Hvilke europeiske folk er russere nærmere genetisk. Genetikk til russiske ukrainere, hviterussere og tatarer, slaver og kaukasiere, jøder, finner og andre befolkninger
Pentagon anerkjente eksperimenter med biomaterialer fra russiske borgere
Den apokalyptiske antagelsen om amerikanernes mulige utvikling av biologiske våpen har fått sterk bekreftelse. Pentagon erkjente det faktum å samle inn biomaterialer fra russiske borgere.
Ifølge Pentagon-talsmann Bo Downey, gjennomfører US Air Force 59th Air Medical Groups senter for molekylær forskning muskel- og skjelettstudier for å identifisere ulike biomarkører assosiert med traumer. Prøver av russisk opprinnelse kreves bare fordi den første batchen var fra Russland, og nå trengs en identisk for kontroll.
Husk at det amerikanske flyvåpenet har til hensikt å kjøpe 12 prøver av RNA-molekylet og 27 prøver av leddvæsken til russerne. Kunngjøringen ble lagt ut på US Government Procurement Portal. Samtidig understreker kontrakten at blant alle innbyggere i Russland er kunden kun interessert i europeere, og innvandrere, for eksempel fra Ukraina, vil ikke bli vurdert.
Spørsmålet om å samle biomaterialer fra russere begynte å bli diskutert i samfunnet etter en offentlig uttalelse om dette fra Vladimir Putin. Ifølge ham samles biomaterialer inn "for forskjellige etniske grupper og mennesker som lever i forskjellige geografiske punkter Den russiske føderasjonen". "Spørsmålet er hvorfor gjør de dette?" spurte presidenten retorisk på et møte med menneskerettighetsaktivister.
Senere forklarte Dmitry Peskov at det dreide seg om informasjon mottatt gjennom spesialtjenestene.
Putins ord provoserte frem kaustisk latterliggjøring fra deler av det russiske samfunnet. "De gamle inkaene var også redde for slike skitne triks, så den øverste herskeren hadde til og med spesielle tjenestejenter, som hadde til oppgave å spise hår, spytt, negleklipp og annet biomateriale som var til overs fra den store, for å forhindre at søppel havnet i uvennlighet. hender», er lærer Andrey sitert av media Nikulin.
Likevel forbereder landet seg på å motvirke den nye trusselen på lovgivende nivå. Det forventes at loven om beskyttelse av biomaterialer i desember vil bli sendt til statsdumaen. "I dag har vi mange laboratorier som utfører klinisk forskning, dette er utenlandske laboratorier, for eksempel Invitro. Vi slipper dem inn i de mest intime," forklarte Gennady Onishchenko, første nestleder i statsdumaens komité for utdanning og vitenskap, problemet.
I «Invitro» benektet de sitt engasjement i eksport av biomaterialer. På sin side rapporterte Institute of General Genetics ved det russiske vitenskapsakademiet "SP" at denne institusjonen var involvert i innsamling av biomaterialer. "Den vitenskapelige direktøren for instituttet vårt, akademiker Nikolai Yankovsky, har nettopp organisert og ledet programmet for innsamling av biomaterialer," sa Ilya Zakharov-Gezekhus, en ansatt ved instituttet. Det var ikke mulig å umiddelbart kontakte Yankovsky "SP" selv.
Senior Forsker laboratorium for genetikk ved Institute of Developmental Biology ved det russiske vitenskapsakademiet Alexey Kulikov er tilbøyelig til å rettferdiggjøre sine amerikanske kolleger.
Du må forstå hva amerikanerne gjorde. De så på hvordan gener fungerer hos pasienter med påvirket muskel- og skjelettsystem og så på sammensetningen av leddvæsken. De kjøpte opprinnelig biomaterialer et sted i Øst-Europa fra syke representanter for den slaviske nasjonen, og derfor trenger de også kontrollprøver av mennesker som har det bra med muskel- og skjelettsystemet fra representanter for den slaviske nasjonen: russere, ukrainere, hviterussere, etc.
"SP": - Vel, hvordan? I anbudet står det eksplisitt at biomaterialer fra Ukraina ikke vil bli vurdert. De trenger russerne...
Det har fortsatt ingenting med intensjoner å gjøre. Genetiske våpen er alt tull. Dette er urealistisk, fordi mennesker i verden er for polymorfe - veldig forskjellige. Det er vanskelig å finne på noe som fungerer for noen og ikke fungerer for andre. Jeg tenker her vi snakker bare om betingelsene for eksperimentet. Spise vitenskapelig erfaring og det er kontroll. Kontrollmaterialer må være fra samme region.
"SP": - Alt er slik, hvis disse "vitenskapelige studiene" ikke ble utført av militæret, Pentagon ...
Vitenskapelig forskning kan også utføres av militæret. Vi har også spesifikke medisinske oppgaver som løses av representanter for rettshåndhevende instanser. Kanskje vi snakker om den lave mobiliteten til pilotene. De må i lang tid sitte. Derfor er det grunnleggende viktig at det ikke er problemer med muskel- og skjelettsystemet. Derfor er amerikanere interessert i disse sykdommene og hvilke spesifikke gener som er ansvarlige for dette.
På sin side mener sjefen for laboratoriet ved Institutt for generell genetikk ved det russiske vitenskapsakademiet, Sergey Kiselev, at det er for risikabelt å lage et genetisk våpen og det er lettere å drepe mennesker. tradisjonelle metoder.
Biomaterialer fra Russland er overført og blir overført. I hvert fall, som Onishchenko sa, innenfor rammen av kliniske studier. For i løpet av de siste 25 årene har dusinvis av utenlandske selskaper gjennomført kliniske legemiddelforsøk i Russland. Biomaterialer er nødvendig for å forstå hvordan medisiner virker.
Selvfølgelig kan enhver slik prøve brukes både til de ovennevnte formålene og for noen andre. Fordi det fortsatt er en nasjonal biologisk prøve. Det viktigste er hvordan informasjonen som er innhentet ved hjelp av prøven senere vil bli brukt. Genetisk informasjon kan brukes til en rekke formål, inkludert for personlig vinning.
"SP": - Hva?
Dagens teknologier lar oss bestemme genomet til hver person i detalj. Det vil si at ved å ta en DNA-prøve fra en skje i spisesalen til CIA-bygningen, kan du finne ut om etterretningsoffiseren virkelig kommer fra delstaten Minnesota, som han skrev da han søkte jobb, eller om han er fra Øst-Sibir . Det vil si at du veldig nøyaktig kan knytte en person til et sted.
Men for dette må du først lage genetisk kart territorier. Jeg tror at de interesserte etterretningstjenestene i mange land i verden har laget slike kart over territorier i lang tid. For å kunne identifisere personer i ulike omstendigheter.
For eksempel folk med farlige yrker. Hvis en av dem er revet i stykker, så er det mulig, ved hjelp av et genetisk kart, å fastslå hvem han er, hvor han kom fra og hylle ham. Altså innenfor landet. Men hvis denne informasjonen kommer inn i tredjeparter, kan den brukes i deres interesser.
"SP": - V denne saken vi er interessert i sannsynligheten for å lage et genetisk, etnisk våpen ...
Etter min mening er det meningsløst å lage et slikt våpen. For det første, for å drepe en person, er det nødvendