าจะเป็นโรคหัด หรือหัดเยอรมัน หรือวัณโรค หรือบาดทะยัก หรืออหิวาตกโรค เป็นต้น จากนั้นชาวอเมริกันก็เสนอวัคซีนป้องกันสิ่งนี้ให้พวกเขา สะดวกสบายมาก
ในช่วง 10 ปีที่ผ่านมา สหรัฐฯ ใช้เงินหลายหมื่นล้านดอลลาร์ จัดตั้งห้องปฏิบัติการมากกว่า 400 แห่งทั่วโลก ซึ่งมีการพัฒนาอาวุธชีวภาพและวัคซีนประเภทใหม่ๆ มีห้องปฏิบัติการประมาณสี่สิบแห่งในประเทศต่างๆ อดีตสหภาพโซเวียต. ได้แก่ ยูเครน มอลโดวา จอร์เจีย อาร์เมเนีย อาเซอร์ไบจาน อุซเบกิสถาน คีร์กีซสถาน และคาซัคสถาน นั่นคือความโกลาหลที่ถูกควบคุม
"SP": - แต่นักพันธุศาสตร์ชาวรัสเซียของเราไม่เชื่อเกี่ยวกับการพัฒนาอาวุธที่มุ่งเป้าไปที่กลุ่มชาติพันธุ์ใดกลุ่มหนึ่ง ...
มีอะไรเหลือให้พวกเขาทำอีก? หลายคนอยู่ในทุน ถ้าฉันได้รับทุนฉันก็จะเงียบเช่นกัน แต่เนื่องจากชาวอเมริกันจะไม่เสนอทุนใดๆ ให้ฉัน ฉันจึงสามารถพูดได้อย่างอิสระในหัวข้อเหล่านี้
ให้คะแนนข่าว
ข่าวพันธมิตร:โดยธรรมชาติแล้ว รหัสพันธุกรรมของทุกคนจะถูกจัดเรียงในลักษณะที่แต่ละคนมีโครโมโซม 23 คู่ ซึ่งเก็บข้อมูลทางพันธุกรรมทั้งหมดที่สืบทอดมาจากทั้งพ่อและแม่
การก่อตัวของโครโมโซมเกิดขึ้นในช่วงเวลาของไมโอซิส เมื่อระหว่างการผสมข้ามโครโมโซมแต่ละอันสุ่มใช้เวลาครึ่งหนึ่งจากโครโมโซมของมารดาและครึ่งหนึ่งมาจากบิดา ซึ่งยีนเฉพาะจะได้รับมรดกจากแม่และใดจากพ่อ ไม่รู้สิ ทุกอย่างถูกกำหนดโดยบังเอิญ
โครโมโซมเพศชายเพียงโครโมโซม Y เดียวเท่านั้นที่ไม่เข้าร่วมในลอตเตอรีนี้ มันถ่ายทอดจากพ่อสู่ลูกโดยสิ้นเชิงเหมือนกระบองวิ่งผลัด ฉันจะชี้แจงว่าผู้หญิงไม่มีโครโมโซม Y นี้เลย
ในแต่ละรุ่นต่อมา การกลายพันธุ์เกิดขึ้นในบางบริเวณของโครโมโซม Y ซึ่งเรียกว่า loci ซึ่งจะถ่ายทอดไปยังรุ่นต่อๆ ไปทั้งหมด ผู้ชาย.
ต้องขอบคุณการกลายพันธุ์เหล่านี้ที่ทำให้สามารถสร้างสกุลขึ้นมาใหม่ได้ โครโมโซม Y มีเพียงประมาณ 400 ตำแหน่ง แต่มีเพียงประมาณร้อยตำแหน่งเท่านั้นที่ใช้สำหรับการวิเคราะห์แฮปโลไทป์เชิงเปรียบเทียบและการสร้างโครโมโซมใหม่
ในตำแหน่งที่เรียกว่า loci หรือเรียกอีกอย่างว่าเครื่องหมาย STR มีการทำซ้ำตีคู่ตั้งแต่ 7 ถึง 42 ครั้ง ภาพรวมซึ่งเป็นเอกลักษณ์เฉพาะของแต่ละคน หลังจากผ่านไปหลายชั่วอายุคน การกลายพันธุ์ก็เกิดขึ้นและจำนวนการเกิดซ้ำตามกันก็เปลี่ยนขึ้นหรือลง ดังนั้นจะเห็นได้บนต้นไม้ทั่วไปว่ายิ่งมีการกลายพันธุ์มากเท่าใด บรรพบุรุษร่วมของกลุ่มแฮปโลไทป์ก็จะยิ่งมีอายุมากขึ้นเท่านั้น
กลุ่มแฮ็ปโลเองไม่มีข้อมูลทางพันธุกรรมเพราะ ข้อมูลทางพันธุกรรมอยู่ในออโตโซม - โครโมโซม 22 คู่แรก คุณสามารถเห็นการกระจายขององค์ประกอบทางพันธุกรรมในยุโรป Haplogroups เป็นเพียงเครื่องหมายของอดีตในช่วงเช้าของการก่อตัวของคนสมัยใหม่
กลุ่มแฮ็ปโลกรุ๊ปใดที่พบได้บ่อยที่สุดในหมู่ชาวรัสเซีย
คน |
มนุษย์ |
||||||||
|---|---|---|---|---|---|---|---|---|---|
ชาวสลาฟตะวันออก ตะวันตก และใต้. |
|||||||||
| ชาวรัสเซีย(ทิศเหนือ) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | |
| ชาวรัสเซีย(ศูนย์) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | |
| ชาวรัสเซีย(ใต้) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 3 | |
| ชาวรัสเซีย (ทั้งหมดชาวรัสเซียผู้ยิ่งใหญ่) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| ชาวเบลารุส | 574 | 52 | 10 | 3 | 16 | 10 | 3 | รัสเซีย, สลาฟ, อินโด-ยูโรเปียน และแฮ็ปโลกรุ๊ป R1a, R1b, N1c, I1 และ I2ในสมัยโบราณเมื่อประมาณ 8-9 พันปีก่อน มีกลุ่มภาษาศาสตร์กลุ่มหนึ่งซึ่งวางรากฐานให้กับภาษาตระกูลอินโด-ยูโรเปียน (ใน ชั้นต้นเป็นไปได้มากว่าเหล่านี้คือ haplogroups R1a และ R1b) ครอบครัวอินโด - ยูโรเปียนรวมถึงกลุ่มภาษาเช่นอินโด - อิหร่าน (เอเชียใต้), สลาฟและบอลต์ ( ยุโรปตะวันออก), เคลต์ (ยุโรปตะวันตก), เยอรมัน (ภาคกลาง, ยุโรปเหนือ).บางทีพวกเขาอาจมีบรรพบุรุษทางพันธุกรรมร่วมกันซึ่งเมื่อประมาณ 7,000 ปีก่อนเนื่องจากการอพยพย้ายถิ่นฐานไปยังส่วนต่าง ๆ ของยูเรเซียบางส่วนไปทางใต้และตะวันออก (R1a-Z93) ซึ่งเป็นรากฐานสำหรับชนชาติอินโด - อิหร่านและ ภาษา (ส่วนใหญ่มีส่วนร่วมใน ethnogenesis ของชนชาติเตอร์ก) และส่วนหนึ่งยังคงอยู่ในดินแดนของยุโรปและวางรากฐานสำหรับการก่อตัวของชนชาติยุโรปจำนวนมาก (R1b-L51) รวมถึงชาวสลาฟและ ชาวรัสเซียโดยเฉพาะอย่างยิ่ง (R1a-Z283, R1b-L51) ในขั้นตอนต่าง ๆ ของการก่อตัวในสมัยโบราณมีจุดตัดของกระแสการอพยพซึ่งทำให้มีแฮ็ปโลกรุ๊ปจำนวนมากในกลุ่มชาติพันธุ์ยุโรปทั้งหมด ภาษาสลาฟเกิดขึ้นจากกลุ่มภาษา Balto-Slavic ที่เคยรวมเป็นหนึ่งเดียว (สันนิษฐานว่าเป็นวัฒนธรรมทางโบราณคดีของ Corded Ware ตอนปลาย) จากการคำนวณของนักภาษาศาสตร์ Starostin สิ่งนี้เกิดขึ้นเมื่อประมาณ 3.3 พันปีที่แล้ว ช่วงเวลาตั้งแต่ศตวรรษที่ 5 ก่อนคริสต์ศักราช ถึงพุทธศตวรรษที่ 4-5 สามารถพิจารณาได้ตามเงื่อนไข Proto-Slavic, tk ชาวบอลต์และชาวสลาฟได้แยกทางกันแล้ว แต่ยังไม่มีชาวสลาฟ พวกเขาจะปรากฏในภายหลังเล็กน้อยในศตวรรษที่ 4-6 ในระยะเริ่มต้นของการก่อตัวของชาวสลาฟประมาณ 80% เป็นแฮ็ปโลกรุ๊ป R1a-Z280 และ I2a-M423 ในช่วงเริ่มต้นของการก่อตัวของ Balts ประมาณ 80% เป็นแฮ็ปโลกรุ๊ป N1c-L1025 และ R1a-Z92 อิทธิพลและจุดตัดของการอพยพของชาวบอลต์และชาวสลาฟนั้นมาจากจุดเริ่มต้นเพราะในหลาย ๆ ด้านการแบ่งนี้เป็นไปตามอำเภอใจและโดยทั่วไปจะสะท้อนเฉพาะแนวโน้มหลักโดยไม่มีรายละเอียด ภาษาอิหร่านเป็นภาษาอินโด - ยูโรเปียนและการออกเดทมีดังนี้ - เก่าแก่ที่สุดตั้งแต่ 2 พันปีก่อนคริสต์ศักราช ถึงศตวรรษที่ 4 ก่อนคริสต์ศักราช คนกลาง - จากศตวรรษที่ 4 ก่อนคริสต์ศักราช ถึงคริสต์ศตวรรษที่ 9 และใหม่ - จากคริสต์ศตวรรษที่ 9 จนถึงตอนนี้. นั่นคือภาษาอิหร่านที่เก่าแก่ที่สุดปรากฏขึ้นหลังจากการจากไปของชนเผ่าที่พูดภาษาอินโด - ยูโรเปียนจากเอเชียกลางไปยังอินเดียและอิหร่าน กลุ่มแฮ็ปโลกรุ๊ปหลักของพวกเขาน่าจะเป็น R1a-Z93, J2a, G2a3 กลุ่มภาษาอิหร่านตะวันตกปรากฏขึ้นในภายหลังประมาณศตวรรษที่ 5 ก่อนคริสต์ศักราช ดังนั้นชาวอินโด-อารยัน, เคลต์, ชาวเยอรมันและชาวสลาฟในทางวิทยาศาสตร์วิชาการจึงกลายเป็นชาวอินโด-ยูโรเปียน คำนี้เหมาะสมที่สุดสำหรับพื้นที่กว้างใหญ่และ กลุ่มที่หลากหลาย. สิ่งนี้ถูกต้องอย่างแน่นอน ในด้านพันธุกรรม ความแตกต่างของสายพันธุ์อินโด-ยูโรเปียนนั้นโดดเด่นทั้งใน Y-haplogroups และในออโตโซม ชาวอินโด - อิหร่านมีลักษณะเฉพาะในระดับที่มากขึ้นจากอิทธิพลทางพันธุกรรมของเอเชียตะวันตกของ BMAC ตามคัมภีร์พระเวทของอินเดีย ชาวอินโด-อารยันที่เดินทางมาอินเดีย (เอเชียใต้) จากทางเหนือ (จากเอเชียกลาง) และเป็นเพลงสวดและตำนานของพวกเขาที่เป็นพื้นฐานของพระเวทอินเดีย และดำเนินการต่อไปเรามาสัมผัสภาษาศาสตร์กันเพราะภาษารัสเซียนี้ (และภาษาบอลติกที่เกี่ยวข้องเช่นภาษาลิทัวเนียซึ่งเป็นส่วนหนึ่งของชุมชนภาษาศาสตร์บัลโต - สลาฟที่มีอยู่เดิม) ค่อนข้างใกล้เคียงกับภาษาสันสกฤตพร้อมกับภาษาเซลติกภาษาเยอรมันและภาษาอื่น ๆ ของตระกูลอินโด-ยูโรเปียนขนาดใหญ่ แต่ในแผนพันธุกรรม ชาวอินโด-อารยันมีขอบเขตมากกว่าชาวเอเชียตะวันตกแล้ว ขณะที่พวกเขาเข้าใกล้อินเดีย อิทธิพลของเวดอยด์ก็ทวีความรุนแรงขึ้นเช่นกัน ก็เลยกลายเป็นว่า แฮปโลกรุ๊ป R1aในลำดับวงศ์ตระกูล DNA นี่คือกลุ่มแฮ็ปโลกรุ๊ปทั่วไปสำหรับส่วนหนึ่งของชาวสลาฟ ส่วนหนึ่งของเติร์ก และส่วนหนึ่งของอินโดอารยัน (เพราะโดยธรรมชาติมีตัวแทนของกลุ่มแฮ็ปโลกรุ๊ปอื่น ๆ ในสภาพแวดล้อมของพวกเขา) แฮ็ปโลกรุ๊ป R1a1ในระหว่างการอพยพไปตามที่ราบรัสเซียพวกเขากลายเป็นส่วนหนึ่งของชนชาติ Finno-Ugric เช่น Mordovians (Erzya และ Moksha) ส่วนหนึ่งของชนเผ่า (สำหรับ แฮ็ปโลกรุ๊ป R1a1นี่คือกลุ่มย่อยของ Z93) ในระหว่างการอพยพพวกเขานำภาษาอินโด - ยูโรเปียนนี้ไปยังอินเดียและอิหร่านเมื่อประมาณ 3,500 ปีที่แล้วนั่นคือในช่วงกลางของ 2 พันปีก่อนคริสต์ศักราช ในอินเดียโดยแรงงานของ Panini ผู้ยิ่งใหญ่มันถูกแปลงเป็นภาษาสันสกฤตในช่วงกลางของ 1 พันปีก่อนคริสต์ศักราชและในเปอร์เซีย - อิหร่านภาษาอารยันกลายเป็นพื้นฐานของกลุ่มภาษาอิหร่านซึ่งเก่าแก่ที่สุด ย้อนหลังไปถึง 2 พันปีก่อนคริสต์ศักราช ข้อมูลเหล่านี้ได้รับการยืนยัน: ลำดับวงศ์ตระกูลดีเอ็นเอและภาษาศาสตร์มีความสัมพันธ์กันที่นี่ ส่วนใหญ่ แฮ็ปโลกรุ๊ป R1a1-Z93แม้แต่ในสมัยโบราณพวกเขาเข้าร่วมกับกลุ่มชาติพันธุ์เตอร์กและปัจจุบันเป็นการอพยพของชาวเติร์กในหลาย ๆ ด้านซึ่งไม่น่าแปลกใจในสมัยโบราณ แฮ็ปโลกรุ๊ป R1a1ในขณะที่ตัวแทน แฮ็ปโลกรุ๊ป R1a1-Z280เป็นส่วนหนึ่งของชนเผ่า Finno-Ugric แต่ในระหว่างการตั้งถิ่นฐานของชาวอาณานิคมสลาฟพวกเขาหลายคนถูกหลอมรวมโดยชาวสลาฟ แต่ถึงตอนนี้ในหมู่ชนชาติจำนวนมากเช่น Erzya ยังคงเป็นแฮ็ปโลกรุ๊ปที่โดดเด่น R1a1-Z280. ข้อมูลใหม่ทั้งหมดนี้สามารถให้เราได้ ลำดับวงศ์ตระกูลดีเอ็นเอโดยเฉพาะอย่างยิ่งวันที่โดยประมาณของการอพยพของผู้ให้บริการแฮ็ปโลกรุ๊ปในดินแดนของที่ราบรัสเซียสมัยใหม่และเอเชียกลางในยุคก่อนประวัติศาสตร์ ดังนั้นนักวิทยาศาสตร์สำหรับชาวสลาฟ เคลต์ เยอรมัน ฯลฯ ทุกคน ให้ชื่ออินโด - ยูโรเปียนซึ่งเป็นความจริงจากมุมมองของภาษาศาสตร์ ชาวอินโด-ยูโรเปียนเหล่านี้มาจากไหน ในความเป็นจริงมีภาษาอินโด - ยูโรเปียนมานานก่อนที่จะมีการอพยพไปยังอินเดียและอิหร่านทั่วที่ราบรัสเซียและไกลถึงคาบสมุทรบอลข่านทางตอนใต้และไปไกลถึงเทือกเขาพิเรนีสทางตะวันตก ต่อมาภาษาได้แพร่กระจายไปยังเอเชียใต้ - ทั้งอิหร่านและอินเดีย แต่ในแง่พันธุกรรมความสัมพันธ์นั้นเล็กกว่ามาก “สิ่งเดียวที่สมเหตุสมผลและได้รับการยอมรับในทางวิทยาศาสตร์ในปัจจุบันคือการใช้คำว่า “อารยัน” เฉพาะกับชนเผ่าและผู้คนที่พูดภาษาอินโด-อิหร่านเท่านั้น” ดังนั้นกระแสอินโด - ยูโรเปียนจึงไปในทิศทางใด - ไปทางทิศตะวันตกไปยังยุโรปหรือในทางกลับกันไปทางทิศตะวันออก? ตามการประมาณการ ตระกูลภาษาอินโด-ยูโรเปียนมีอายุประมาณ 8500 ปี ยังไม่ได้กำหนดบ้านบรรพบุรุษของชาวอินโด - ยูโรเปียน แต่ตามรุ่นใดรุ่นหนึ่งอาจเป็นภูมิภาคทะเลดำ - ทางใต้หรือทางเหนือ อย่างที่เราทราบกันดีอยู่แล้วว่าในอินเดีย ภาษาอินโด-อารยันถูกนำมาใช้เมื่อประมาณ 3,500 ปีที่แล้ว โดยน่าจะมาจากดินแดนของเอเชียกลาง และชาวอารยันเองก็เป็นกลุ่มที่มีสายพันธุกรรม Y ต่างกัน เช่น R1a1-L657, G2a, J2a, J2b, H เป็นต้น Haplogroup R1a1 ในยุโรปตะวันตกและยุโรปใต้การวิเคราะห์ 67 marker haplotypes แฮ็ปโลกรุ๊ป R1a1จากประเทศในยุโรปทั้งหมดทำให้สามารถกำหนดเส้นทางการอพยพโดยประมาณของบรรพบุรุษ R1a1 ในทิศทางของยุโรปตะวันตกได้ และจากการคำนวณพบว่าเกือบทั่วทั้งยุโรป ตั้งแต่ไอซ์แลนด์ทางตอนเหนือไปจนถึงกรีซทางตอนใต้ บรรพบุรุษร่วมกันของกลุ่มแฮ็ปโลกรุ๊ป R1a1 คือกลุ่มหนึ่งเมื่อประมาณ 7,000 ปีก่อน!กล่าวอีกนัยหนึ่งลูกหลานเช่นการแข่งขันวิ่งผลัดส่งต่อ haplotype ของพวกเขาไปยังลูกหลานของพวกเขาจากรุ่นสู่รุ่นโดยแยกย้ายกันไปในกระบวนการอพยพจากสถานที่ทางประวัติศาสตร์เดียวกัน - ซึ่งน่าจะเป็นเทือกเขาอูราลหรือที่ราบลุ่มทะเลดำ . บนแผนที่สมัยใหม่ ประเทศเหล่านี้ส่วนใหญ่อยู่ในยุโรปตะวันออกและยุโรปกลาง - โปแลนด์ เบลารุส ยูเครน รัสเซีย แต่ช่วงของแฮ็ปโลไทป์ที่เก่าแก่กว่าของแฮ็ปโลกรุ๊ป R1a1นำไปสู่ตะวันออก - สู่ไซบีเรีย และอายุขัยของบรรพบุรุษคนแรกซึ่งระบุโดย haplotypes ที่เก่าแก่ที่สุดและกลายพันธุ์มากที่สุดคือ 7.5 พันปีก่อน ในสมัยนั้นไม่มีชาวสลาฟ ไม่มีชาวเยอรมัน ไม่มีชาวเคลต์ ยุโรปกลางและตะวันออกโปแลนด์ บรรพบุรุษร่วมกันของ R1a1 มีชีวิตอยู่ประมาณ 5,000 ปีที่แล้ว (ส่วนใหญ่เป็นกลุ่มย่อยของ R1a1-M458 และ Z280) สำหรับภาษารัสเซีย - ยูเครน - 4500 ปีที่แล้วซึ่งสอดคล้องกับความแม่นยำในการคำนวณและแม้ว่าสี่ชั่วอายุคนจะไม่แตกต่างกันสำหรับเงื่อนไขดังกล่าว ในโปแลนด์สมัยใหม่ แฮ็ปโลกรุ๊ป R1a1เฉลี่ยอยู่ที่ 56% และในบางพื้นที่สูงถึง 62% ส่วนที่เหลือส่วนใหญ่เป็นชาวยุโรปตะวันตก แฮ็ปโลกรุ๊ป R1b(12%), สแกนดิเนเวีย แฮ็ปโลกรุ๊ป I1(17%) และทะเลบอลติก แฮ็ปโลกรุ๊ป N1c1 (8%). ในสาธารณรัฐเช็กและสโลวาเกีย บรรพบุรุษ Proto-Slavic ร่วมกันมีชีวิตอยู่เมื่อ 4200 ปีที่แล้ว น้อยกว่ารัสเซียและยูเครนเพียงเล็กน้อย นั่นคือเรากำลังพูดถึงการตั้งถิ่นฐานในดินแดนของโปแลนด์ยุคใหม่ สาธารณรัฐเช็ก สโลวาเกีย ยูเครน เบลารุส รัสเซีย - ทั้งหมดนี้เกิดขึ้นภายในไม่กี่ชั่วอายุคน แต่มากกว่าสี่พันปีที่แล้ว ในทางโบราณคดี ความถูกต้องของการหาคู่นั้นเป็นเรื่องที่คิดไม่ถึง ในลูกหลานของเช็กและสโลวาเกีย แฮ็ปโลกรุ๊ป R1a1ประมาณ 40% ส่วนที่เหลือมีชาวยุโรปตะวันตกเป็นส่วนใหญ่ R1b(22-28%) สแกนดิเนเวีย I1และบอลข่าน แฮ็ปโลกรุ๊ป I2a(สะสม 18%) ในดินแดนของฮังการีสมัยใหม่ บรรพบุรุษร่วมกัน R1a1 มีชีวิตอยู่เมื่อ 5,000 ปีที่แล้ว ปัจจุบันมีลูกหลานของกลุ่มแฮ็ปโลกรุ๊ป R1a1 มากถึงหนึ่งในสี่ ส่วนที่เหลือส่วนใหญ่เป็นกลุ่มแฮ็ปโลกรุ๊ปยุโรปตะวันตก R1b (20%) และแฮ็ปโลกรุ๊ปสแกนดิเนเวีย I1 และบอลข่าน I2 ที่รวมกัน (รวม 26%) เนื่องจากชาวฮังกาเรียนพูดภาษาในกลุ่มภาษา Finno-Ugric กลุ่มแฮ็ปโลกรุ๊ปที่พบมากที่สุดคือ N1c1ในหลุมฝังศพของชาว Magyars ที่ร่ำรวยในฮังการีโบราณส่วนใหญ่พบซากศพของผู้ชายที่มีแฮ็ปโลกรุ๊ป N1c1ซึ่งเป็นผู้นำคนแรกของชนเผ่าที่มีส่วนร่วมในการก่อตั้งอาณาจักร ในลิทัวเนียและลัตเวีย บรรพบุรุษร่วมกันถูกสร้างขึ้นใหม่จนลึกถึง 4,800 ปี โดยทั่วไปมี subclade วันนี้ Z92, Z280 และ M458 ที่พบมากที่สุดในหมู่ชาวลิทัวเนียคือกลุ่มแฮ็ปโลกรุ๊ป N1c1 ของทะเลบอลติกถึง 47% โดยทั่วไป ลิทัวเนียและลัตเวียมีลักษณะเฉพาะตามกลุ่มย่อยทะเลบอลติกใต้ L1025 ของกลุ่มแฮ็ปโลกรุ๊ป N1c1 โดยทั่วไปสถานการณ์มีความชัดเจน ฉันจะเพิ่มเฉพาะในประเทศแถบยุโรป - ไอซ์แลนด์ เนเธอร์แลนด์ เดนมาร์ก สวิตเซอร์แลนด์ เบลเยียม ลิทัวเนีย ฝรั่งเศส อิตาลี โรมาเนีย แอลเบเนีย มอนเตเนโกร สโลวีเนีย โครเอเชีย สเปน กรีซ บัลแกเรีย มอลโดวา - บรรพบุรุษร่วมกันอาศัยอยู่ 5,000- 5500 ปีที่แล้ว เป็นไปไม่ได้ที่จะแม่นยำกว่านี้ นี่คือบรรพบุรุษร่วมกัน แฮปโลกรุ๊ป R1aสำหรับทุกประเทศที่ระบุไว้ บรรพบุรุษของชาวยุโรปร่วมกัน กล่าวคือ ไม่นับภูมิภาคบอลข่านที่แสดงไว้ด้านบน ซึ่งเป็นบ้านของบรรพบุรุษของชาวอินโด-ยูโรเปียนเมื่อประมาณ 7,500 ปีที่แล้ว ส่วนแบ่งของผู้ขนส่ง แฮ็ปโลกรุ๊ป R1a1ในประเทศต่อไปนี้แตกต่างกันไปจาก 4% ในฮอลแลนด์และอิตาลี 9% ในแอลเบเนีย 8-11% ในกรีซ (มากถึง 14% ในเทสซาโลนิกิ) 12-15% ในบัลแกเรียและเฮอร์เซโกวีนา 14-17% ในเดนมาร์กและ เซอร์เบีย 15-25% ในบอสเนียและมาซิโดเนีย 3% ในสวิตเซอร์แลนด์ 20% ในโรมาเนียและฮังการี 23% ในไอซ์แลนด์ 22-39% ในมอลโดวา 29-34% ในโครเอเชีย 30-37% ในสโลวีเนีย (16 % ในคาบสมุทรบอลข่านโดยรวม) และในเวลาเดียวกัน - 32-37% ในเอสโตเนีย 34-38% ในลิทัวเนีย 41% ในลัตเวีย 40% ในเบลารุส 45-54% ในยูเครน ในยุโรปตะวันออกของรัสเซีย แฮปโลกรุ๊ป R1aดังที่ฉันได้กล่าวไปแล้วโดยเฉลี่ย 47% เนื่องจากส่วนแบ่งที่สูงของทะเลบอลติก แฮ็ปโลกรุ๊ป N1c1ทางตอนเหนือและตะวันตกเฉียงเหนือของรัสเซีย แต่ในตอนใต้และตอนกลางของรัสเซีย ส่วนแบ่งของ subclades ที่แตกต่างกันของ haplogroup R1a ถึง 55% เติร์กและแฮ็ปโลกรุ๊ป R1a1Haplotypes ของบรรพบุรุษนั้นแตกต่างกันไปในทุกที่ ภูมิภาคต่าง ๆ มีลักษณะเฉพาะตามกลุ่มย่อยของตนเอง ชาวอัลไตและชาวเติร์กอื่น ๆ ก็มีกลุ่มแฮ็ปโลกรุ๊ป R1a1 สูงเช่นกัน Bashkirs ถึง 40% ของกลุ่มย่อย Z2123 นี่คือสายย่อยจาก Z93 และสามารถเรียกโดยทั่วไปว่า Turkic และไม่เกี่ยวข้องกับการอพยพของชาวอินโด - อิหร่านวันนี้เป็นจำนวนมาก แฮ็ปโลกรุ๊ป R1a1จะอยู่ในภูมิภาคซายาโน-อัลไต ท่ามกลางประชากรเตอร์กในเอเชียกลาง ในบรรดาคีร์กีซถึง 63% คุณไม่สามารถเรียกพวกเขาว่ารัสเซียหรืออิหร่านได้ ปรากฎว่าชื่อทั้งหมด แฮ็ปโลกรุ๊ป R1a1ชื่อเดียว - การพูดเกินจริงขั้นต้น อย่างน้อย แต่ส่วนใหญ่ - ความไม่รู้ Haplogroups ไม่ใช่กลุ่มชาติพันธุ์ความเกี่ยวข้องทางภาษาและชาติพันธุ์ของผู้ให้บริการไม่ได้บันทึกไว้ในพวกเขา Haplogroups ไม่มีความสัมพันธ์โดยตรงกับยีน ชาวเติร์กมีลักษณะเด่นโดยกลุ่มย่อยต่างๆ ของ Z93 แต่ในภูมิภาคโวลก้าก็มี R1a1-Z280 เช่นกัน ซึ่งอาจถ่ายโอนไปยังโวลก้าเติร์กจากแม่น้ำโวลก้าฟินน์ Haplogroup R1a1-Z93 ยังเป็นเรื่องปกติสำหรับชาวอาหรับที่มีความถี่ปานกลาง สำหรับชาวเลวี - กลุ่มย่อยของชาวยิวอาซเคนาซี (กลุ่มย่อย CTS6 ได้รับการยืนยันในช่วงหลัง) บรรทัดนี้ในช่วงแรก ๆ มีส่วนร่วมใน ethnogenesis ของชนชาติเหล่านี้ อาณาเขตของการกระจายเริ่มต้น แฮ็ปโลกรุ๊ป R1a1ในยุโรป - นี่อาจเป็นดินแดนของยุโรปตะวันออกและอาจเป็นที่ราบลุ่มทะเลดำ ก่อนหน้านั้นอาจอยู่ในเอเชีย เอเชียใต้หรือจีนเหนือ คอเคเซียน R1a1 haplotypesอาร์เมเนีย อายุของบรรพบุรุษร่วมกันของแฮ็ปโลกรุ๊ป R1a1- 6500 ปีที่แล้ว โดยทั่วไปยังเป็นกลุ่มย่อยของ R1a1-Z93 แม้ว่าจะมี R1a1-Z282 ด้วยเอเชียไมเนอร์ คาบสมุทรอนาโตเลีย ทางแยกประวัติศาสตร์ระหว่างตะวันออกกลาง ยุโรป และเอเชีย เป็นผู้สมัครคนแรกหรือคนที่สองสำหรับ "อินโด-ยูโรเปียนมาตุภูมิ" อย่างไรก็ตาม บรรพบุรุษร่วมกันของกลุ่มแฮ็ปโลกรุ๊ป R1a1 อาศัยอยู่ที่นั่นเมื่อประมาณ 6,500 ปีก่อน เป็นที่ชัดเจนว่าบ้านของบรรพบุรุษนี้ซึ่งตัดสินโดย haplotypes อาจอยู่ในอานาโตเลียหรือชาวอินโด - ยูโรเปียนดั้งเดิมเป็นพาหะ แฮ็ปโลกรุ๊ป R1b. แต่มีความเป็นไปได้สูงที่จะเป็นตัวแทนของบุคคลจากตุรกีในระดับต่ำในฐานข้อมูล haplotype ทั่วไป ดังนั้นทั้งชาวอาร์มีเนียและชาวอนาโตเลีย - พวกเขาทั้งหมดมีบรรพบุรุษเดียวกันหรือบรรพบุรุษอยู่ใกล้กันมากในเวลาหลายชั่วอายุคน - นี่คือกลุ่มย่อยของ Z93 และ Z282 * ควรสังเกตว่า 4,500 ปีก่อนบรรพบุรุษร่วมกันของกลุ่มแฮ็ปโล R1a1-Z93 ในอานาโตเลียเป็นข้อตกลงที่ดีกับเวลาของการปรากฏตัวของชาวฮิตไทต์ในเอเชียไมเนอร์ในไตรมาสสุดท้ายของสหัสวรรษที่ 3 ก่อนคริสต์ศักราช แม้ว่า R1a1-Z93 จำนวนมาก เส้นอาจปรากฏขึ้นที่นั่นหลังจากการอพยพของชาวเตอร์กไปยังคาบสมุทรในยุคของเรา อเล็กเซย์ ซอร์ริน *** | |
โครโมโซมเพศชายเพียงโครโมโซม Y เดียวเท่านั้นที่ไม่ได้เข้าร่วมในลอตเตอรีนี้ มันถ่ายทอดจากพ่อสู่ลูกได้อย่างสมบูรณ์เหมือนไม้กระบอง ฉันจะชี้แจงว่าผู้หญิงไม่มีโครโมโซม Y นี้เลย
ในแต่ละรุ่นต่อมา การกลายพันธุ์เกิดขึ้นในบางพื้นที่ของโครโมโซม Y ซึ่งเรียกว่า loci ซึ่งจะถ่ายทอดไปยังรุ่นต่อๆ ไปทั้งหมดโดยเพศชาย ต้องขอบคุณการกลายพันธุ์เหล่านี้ที่ทำให้สามารถสร้างสกุลขึ้นมาใหม่ได้ โครโมโซม Y มีเพียงประมาณ 1,000 ตำแหน่ง แต่มีเพียงมากกว่าร้อยตำแหน่งเท่านั้นที่ใช้สำหรับการวิเคราะห์เปรียบเทียบแฮปโลไทป์และการสร้างสกุลใหม่
ในตำแหน่งที่เรียกว่า loci หรือเรียกอีกอย่างว่าเครื่องหมาย STR มีการทำซ้ำตีคู่ตั้งแต่ 7 ถึง 42 ครั้ง รูปแบบโดยรวมจะไม่ซ้ำกันสำหรับแต่ละคน หลังจากผ่านไปหลายชั่วอายุคน การกลายพันธุ์ก็เกิดขึ้นและจำนวนการเกิดซ้ำตามกันก็เปลี่ยนขึ้นหรือลง ดังนั้นจะเห็นได้บนต้นไม้ทั่วไปว่ายิ่งมีการกลายพันธุ์มากเท่าใด บรรพบุรุษร่วมของกลุ่มแฮปโลไทป์ก็จะยิ่งมีอายุมากขึ้นเท่านั้น
กลุ่มแฮ็ปโลเองไม่มีข้อมูลทางพันธุกรรมเพราะ ข้อมูลทางพันธุกรรมอยู่ในออโตโซม - โครโมโซม 22 คู่แรก คุณสามารถเห็นการกระจายขององค์ประกอบทางพันธุกรรมในยุโรป Haplogroups เป็นเพียงเครื่องหมายของอดีตในช่วงเช้าของการก่อตัวของคนสมัยใหม่
กลุ่มแฮ็ปโลกรุ๊ปใดที่พบได้บ่อยที่สุดในหมู่ชาวรัสเซีย
| คน | จำนวน มนุษย์ | R1a1, | R1b1, | I1, | I2, | N1c1, | E1b1b1, | เจทู | จีทูเอ, |
|---|---|---|---|---|---|---|---|---|---|
| ชาวสลาฟตะวันออก ตะวันตก และใต้. | |||||||||
| ชาวรัสเซีย(ทิศเหนือ) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | 1 |
| ชาวรัสเซีย(ศูนย์) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | 1 |
| ชาวรัสเซีย(ใต้) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 4 | 3 |
| ชาวรัสเซีย (ทั้งหมดชาวรัสเซียผู้ยิ่งใหญ่) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| ชาวเบลารุส | 574 | 52 | 10 | 3 | 16 | 10 | 3 | 2 | 2 |
| ชาวยูเครน | 93 | 54 | 2 | 5 | 16 | 8 | 8 | 6 | 3 |
| ชาวรัสเซีย(ร่วมกับชาวยูเครนและชาวเบลารุส) | 1874 | 48 | 7 | 4 | 13 | 16 | 4 | 3 | 3 |
| เสา | 233 | 56 | 16 | 7 | 10 | 8 | 4 | 3 | 2 |
| สโลวาเกีย | 70 | 47 | 17 | 6 | 11 | 3 | 9 | 4 | 1 |
| เช็ก | 53 | 38 | 19 | 11 | 12 | 3 | 8 | 6 | 5 |
| ชาวสโลวีเนีย | 70 | 37 | 21 | 12 | 20 | 0 | 7 | 3 | 2 |
| ชาวโครเอเชีย | 108 | 24 | 10 | 6 | 39 | 1 | 10 | 6 | 2 |
| ชาวเซิร์บ | 113 | 16 | 11 | 6 | 29 | 1 | 20 | 7 | 1 |
| ชาวบัลแกเรีย | 89 | 15 | 11 | 5 | 20 | 0 | 21 | 11 | 5 |
| บอลต์, ฟินน์, เยอรมัน, กรีก ฯลฯ | |||||||||
| ลิทัวเนีย | 164 | 34 | 5 | 5 | 5 | 44 | 1 | 0 | 0 |
| ชาวลัตเวีย | 113 | 39 | 10 | 4 | 3 | 42 | 0 | 0 | 0 |
| ฟินน์ (ตะวันออก) | 306 | 6 | 3 | 19 | 0 | 71 | 0 | 0 | 0 |
| ฟินน์ (ตะวันตก) | 230 | 9 | 5 | 40 | 0 | 41 | 0 | 0 | 0 |
| ชาวสวีเดน | 160 | 16 | 24 | 36 | 3 | 11 | 3 | 3 | 1 |
| ชาวเยอรมัน | 98 | 8 | 48 | 25 | 0 | 1 | 5 | 4 | 3 |
| ชาวเยอรมัน (บาวาเรีย) | 80 | 15 | 48 | 16 | 4 | 0 | 8 | 6 | 5 |
| ภาษาอังกฤษ | 172 | 5 | 67 | 14 | 6 | 0.1 | 3 | 3 | 1 |
| ไอริช | 257 | 1 | 81 | 6 | 5 | 0 | 2 | 1 | 1 |
| ชาวอิตาเลียน | 99 | 2 | 44 | 3 | 4 | 0 | 13 | 18 | 8 |
| ชาวโรมาเนีย | 45 | 20 | 18 | 2 | 18 | 0 | 7 | 13 | 7 |
| ชาวออสเซเชียน | 359 | 1 | 7 | 0 | 0 | 1 | 16 | 67 | |
| อาร์เมเนีย | 112 | 2 | 26 | 0 | 4 | 0 | 6 | 20 | 10 |
| ชาวกรีก | 116 | 4 | 14 | 3 | 10 | 0 | 21 | 23 | 5 |
| เติร์ก | 103 | 7 | 17 | 1 | 5 | 4 | 10 | 24 | 12 |
สิ่งที่น่าสังเกตเป็นพิเศษคือ haplogroups ที่พบมากที่สุด 4 กลุ่มในหมู่ชาวรัสเซีย:
R1a1 47.0%, N1c1 20.0%, I2 10.6%, I1 6.2%
การพูด ในแง่ง่ายๆ: องค์ประกอบทางพันธุกรรม ชาวรัสเซียตามแนวเส้นตรงของโครโมโซม Y ของเพศชายมีดังนี้
ชาวยุโรปตะวันออก - 47%
ทะเลบอลติก - 20%
และแฮ็ปโลกรุ๊ปสองกลุ่มของชาวยุโรปดั้งเดิมตั้งแต่ยุคหิน
สแกนดิเนเวีย - 6%
คาบสมุทรบอลข่าน - 11%
ชื่อมีเงื่อนไขและกำหนดตามขอบเขตสูงสุด ยุโรปคลาสย่อยสำหรับแฮ็ปโลกรุ๊ป R1a1, N1c1, I1 และ I2 ประเด็นพื้นฐานคือไม่มีลูกหลานของชาวมองโกลหลังจากแอกตาตาร์-มองโกลสองร้อยปี ทั้งสองเหลืออยู่ แต่มีทายาททางพันธุกรรมโดยตรงจำนวนน้อยมากจากความสัมพันธ์ดังกล่าว ด้วยคำเหล่านี้ฉันไม่ต้องการตั้งคำถาม แหล่งประวัติศาสตร์เกี่ยวกับชาวมองโกลในมาตุภูมิ แต่เพียงให้ความสนใจกับอิทธิพลทางพันธุกรรมที่คาดคะเนจากชาวมองโกล - ตาตาร์ที่มีต่อชาวรัสเซีย - มันไม่ได้อยู่ที่นั่นหรือไม่มีนัยสำคัญ นอกจากนี้ยังมีพาหะจำนวนมากในจีโนมของ Bulgar Tatars ฮาโปรกรุ๊ป R1a1(ประมาณ 30%) และ N1c1(ประมาณ 20%) แต่ส่วนใหญ่ไม่ใช่ชาวยุโรป
อื่น จุดสำคัญ, ชาวรัสเซียตอนใต้ไม่แตกต่างจากชาวยูเครนในระยะขอบของความผิดพลาด และชาวรัสเซียตอนเหนือที่มีกลุ่มแฮ็ปโลกรุ๊ป R1a1 ที่โดดเด่นก็มีเปอร์เซ็นต์ของกลุ่มแฮ็ปโล N1c1 ที่สูงขึ้นเช่นกัน แต่เปอร์เซ็นต์ของ N1c1 haplotypes นั้นอยู่ที่ 20% โดยเฉลี่ยในรัสเซีย
จักรพรรดิ นิโคลัส 2
บรรพบุรุษคนแรกที่เป็นที่รู้จักของ Grand Ducal House of Oldenburg คือ Egilmar เคานต์แห่ง Lerigau (d. 1108) ซึ่งกล่าวถึงในพงศาวดารสำหรับปี 1091
Nicholas II กลายเป็นพาหะของกลุ่มแฮ็ปโลกรุ๊ป R1b1a2- ตัวแทนของสายยุโรปตะวันตกจากราชวงศ์ Holstein-Gottorp ราชวงศ์เยอรมันนี้โดดเด่นด้วยเทอร์มินัลสนิป U106 ซึ่งพบมากที่สุดใน ยุโรปตะวันตกเฉียงเหนือในสถานที่ตั้งถิ่นฐานของชนเผ่าดั้งเดิม ไม่ใช่เรื่องปกติอย่างแน่นอน คนรัสเซียเครื่องหมายดีเอ็นเอ แต่การมีอยู่ในหมู่ชาวรัสเซียอาจเกี่ยวข้องกับการติดต่อระหว่างชาวเยอรมันและชาวสลาฟในระยะแรก
เจ้าชายตามธรรมชาติ รุริโควิชิ
Vladimir Monomakh และลูกหลานของเขาที่เรียกว่า "Monomachis" อยู่ในแฮ็ปโลกรุ๊ป N1c1-L550ซึ่งแพร่หลายในภูมิภาคบอลติกใต้ (กลุ่มย่อย L1025) และใน Fennoscandia (กลุ่มย่อย Y7795, Y9454, Y17113, Y17415, Y4338) ราชวงศ์ Rurik โดดเด่นด้วยเทอร์มินัลสนิป Y10931
บางคนที่นักประวัติศาสตร์เรียกว่า Olgoviches (ตั้งชื่อตาม Oleg Svyatoslavich - คู่ต่อสู้หลักของ Vladimir Monomakh ในการต่อสู้เกี่ยวกับระบบศักดินา - และตามที่แหล่งข่าวทั้งหมดยืนยันว่าลูกพี่ลูกน้องของเขา) ไม่เกี่ยวข้องกับ Rurikovichs จากตระกูล Monomashich (ในผู้ชายโดยตรง เส้น). นี่คือลูกหลานของ Yuri Tarussky
รัสเซีย, สลาฟ, อินโด-ยูโรเปียน และแฮ็ปโลกรุ๊ป R1a, R1b, N1c, I1 และ I2
ในสมัยโบราณเมื่อประมาณ 8-9 พันปีที่แล้ว มีกลุ่มภาษาศาสตร์กลุ่มหนึ่งที่วางรากฐานสำหรับตระกูลภาษาอินโด-ยูโรเปียน ครอบครัวอินโด - ยูโรเปียนรวมถึงกลุ่มภาษาเช่นอินโด - อิหร่าน (เอเชียใต้), สลาฟและบอลต์ (ยุโรปตะวันออก), เคลต์ (ยุโรปตะวันตก), เยอรมัน (กลาง, ยุโรปเหนือ) บางทีพวกเขาอาจมีบรรพบุรุษทางพันธุกรรมร่วมกันซึ่งเมื่อประมาณ 7,000 ปีก่อนเนื่องจากการอพยพย้ายถิ่นฐานไปยังส่วนต่าง ๆ ของยูเรเซียบางส่วนไปทางใต้และตะวันออก (R1a-Z93) ซึ่งเป็นรากฐานสำหรับชนชาติอินโด - อิหร่านและ ภาษา (ส่วนใหญ่มีส่วนร่วมใน ethnogenesis ของชนชาติเตอร์ก) และส่วนหนึ่งยังคงอยู่ในดินแดนของยุโรปและวางรากฐานสำหรับการก่อตัวของชนชาติยุโรปจำนวนมาก (R1b-L51) รวมถึงชาวสลาฟและ ชาวรัสเซียโดยเฉพาะอย่างยิ่ง (R1a-Z283, R1b-L51) ในขั้นตอนต่าง ๆ ของการก่อตัวในสมัยโบราณมีจุดตัดของกระแสการอพยพซึ่งทำให้มีแฮ็ปโลกรุ๊ปจำนวนมากในกลุ่มชาติพันธุ์ยุโรปทั้งหมดภาษาสลาฟเกิดขึ้นจากกลุ่มภาษา Balto-Slavic ที่เคยรวมเป็นหนึ่งเดียว (สันนิษฐานว่าเป็นวัฒนธรรมทางโบราณคดีของ Corded Ware ตอนปลาย) จากการคำนวณของนักภาษาศาสตร์ Starostin สิ่งนี้เกิดขึ้นเมื่อประมาณ 3.3 พันปีที่แล้ว ช่วงเวลาตั้งแต่ศตวรรษที่ 5 ก่อนคริสต์ศักราช ถึงพุทธศตวรรษที่ 4-5 สามารถพิจารณาได้ตามเงื่อนไข Proto-Slavic, tk ชาวบอลต์และชาวสลาฟได้แยกทางกันแล้ว แต่ยังไม่มีชาวสลาฟ พวกเขาจะปรากฏในภายหลังเล็กน้อยในศตวรรษที่ 4-6 ในระยะเริ่มต้นของการก่อตัวของชาวสลาฟประมาณ 80% เป็นแฮ็ปโลกรุ๊ป R1a-Z280 และ I2a-M423 ในช่วงเริ่มต้นของการก่อตัวของ Balts ประมาณ 80% เป็นแฮ็ปโลกรุ๊ป N1c-L1025 และ R1a-Z92 อิทธิพลและจุดตัดของการอพยพของชาวบอลต์และชาวสลาฟนั้นมาจากจุดเริ่มต้นเพราะในหลาย ๆ ด้านการแบ่งนี้เป็นไปตามอำเภอใจและโดยทั่วไปจะสะท้อนเฉพาะแนวโน้มหลักโดยไม่มีรายละเอียด
ภาษาอิหร่านเป็นภาษาอินโด - ยูโรเปียนและการออกเดทมีดังนี้ - เก่าแก่ที่สุดตั้งแต่ 2 พันปีก่อนคริสต์ศักราช ถึงศตวรรษที่ 4 ก่อนคริสต์ศักราช คนกลาง - จากศตวรรษที่ 4 ก่อนคริสต์ศักราช ถึงคริสต์ศตวรรษที่ 9 และใหม่ - จากคริสต์ศตวรรษที่ 9 จนถึงตอนนี้. นั่นคือภาษาอิหร่านที่เก่าแก่ที่สุดปรากฏขึ้นหลังจากการจากไปของชนเผ่าที่พูดภาษาอินโด - ยูโรเปียนจากเอเชียกลางไปยังอินเดียและอิหร่าน กลุ่มแฮ็ปโลกรุ๊ปหลักของพวกเขาน่าจะเป็น R1a-Z93, J2a, G2a3 กลุ่มภาษาอิหร่านตะวันตกปรากฏขึ้นในภายหลังประมาณศตวรรษที่ 5 ก่อนคริสต์ศักราช
ดังนั้นชาวอินโด - อารยัน, เคลต์, ชาวเยอรมันและชาวสลาฟในวิทยาศาสตร์การศึกษาจึงกลายเป็นชาวอินโด - ยูโรเปียน คำนี้เพียงพอที่สุดสำหรับกลุ่มที่กว้างใหญ่และหลากหลายเช่นนี้ สิ่งนี้ถูกต้องอย่างแน่นอน ในด้านพันธุกรรม ความแตกต่างของสายพันธุ์อินโด-ยูโรเปียนนั้นโดดเด่นทั้งใน Y-haplogroups และในออโตโซม ชาวอินโด - อิหร่านมีลักษณะเฉพาะในระดับที่มากขึ้นจากอิทธิพลทางพันธุกรรมของเอเชียตะวันตกของ BMAC
ตามคัมภีร์พระเวทของอินเดีย ชาวอินโด-อารยันที่เดินทางมาอินเดีย (เอเชียใต้) จากทางเหนือ (จากเอเชียกลาง) และเป็นเพลงสวดและตำนานของพวกเขาที่เป็นพื้นฐานของพระเวทอินเดีย และดำเนินการต่อไปเรามาสัมผัสภาษาศาสตร์กันเพราะภาษารัสเซียนี้ (และภาษาบอลติกที่เกี่ยวข้องเช่นภาษาลิทัวเนียซึ่งเป็นส่วนหนึ่งของชุมชนภาษาศาสตร์บัลโต - สลาฟที่มีอยู่เดิม) ค่อนข้างใกล้เคียงกับภาษาสันสกฤตพร้อมกับภาษาเซลติกภาษาเยอรมันและภาษาอื่น ๆ ของตระกูลอินโด-ยูโรเปียนขนาดใหญ่ แต่ในแผนพันธุกรรม ชาวอินโด-อารยันมีขอบเขตมากกว่าชาวเอเชียตะวันตกแล้ว ขณะที่พวกเขาเข้าใกล้อินเดีย อิทธิพลของเวดอยด์ก็ทวีความรุนแรงขึ้นเช่นกัน
ก็เลยกลายเป็นว่า แฮปโลกรุ๊ป R1aในลำดับวงศ์ตระกูล DNA นี่คือกลุ่มแฮ็ปโลกรุ๊ปทั่วไปสำหรับส่วนหนึ่งของชาวสลาฟ ส่วนหนึ่งของเติร์ก และส่วนหนึ่งของอินโดอารยัน (เพราะโดยธรรมชาติมีตัวแทนของกลุ่มแฮ็ปโลกรุ๊ปอื่น ๆ ในสภาพแวดล้อมของพวกเขา) แฮ็ปโลกรุ๊ป R1a1ในระหว่างการอพยพไปตามที่ราบรัสเซียพวกเขากลายเป็นส่วนหนึ่งของชนชาติ Finno-Ugric เช่น Mordovians (Erzya และ Moksha) ส่วนหนึ่งของชนเผ่า (สำหรับ แฮ็ปโลกรุ๊ป R1a1นี่คือกลุ่มย่อยของ Z93) ในระหว่างการอพยพพวกเขานำภาษาอินโด - ยูโรเปียนนี้ไปยังอินเดียและอิหร่านเมื่อประมาณ 3,500 ปีที่แล้วนั่นคือในช่วงกลางของ 2 พันปีก่อนคริสต์ศักราช ในอินเดียโดยแรงงานของ Panini ผู้ยิ่งใหญ่มันถูกแปลงเป็นภาษาสันสกฤตในช่วงกลางของ 1 พันปีก่อนคริสต์ศักราชและในเปอร์เซีย - อิหร่านภาษาอารยันกลายเป็นพื้นฐานของกลุ่มภาษาอิหร่านซึ่งเก่าแก่ที่สุด ย้อนหลังไปถึง 2 พันปีก่อนคริสต์ศักราช ข้อมูลเหล่านี้ได้รับการยืนยัน: ลำดับวงศ์ตระกูลดีเอ็นเอและภาษาศาสตร์มีความสัมพันธ์กันที่นี่
ส่วนใหญ่ แฮ็ปโลกรุ๊ป R1a1-Z93แม้แต่ในสมัยโบราณพวกเขาเข้าร่วมกับกลุ่มชาติพันธุ์เตอร์กและปัจจุบันเป็นการอพยพของชาวเติร์กในหลาย ๆ ด้านซึ่งไม่น่าแปลกใจในสมัยโบราณ แฮ็ปโลกรุ๊ป R1a1ในขณะที่ตัวแทน แฮ็ปโลกรุ๊ป R1a1-Z280เป็นส่วนหนึ่งของชนเผ่า Finno-Ugric แต่ในระหว่างการตั้งถิ่นฐานของชาวอาณานิคมสลาฟพวกเขาหลายคนถูกหลอมรวมโดยชาวสลาฟ แต่ถึงตอนนี้ในหมู่ชนชาติจำนวนมากเช่น Erzya ยังคงเป็นแฮ็ปโลกรุ๊ปที่โดดเด่น R1a1-Z280.
ข้อมูลใหม่ทั้งหมดนี้สามารถให้เราได้ ลำดับวงศ์ตระกูลดีเอ็นเอโดยเฉพาะอย่างยิ่งวันที่โดยประมาณของการอพยพของผู้ให้บริการแฮ็ปโลกรุ๊ปในดินแดนของที่ราบรัสเซียสมัยใหม่และเอเชียกลางในยุคก่อนประวัติศาสตร์
ดังนั้นนักวิทยาศาสตร์สำหรับชาวสลาฟ เคลต์ เยอรมัน ฯลฯ ทุกคน ให้ชื่ออินโด - ยูโรเปียนซึ่งเป็นความจริงจากมุมมองของภาษาศาสตร์
ชาวอินโด-ยูโรเปียนเหล่านี้มาจากไหน ในความเป็นจริงมีภาษาอินโด - ยูโรเปียนมานานก่อนที่จะมีการอพยพไปยังอินเดียและอิหร่านทั่วที่ราบรัสเซียและไกลถึงคาบสมุทรบอลข่านทางตอนใต้และไปไกลถึงเทือกเขาพิเรนีสทางตะวันตก ต่อมาภาษาได้แพร่กระจายไปยังเอเชียใต้ - ทั้งอิหร่านและอินเดีย แต่ในแง่พันธุกรรมความสัมพันธ์นั้นเล็กกว่ามาก
“สิ่งเดียวที่สมเหตุสมผลและได้รับการยอมรับในทางวิทยาศาสตร์ในปัจจุบันคือการใช้คำว่า “อารยัน” เฉพาะกับชนเผ่าและผู้คนที่พูดภาษาอินโด-อิหร่านเท่านั้น”
ดังนั้นกระแสอินโด - ยูโรเปียนจึงไปในทิศทางใด - ไปทางทิศตะวันตกไปยังยุโรปหรือในทางกลับกันไปทางทิศตะวันออก? ตามการประมาณการ ตระกูลภาษาอินโด-ยูโรเปียนมีอายุประมาณ 8500 ปี ยังไม่ได้กำหนดบ้านบรรพบุรุษของชาวอินโด - ยูโรเปียน แต่ตามรุ่นใดรุ่นหนึ่งอาจเป็นภูมิภาคทะเลดำ - ทางใต้หรือทางเหนือ อย่างที่เราทราบกันดีอยู่แล้วว่าในอินเดีย ภาษาอินโด-อารยันถูกนำมาใช้เมื่อประมาณ 3,500 ปีที่แล้ว โดยน่าจะมาจากดินแดนของเอเชียกลาง และชาวอารยันเองก็เป็นกลุ่มที่มีสายพันธุกรรม Y ต่างกัน เช่น R1a1-L657, G2a, J2a, J2b, H เป็นต้น
Haplogroup R1a1 ในยุโรปตะวันตกและยุโรปใต้
การวิเคราะห์ 67 marker haplotypes แฮ็ปโลกรุ๊ป R1a1จากประเทศในยุโรปทั้งหมดทำให้สามารถกำหนดเส้นทางการอพยพโดยประมาณของบรรพบุรุษ R1a1 ในทิศทางของยุโรปตะวันตกได้ และจากการคำนวณพบว่าเกือบทั่วทั้งยุโรป ตั้งแต่ไอซ์แลนด์ทางตอนเหนือไปจนถึงกรีซทางตอนใต้ บรรพบุรุษร่วมกันของกลุ่มแฮ็ปโลกรุ๊ป R1a1 คือกลุ่มหนึ่งเมื่อประมาณ 7,000 ปีก่อน! กล่าวอีกนัยหนึ่งลูกหลานเช่นการแข่งขันวิ่งผลัดส่งต่อ haplotype ของพวกเขาไปยังลูกหลานของพวกเขาจากรุ่นสู่รุ่นโดยแยกย้ายกันไปในกระบวนการอพยพจากสถานที่ทางประวัติศาสตร์เดียวกัน - ซึ่งน่าจะเป็นเทือกเขาอูราลหรือที่ราบลุ่มทะเลดำ . บนแผนที่สมัยใหม่ ประเทศเหล่านี้ส่วนใหญ่อยู่ในยุโรปตะวันออกและยุโรปกลาง - โปแลนด์ เบลารุส ยูเครน รัสเซีย แต่ช่วงของแฮ็ปโลไทป์ที่เก่าแก่กว่าของแฮ็ปโลกรุ๊ป R1a1นำไปสู่ตะวันออก - สู่ไซบีเรีย และอายุขัยของบรรพบุรุษคนแรกซึ่งระบุโดย haplotypes ที่เก่าแก่ที่สุดและกลายพันธุ์มากที่สุดคือ 7.5 พันปีก่อน ในสมัยนั้นไม่มีชาวสลาฟ ไม่มีชาวเยอรมัน ไม่มีชาวเคลต์ข้อเสียของวิธีการ
ถ้าเจ้าทำการทดสอบได้และเจ้าพอใจมาก เราก็จะรีบนำทัพพีน้ำมันดินเข้ามา ใช่ โครโมโซม Y ถ่ายทอดจากพ่อสู่ลูกโดยแทบไม่เปลี่ยนแปลง แต่จริงๆ แล้วเป็นลักษณะทางพันธุกรรม ข้อมูลที่เป็นประโยชน์ไม่ได้อยู่ในโครโมโซมคู่อื่น ๆ มียีนมากกว่านั้นมาก
และอีก 22 ตัวที่เหลือจะถูกสับแบบสุ่ม โดยไม่มีร่องรอยของการผสมดังกล่าวเหลืออยู่บน Y
จินตนาการ. ลูกเรือชาวแองโกลแซกซอนยึดรัฐนิโกร ผู้หญิงไม่ได้เดินทางเช่นนี้และคุณต้องติดต่อกับประชากรในท้องถิ่น มีตัวเลือกอะไรบ้าง?
1) แองโกล-แซกซอนมีลูกจากผู้หญิงผิวดำ แต่โอนสัญชาติให้เด็กผู้ชายเท่านั้น ในกรณีนี้ โครโมโซม Y จะถูกถ่ายโอนไปยังโครโมโซมยุโรป แต่สัดส่วนของยีนยุโรปที่มีนัยสำคัญจริงๆ จะลดลง รุ่นแรกจะเป็นคนผิวดำครึ่งหนึ่งและอดีต "ขุนนาง" ในกรณีเช่นนี้จะสลายไปอย่างรวดเร็วแม้ว่า Y จะมาจากสิ่งนี้ กลุ่มชาติพันธุ์. มันจะไม่สมเหตุสมผลมากนัก บางทีสิ่งที่คล้ายกันอาจเกิดขึ้นกับชาวฟินน์และชาวอินเดียนแดง ชาวยาคุตและฟินน์มีเปอร์เซ็นต์สูงสุดของลักษณะแฮ็ปโลกรุ๊ป N1c1 ของพวกเขา แต่โดยพันธุกรรมแล้ว คนเหล่านี้เป็นชนชาติที่แตกต่างกันอย่างสิ้นเชิงโดยมีกลุ่มย่อยที่แตกต่างกันของแฮ็ปโลกรุ๊ป N1c1 ที่มีประวัติอันเป็นเอกลักษณ์ของตนเอง ซึ่งแยกจากกันเมื่อกว่า 6 พันปีที่แล้ว และในทางกลับกัน ชาวอินเดีย - มีเปอร์เซ็นต์สูง แฮ็ปโลกรุ๊ป R1a1พันธุกรรมมีความเหมือนกันน้อยมากกับตัวแทนชาวยุโรปของกลุ่มแฮ็ปโลกรุ๊ปนี้ tk นอกจากนี้ยังมี subclades ต่างๆ ที่มีประวัติของตัวเองซึ่งแยกออกจากกันเมื่อกว่า 6 พันปีที่แล้ว
2) ชาวอินโดอารยันมีความพึงพอใจ ระบบวรรณะ. รุ่นแรกจะเป็นกึ่งนิโกร แต่ถ้าชนชั้นสูงผสมพันธุ์กันเองเท่านั้น เปอร์เซ็นต์ของพันธุกรรมดั้งเดิมจะลอยอยู่ประมาณ 50% แต่ในทางปฏิบัติแล้ว การแต่งงานกับผู้หญิงในท้องถิ่นเป็นส่วนใหญ่ และยิ่งกว่านั้นอีก ดังนั้นจึงเป็นไปไม่ได้เลยที่จะได้ยีนดั้งเดิมของผู้พิชิต และสิ่งนี้เกิดขึ้นในประวัติศาสตร์ของโลก วรรณะบนชาวอินเดียจาก 20% เป็น 72% มี แฮ็ปโลกรุ๊ป R1a1(โดยเฉลี่ย 43%) แต่โดยพันธุกรรมแล้วพวกเขามีความเหมือนกันน้อยมากกับตัวแทนของยุโรปหรือเตอร์กในสิ่งเดียวกัน แฮ็ปโลกรุ๊ป R1a1และเหตุผลอีกครั้งก็คือ subclades ต่างๆ ที่มีประวัติพิเศษของตัวเอง
สถานการณ์ที่คล้ายกันนี้อาจเกิดขึ้นในแคเมอรูน ซึ่งเป็นประเทศในแอฟริกากลางที่ Y มีมากถึง 95% แฮ็ปโลกรุ๊ป R1b-V88 แต่ในขณะเดียวกันก็อยู่ในกลุ่มประชากรแอฟริกันเนกรอยด์ทั่วไปทางมานุษยวิทยา
สรุปได้ว่าการมีเครื่องหมายและแฮ็ปโลกรุ๊ปเป็นเงื่อนไขสำคัญในการกำหนดสัญชาติ แต่ยังไม่เพียงพอ ในการระบุชาติกำเนิดของบุคคลนั้น Family Tree DNA มีการทดสอบ autosomal ที่เรียกว่า Family Finder
อเล็กเซย์ ซอร์ริน 
เมื่อเร็วๆ นี้ นักวิทยาศาสตร์ใกล้จะถอดรหัสรหัสพันธุกรรมของมนุษย์ได้แล้ว ในหลาย ๆ ด้านสิ่งนี้ทำให้สามารถดูประวัติศาสตร์ของ ethnos รัสเซียได้ใหม่ซึ่งกลายเป็นสิ่งที่เก่าแก่กว่าและไม่เป็นเนื้อเดียวกันอย่างที่คิดไว้ก่อนหน้านี้
ในส่วนลึกของศตวรรษ
จีโนมมนุษย์เป็นสิ่งที่เปลี่ยนแปลงได้ ในช่วงวิวัฒนาการของมนุษยชาติ กลุ่มแฮ็ปโลกรุ๊ปได้ผ่านการกลายพันธุ์มากกว่าหนึ่งครั้ง วันนี้นักวิทยาศาสตร์ได้เรียนรู้ที่จะกำหนดเวลาโดยประมาณเมื่อสิ่งนี้หรือการกลายพันธุ์เกิดขึ้น ดังนั้น, นักพันธุศาสตร์ชาวอเมริกันพบว่าการกลายพันธุ์ครั้งหนึ่งเกิดขึ้นเมื่อประมาณ 4,500 ปีที่แล้วบนที่ราบรัสเซียตอนกลาง เด็กชายคนหนึ่งเกิดมาซึ่งมีชุดนิวคลีโอไทด์ที่แตกต่างจากพ่อของเขา - เขาได้รับการจัดประเภททางพันธุกรรม R1a1 ซึ่งเกิดขึ้นแทน R1a ของพ่อ
การกลายพันธุ์นี้ไม่เหมือนคนอื่น ๆ ได้รับการพิสูจน์แล้วว่าเป็นไปได้ สกุล R1a1 ไม่เพียงรอดชีวิตมาได้ แต่ยังตั้งรกรากอยู่ในส่วนสำคัญของทวีปยูเรเซียอีกด้วย ปัจจุบัน ประมาณ 70% ของประชากรชายในรัสเซีย เบลารุส และยูเครน เป็นพาหะของกลุ่มแฮ็ปโลกรุ๊ป R1a1 และในเมืองเก่าของรัสเซีย จำนวนนี้สูงถึง 80% ดังนั้น R1a1 จึงทำหน้าที่เป็นเครื่องหมายของกลุ่มชาติพันธุ์รัสเซีย ปรากฎว่าเลือดของเด็กชายโบราณที่อาศัยอยู่ในยุคหินใหม่ตอนปลายไหลในเส้นเลือดของผู้ชายส่วนใหญ่ในรัสเซียยุคใหม่
ประมาณ 500 ปีหลังจากการกำเนิดของกลุ่มแฮ็ปโลกรุ๊ป R1a1 กระแสการอพยพของตัวแทนแพร่กระจายไปทางทิศตะวันออก - เลยเทือกเขาอูราลไปทางใต้ - ไปยังฮินดูสถานและไปทางทิศตะวันตก - ไปยังดินแดนสมัยใหม่ ประเทศในยุโรป. ความจริงที่ว่าผู้อาศัยในที่ราบรัสเซียตอนกลางไปไกลเกินกว่าพื้นที่ดั้งเดิมของพวกเขาก็ได้รับการยืนยันจากนักโบราณคดีเช่นกัน การวิเคราะห์ซากกระดูกของการฝังศพในอัลไตของ 1 พันปีก่อนคริสต์ศักราช อี แสดงให้เห็นว่านอกจากชาวมองโกลอยด์แล้วชาวคอเคเชียนที่เด่นชัดก็อาศัยอยู่ที่นั่นด้วย
ไม่มีตาตาร์
ในประเด็นหนึ่งของสิ่งพิมพ์วิทยาศาสตร์ยอดนิยม The American Journal of Human Genetics มีการตีพิมพ์บทความเกี่ยวกับการวิจัยโดยทีมนักวิทยาศาสตร์รัสเซีย - เอสโตเนียของกลุ่มยีนของชาวรัสเซีย การค้นพบของนักวิจัยเป็นสิ่งที่คาดไม่ถึง ประการแรก: Ethnos ของรัสเซียนั้นต่างกันโดยธรรมชาติทางพันธุกรรม ชาวรัสเซียส่วนหนึ่งที่อาศัยอยู่ในภาคกลางและภาคใต้ของประเทศอยู่ใกล้กับชนชาติสลาฟที่อยู่ใกล้เคียง ส่วนอื่น ๆ - ทางตอนเหนือของรัสเซีย - มีความสัมพันธ์อย่างใกล้ชิดทางพันธุกรรมกับชนชาติ Finno-Ugric
ข้อสรุปต่อไปน่าสนใจยิ่งขึ้น นักวิทยาศาสตร์ไม่สามารถตรวจพบองค์ประกอบเอเชียที่ฉาวโฉ่ในจีโนมของรัสเซียได้ ไม่มีชุดยีนตาตาร์-มองโกเลียในจำนวนที่สังเกตได้ในประชากรรัสเซีย ปรากฎว่าการแสดงออกที่มั่นคง "Scratch a Russian - คุณจะพบ Tatar" นั้นผิดพลาด
ศาสตราจารย์ Oleg Balanovsky หัวหน้าห้องปฏิบัติการภูมิศาสตร์จีโนมที่สถาบันพันธุศาสตร์ทั่วไปของ Russian Academy of Sciences พิจารณากลุ่มยีนของรัสเซีย "เกือบจะเป็นยุโรปทั้งหมด" และเรียกความแตกต่างจากเอเชียกลางว่า "ยอดเยี่ยมจริงๆ" เช่น ถ้าพวกเขาเป็นสองโลกที่แตกต่างกัน
นักวิชาการ Konstantin Skryabin หัวหน้าฝ่ายทิศทางจีโนมของสถาบัน Kurchatov เห็นด้วยกับ Balanovsky เขากล่าวว่า: "เราไม่พบการแนะนำตาตาร์ที่เห็นได้ชัดเจนในจีโนมของรัสเซีย ซึ่งหักล้างทฤษฎีเกี่ยวกับอิทธิพลการทำลายล้างของแอกมองโกล" นอกจากนี้นักวิทยาศาสตร์กล่าวว่าไซบีเรียนยังมีพันธุกรรมเหมือนกับ Old Believers - พวกเขามี "จีโนมรัสเซีย" ที่เหมือนกัน
นักวิจัยยังให้ความสนใจกับความแตกต่างเล็กน้อยในจีโนไทป์ระหว่างชาวรัสเซียในด้านหนึ่งกับชนชาติสลาฟที่อยู่ใกล้เคียง เช่น ชาวยูเครน ชาวเบลารุส และชาวโปแลนด์ ความแตกต่างระหว่างชาวสลาฟทางใต้และตะวันตกจากชาวรัสเซียเหนือนั้นชัดเจนกว่า
เครื่องหมายพิเศษ
ตามที่นักมานุษยวิทยา Vasily Deryabin กล่าวว่า จีโนไทป์ของรัสเซียก็มีเครื่องหมายทางสรีรวิทยาที่ชัดเจนเช่นกัน หนึ่งในนั้นคือความเด่นของดวงตาสีอ่อนในรัสเซีย: เทา, น้ำเงิน, เทา - น้ำเงิน, น้ำเงิน เรามี 45 เปอร์เซ็นต์ของพวกเขาใน ยุโรปตะวันตกน้อยกว่า - ประมาณ 35 เปอร์เซ็นต์ หลายคนในหมู่ชาวรัสเซียและมีผมสีขาว ตามที่นักมานุษยวิทยาชาวรัสเซียที่มีผมสีดำธรรมชาติไม่เกิน 5 เปอร์เซ็นต์ ในยุโรปตะวันตก โอกาสที่จะเจอคนผมดำคือ 45%
ตรงกันข้ามกับความเชื่อที่นิยมในหมู่ชาวรัสเซียมีคนจมูกดูแคลนไม่มาก - ประมาณ 7% ในประมาณ 75% ของกรณีจมูกตรง นอกจากนี้ยังไม่พบ epicanthus ในหมู่ชาวรัสเซียซึ่งเป็นแบบฉบับของตัวแทนของชนชาติมองโกลอยด์ที่มุมด้านในของดวงตา
กลุ่มชาติพันธุ์รัสเซียมีลักษณะเด่นของกลุ่มเลือด I และ II ในหมู่ชาวยิวเช่นกลุ่ม IV เป็นเรื่องธรรมดามาก การศึกษาทางชีวเคมียังแสดงให้เห็นว่าในเลือดของชาวรัสเซียรวมถึงชาวยุโรปอื่น ๆ มียีนพิเศษ PH-c แต่ไม่มีอยู่ใน Mongoloids
ชาวเหนือมีความใกล้ชิด
สถาบันวิจัยอณูพันธุศาสตร์ของ Russian Academy of Sciences และสถาบันมานุษยวิทยา ดี.เอ็น. Anuchin Moscow State University ได้ทำการศึกษาเชิงลึกเกี่ยวกับกลุ่มยีนของชาวรัสเซียในระหว่างที่มีการสร้างความแตกต่างในจีโนไทป์ระหว่างชาวรัสเซียและเพื่อนบ้านทางตอนเหนือของเราคือ Finns ซึ่งมีจำนวนถึงสามสิบหน่วย แต่ความแตกต่างทางพันธุกรรมระหว่างกลุ่มชาติพันธุ์รัสเซียกับชนชาติ Finno-Ugric (Mordovians, Mari, Veps, Karelians, Komi-Zyryans, Izhors) ซึ่งอาศัยอยู่ตามประเพณีทางตอนเหนือของประเทศของเรานั้นมีเพียงสามหน่วยเท่านั้น
นักวิทยาศาสตร์กำลังพูดถึงเอกภาพทางพันธุกรรมของชาวรัสเซียกับชาว Finno-Ugric แต่เกี่ยวกับต้นกำเนิดร่วมกัน ยิ่งไปกว่านั้น โครงสร้างเฉพาะของโครโมโซม Y ของกลุ่มชาติพันธุ์เหล่านี้ส่วนใหญ่เหมือนกับชาวฮินดูสถาน แต่ก็ไม่น่าแปลกใจเนื่องจากทิศทางของการตั้งถิ่นฐานของบรรพบุรุษทางพันธุกรรมของชาวรัสเซีย
กลุ่มแฮ็ปโลกรุ๊ปของมนุษย์ถูกส่งผ่านสายตรงระหว่างชายและหญิง แต่สำหรับพันธุกรรมของทั้งชายและหญิง ข้อมูลที่เก็บไว้ในออโตโซมของ DNA มีหน้าที่รับผิดชอบ ออโตโซมเป็นโครโมโซมของมนุษย์ 22 คู่แรกที่ถ่ายทอดลงมาจากทั้งพ่อและแม่หลังจากการข้ามผ่าน ซึ่งเป็นกระบวนการของการรวมตัวกันอีกครั้ง ดังนั้นข้อมูลทางพันธุกรรมประมาณครึ่งหนึ่งจึงถ่ายทอดจากพ่อและแม่ไปยังลูกหลานใน การศึกษาครั้งนี้มีการใช้ autosomal snips มากกว่า 80,000 จุด ความน่าเชื่อถือนั้นสูงมาก ความละเอียดสูงซึ่งทำให้สามารถจับอิทธิพลแม้เพียงเล็กน้อยในระดับพันธุกรรมในกลุ่มคนจำนวนมากได้ ข้อมูลการวิเคราะห์เปรียบเทียบนำมาจากการศึกษาแบบเปิดโดย V.Verenich ผู้เชี่ยวชาญใน การวิเคราะห์เปรียบเทียบองค์ประกอบทางพันธุกรรม เครื่องคำนวณทางพันธุกรรมอยู่ในบริการ GedMatch และอนุญาตให้ทุกคนค้นหาตำแหน่งเปรียบเทียบบนกราฟพันธุกรรม ในการทำเช่นนี้ ก็เพียงพอที่จะมีผลการทดสอบ autosomal จาก FTDNA หรือ 23andMe แผนที่มีให้เมื่อสิ้นสุดการศึกษา การกระจายทางภูมิศาสตร์และความถี่สูงสุดสำหรับส่วนประกอบ autosomal ที่สำคัญจากโครงการ MDLP World-22
กราฟด้านล่างแสดงองค์ประกอบหลักและเปอร์เซ็นต์เฉลี่ยของประชากรแต่ละกลุ่ม ในหนึ่งบรรทัด - การแบ่งเปอร์เซ็นต์สำหรับหนึ่งประชากร แต่ละส่วน (แถบแนวตั้ง) สอดคล้องกับ 10% และชื่อของส่วนประกอบ autosomal อยู่ในลำดับเดียวกันจากซ้ายไปขวาตามคำอธิบายจากบนลงล่าง ยิ่งองค์ประกอบร้อยละของพันธุศาสตร์ทั่วไปมีความคล้ายคลึงกันมากเท่าใด คนที่แตกต่างกันยิ่งตัวเลขในกราฟด้านบนดูคล้ายกันมากเท่าไหร่ มาเริ่มกันเลย...
พันธุศาสตร์ของชาวเยอรมัน ลิทัวเนีย รัสเซีย สวีเดน ฟินน์ ฯลฯ
กราฟนี้แสดงองค์ประกอบทางพันธุกรรมหลักสำหรับชาวยุโรปและสอดคล้องกับการลดลงขององค์ประกอบในยุโรปตะวันออก (ยุโรปตะวันออกเฉียงเหนือ) ในประชากรต่างๆ อย่างที่คุณเห็น คนยุโรปทุกคนมีความแตกต่างกันค่อนข้างมากในแง่ของพันธุกรรม และการมีองค์ประกอบทางพันธุกรรมในชุดต้นกำเนิดเดียวกัน พวกเขาก็มีเปอร์เซ็นต์ที่แตกต่างกันมาก สำหรับชาวสลาฟและบอลต์โดยทั่วไป หนึ่งในองค์ประกอบที่สำคัญที่สุดคือองค์ประกอบนี้ของยุโรปตะวันออก ซึ่งสูงสุดในหมู่ชาวลิทัวเนียและเบลารุส อาจมาจากช่วงเวลาของโบราณคดี "วัฒนธรรม Corded Ware" ดินแดนของประเทศเหล่านี้เป็นศูนย์กลางของแหล่งกำเนิดของส่วนประกอบนี้ มีตัวแทนมากกว่า 80% ในหมู่ชาวลิทัวเนียและเพียง 20% ในหมู่ชาวอิตาลีสีม่วงหมายถึงส่วนประกอบของแอตแลนโต-เมดิเตอร์เรเนียน และจะเพิ่มขึ้นเมื่อเคลื่อนตัวจากตะวันออกเฉียงเหนือไปทางตะวันตกเฉียงใต้ ดังนั้นในหมู่ชาวฟินน์จึงมีค่าเฉลี่ยถึง 15% และในหมู่ชาวอิตาลี 40% ส่วนประกอบที่เหลือมีความเด่นชัดน้อยกว่า
พันธุศาสตร์ของรัสเซีย Ukrainians เบลารุส

กราฟนี้แสดงชาวสลาฟตะวันออก - ชาวรัสเซีย ชาวเบลารุส ชาวยูเครน. ความสนใจถูกดึงดูดไปที่ความคล้ายคลึงกันของรูปแบบทางพันธุกรรมของทั้งสามคนที่อยู่ในรายการและภายในขอบเขตของข้อผิดพลาดพวกเขาแตกต่างกันน้อยมาก - ชาวยูเครนและรัสเซียตอนใต้มีองค์ประกอบในเอเชียตะวันตกเพิ่มขึ้นเล็กน้อยและรัสเซียทางเหนือเพิ่มขึ้นเล็กน้อยในหนึ่งเดียว ของส่วนประกอบไซบีเรียที่เรียกว่า Samoyedic อย่างมีเงื่อนไขและส่วนประกอบที่เพิ่มขึ้นของหินของยุโรปเป็นประมาณ 10% ซึ่งตามตัวบ่งชี้หลังทำให้พวกเขาใกล้ชิดกับประชากรสแกนดิเนเวียที่พูดภาษาเยอรมัน - ชาวสวีเดน
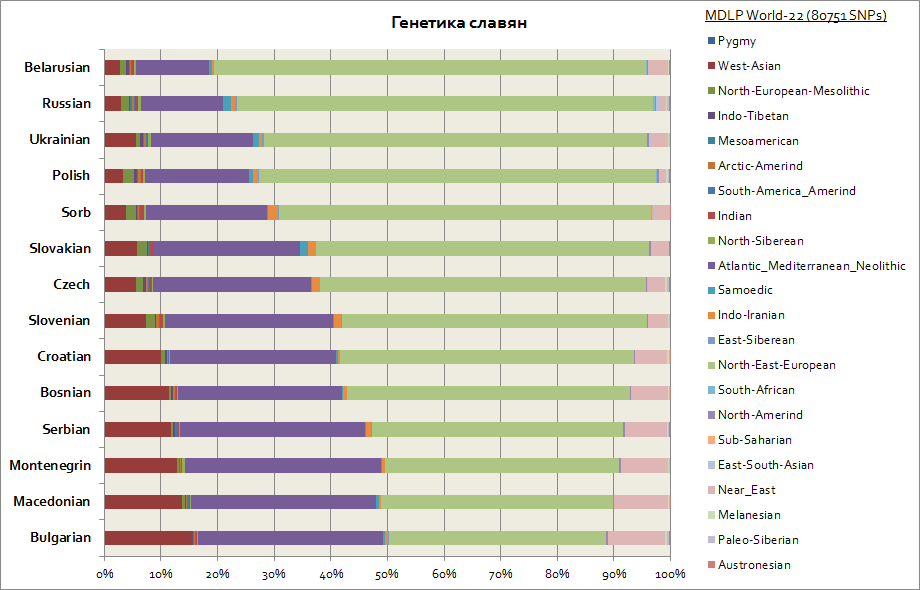
แผนภูมินี้แสดงภาพชาวสลาฟทั้งหมด รวมถึงชาวตะวันตก - โปแลนด์และเช็ก เช่นเดียวกับทางใต้ - เซอร์เบีย บัลแกเรีย มาซิโดเนีย เป็นต้น
ชาวสลาฟทั้งหมดมี 2 องค์ประกอบหลัก ได้แก่ ยุโรปตะวันออกและแอตแลนโต-เมดิเตอร์เรเนียน อย่างแรกคือสูงสุดสำหรับชาวเบลารุสและอย่างที่สองสำหรับชาวสลาฟทางตอนใต้ทั้งหมด - ชาวเซิร์บ, มาซิโดเนีย, บัลแกเรีย ส่วนประกอบของยุโรปตะวันออกเป็นแหล่งกำเนิดหลักในหมู่ชาวสลาฟ และส่วนประกอบของแอตแลนโต-เมดิเตอร์เรเนียนนั้นได้มามากขึ้นเมื่อชาวสลาฟอพยพไปยังคาบสมุทรบอลข่าน ชาวยูเครนตะวันตกและชาวสโลวาเกียมีส่วนประกอบของซามอยดิคเพิ่มขึ้นเล็กน้อยเมื่อเทียบกับเพื่อนบ้าน ชาวสลาฟ- เบลารุส, เช็ก, โปแลนด์; นี่อาจเป็นร่องรอยทางพันธุกรรมของการอพยพในยุคกลางของ Huns และ Ugrians ไปยังยุโรปกลาง
พันธุศาสตร์ของชาวสลาฟ รัสเซียและตาตาร์ เยอรมัน คอเคเชียน ยิว ฯลฯ
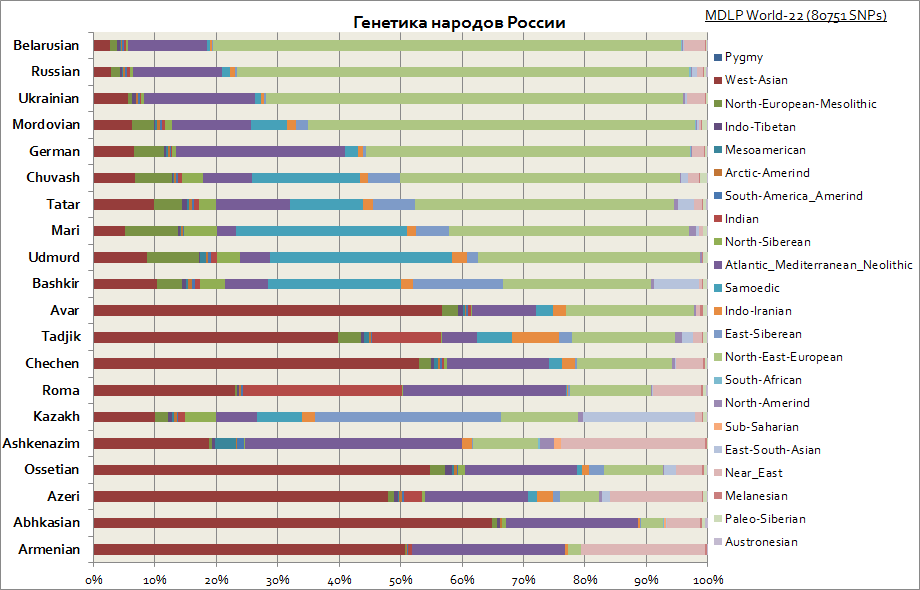
กราฟนี้แสดงต้นกำเนิดที่แตกต่างกันในหมู่ชนชาติรัสเซีย ดังที่เห็นได้ในหมู่ชาวสลาฟองค์ประกอบหลักคือยุโรปตะวันออกในขณะที่ในหมู่ประชาชนในภูมิภาคโวลก้าส่วนแบ่งของส่วนประกอบของไซบีเรียเพิ่มขึ้น ในขณะที่ชาวคอเคเชียน ส่วนประกอบของเอเชียตะวันตก ทะเลเมดิเตอร์เรเนียนและตะวันออกกลางมีลักษณะเฉพาะมากที่สุด
พันธุศาสตร์ของ Finns, Ugrians, Udmurts, Hungarians, Saami ฯลฯ
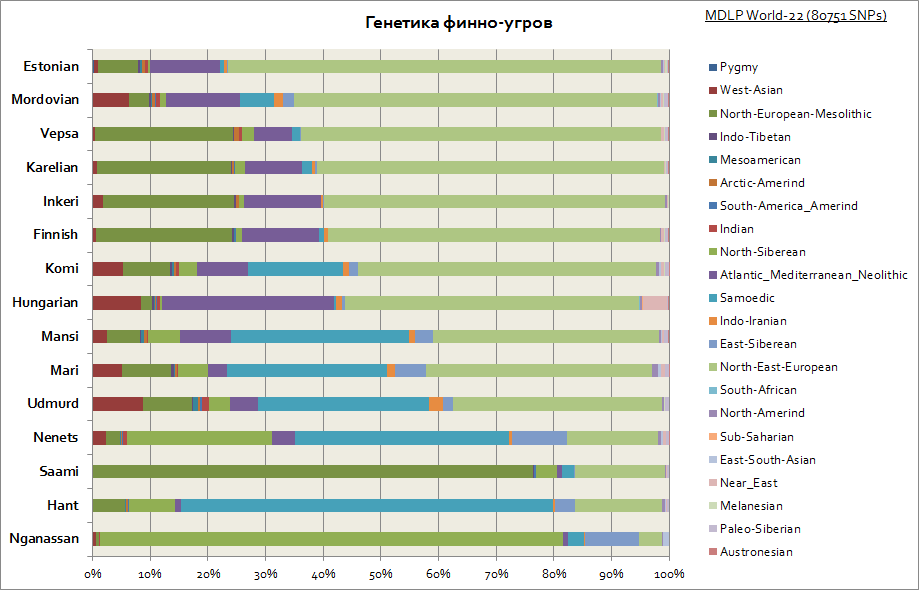
อย่างที่เห็น Finns, Vepsians และ Karelian มีลักษณะคล้ายคลึงกัน ต้นกำเนิดทางพันธุกรรมกับพวกสลาฟ พวกเขายังมีส่วนประกอบของยุโรปตะวันออกที่ใหญ่ที่สุด โดยลดลงใกล้กับเทือกเขาอูราลและภูมิภาคโวลก้า โดยมีส่วนประกอบของไซบีเรียเพิ่มขึ้นในภูมิภาคนี้ นอกจากนี้ ชาว Finno-Ugric ทั้งหมดยังมีองค์ประกอบสำคัญของหินหินของยุโรป ซึ่งถึงเกือบ 80% ในหมู่ชาวซามิ และมีความเกี่ยวข้องกับประชากรก่อนยุคอินโด-ยูโรเปียนและยุคก่อนยุคหินใหม่ในยุโรป สำหรับชาวฮังกาเรียนโดยรวม ชุดขององค์ประกอบทางพันธุกรรมเหล่านี้เป็นลักษณะเฉพาะ เช่นเดียวกับประชากรอื่น ๆ ในภูมิภาคคาร์เพเทียนและยุโรปกลาง
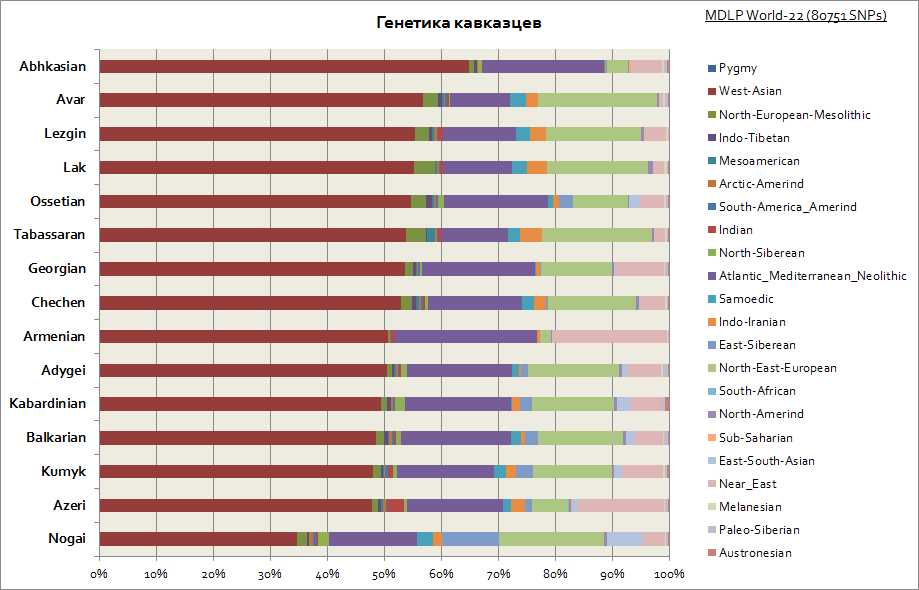
อย่างที่เห็น คอเคซัสทั้งหมดมีลักษณะต้นกำเนิดทางพันธุกรรมที่ค่อนข้างคล้ายคลึงกัน ซึ่งเป็นสัดส่วนที่มากของส่วนประกอบของเอเชียตะวันตกและทะเลเมดิเตอร์เรเนียน มีเพียง Nogais เท่านั้นที่โดดเด่นเล็กน้อย - พวกเขามีส่วนประกอบของไซบีเรียนเพิ่มขึ้น

ดังที่เห็นได้ใน Ashkenazim และ Sephardim มีความถี่สูงในส่วนประกอบของเอเชียตะวันตก แอตแลนโต-เมดิเตอร์เรเนียน และตะวันออกกลาง ในเวลาเดียวกัน Ashkenazim มีส่วนประกอบของไซบีเรียเพิ่มขึ้นเล็กน้อย ซึ่งอาจเกิดจากมรดกของ Khazar และเพิ่มขึ้นมากถึง 30% ของส่วนประกอบของยุโรปตะวันออก ซึ่งตามตัวบ่งชี้นี้ ทำให้พวกเขาเข้าใกล้ ประเทศทางตอนใต้ของยุโรป
เฉพาะชาวยิวเอธิโอเปียและชาวยิวอินเดียเท่านั้นที่ถูก "บริษัท" ของพวกเขาขับไล่ กลุ่มแรกมีสัดส่วนสูงในแอฟริกาตอนใต้ของทะเลทรายซาฮารา (มากถึง 40%) ในขณะที่กลุ่มหลังมีส่วนแบ่งขององค์ประกอบทางพันธุกรรมของเอเชียใต้ ซึ่งเรียกว่าอินเดียแบบมีเงื่อนไข (มากถึง 50%)
พันธุศาสตร์ของ Tatars, Bashkirs, Azerbaijanis, Chuvashs เป็นต้น
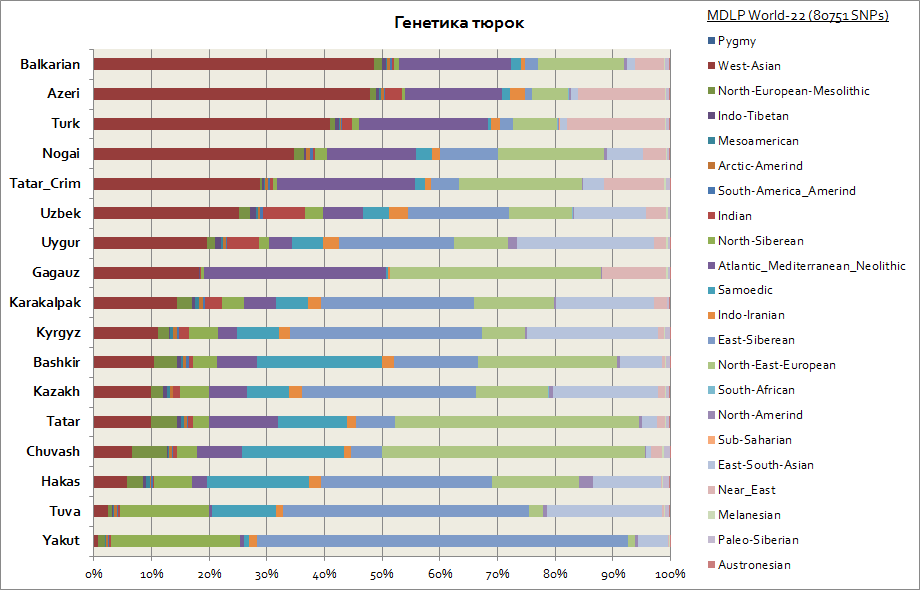
ชาวเติร์กในแง่พันธุกรรมกลายเป็นกลุ่มชาติพันธุ์ที่ต่างกันมากที่สุดกลุ่มหนึ่ง เนื่องจากองค์ประกอบทางพันธุกรรมของพวกเขาแตกต่างกันอย่างมาก ดังนั้นเนื่องจากบ้านเกิดหลักของพวกเติร์กคือไซบีเรียผู้คนเช่น Yakuts, Tuvans, Khakasses จึงยังคงรักษาส่วนประกอบ autosomal ของไซบีเรียตะวันออกไว้ในเปอร์เซ็นต์ที่ใหญ่ที่สุดซึ่งสูงถึง 30 ถึง 65% ในพวกเขา องค์ประกอบทางพันธุกรรมนี้เป็นองค์ประกอบหลักในหมู่คีร์กีซและคาซัค ส่วนประกอบที่เหลือทำให้พวกเติร์กใกล้ชิดกับผู้คนจากภูมิภาคที่อยู่อาศัยมากขึ้น ดังนั้นสำหรับ Yakuts และ Tuvans นี่คือส่วนประกอบของไซบีเรียเหนือและ Samoyed โดยรวมแล้วไซบีเรียนทั้ง 3 ตัวนี้ในบรรดายาคุตมีมากถึง 90% ในบรรดาทูวานมากถึง 70% โดยเพิ่มขึ้นเป็น 20% ขององค์ประกอบเอเชียตะวันออก - ใต้ซึ่งเกี่ยวข้องกับกระแสการย้ายถิ่นฐานของประชากรมากขึ้น เอเชียตะวันออก. สำหรับ Bashkirs ส่วนแบ่งของส่วนประกอบ 3 ของไซบีเรียสูงถึง 45% และส่วนประกอบของเอเชียตะวันออกเฉียงใต้สูงถึง 10% ตาตาร์มีข้อมูลขององค์ประกอบทางพันธุกรรมของไซบีเรีย 3 รายการโดยเฉลี่ยตั้งแต่ 25 ถึง 50% ในเวลาเดียวกันสัดส่วนของส่วนประกอบของประชากรคอเคซอยด์ในหมู่ Bashkirs นั้นสูงถึง 45% และในหมู่พวกตาตาร์โดยเฉลี่ยจาก 50 ถึง 70% พันธุศาสตร์ของอาเซอร์ไบจานและเติร์กในทางปฏิบัติไม่แตกต่างกันในขอบเขตของข้อผิดพลาด พวกเขาเช่นเดียวกับคนอื่น ๆ ในคอเคซัสและทรานคอเคเซียมีส่วนประกอบของเอเชียตะวันตก (ถึง 50%) และส่วนประกอบของแอตแลนโต - เมดิเตอร์เรเนียน (มากถึง เฉลี่ย 20%) ส่วนแบ่งของ 3 ส่วนประกอบของไซบีเรียแสดงโดยอาเซอร์ไบจาน, เติกส์และบัลการ์ - ที่ระดับ 3-7%
บทสรุป
พันธุกรรมของผู้คนไม่มีความสัมพันธ์โดยตรงและมีนัยสำคัญกับการกระจายของตระกูลภาษาหรือกับเปอร์เซ็นต์ของเครื่องหมายฝ่ายเดียว - กลุ่มแฮ็ปโล Y-DNA และ mt-DNA ซึ่งแสดงอยู่ในประชากรเฉพาะ ความสัมพันธ์ที่ยิ่งใหญ่ที่สุดสามารถตรวจสอบได้ตามหลักดินแดนและภูมิศาสตร์ ดังนั้นส่วนแบ่งของส่วนประกอบของไซบีเรียซึ่งเป็นลักษณะทั่วไปของ เผ่าพันธุ์มองโกลอยด์ค่อยๆ ลดลงจากตะวันออกไปตะวันตก และสัดส่วนของลักษณะส่วนประกอบของเผ่าพันธุ์คอเคซอยด์ก็เพิ่มขึ้นตามไปด้วย ในดินแดนชายแดนตามแนวจากทางเหนือของเทือกเขาอูราลถึง เอเชียกลางอัตราส่วนโดยประมาณเท่ากัน ในภูมิภาคทางตะวันออกของทะเลสาบไบคาลลักษณะองค์ประกอบทางพันธุกรรมของเผ่าพันธุ์คอเคเชียนขนาดใหญ่นั้นไม่ได้แสดงอยู่จริงในขณะเดียวกันในภูมิภาคทางตะวันตกของแนว Pechora-Volga ส่วนประกอบของไซบีเรียของเผ่าพันธุ์มองโกลอยด์ขนาดใหญ่กำลังหายไป .การแพร่กระจายขององค์ประกอบทางพันธุกรรมของยุโรปตะวันออกไปยังไซบีเรียเกิดขึ้นในระดับมากแล้วในยุคสำริด (วัฒนธรรมของ Andronov Circle) แม้ว่ายอดเขาแต่ละแห่งทางตะวันออกสุดของไซบีเรียท่ามกลาง Chukchi อาจเกี่ยวข้องกับการอพยพของรัสเซียใน ศตวรรษที่ 17.
ส่วนแบ่งขององค์ประกอบ Sub-Saharan ซึ่งเป็นลักษณะของการแข่งขัน Negroid นั้นกระจายไปทั่วแอฟริกา - จนถึงทะเลเมดิเตอร์เรเนียนทางตอนใต้และชายแดนทางตอนเหนือของทวีปแอฟริกาถึงจุดสูงสุดในบริเวณเส้นศูนย์สูตรและไม่เคยพบนอกเลย พื้นหลังสีอ่อนกระจายอยู่บนคาบสมุทรอาหรับและทางตอนใต้ของที่ราบสูงอิหร่าน
ภูมิศาสตร์ขององค์ประกอบทางพันธุกรรม
อเล็กเซย์ ซอร์ริน
โครงการ



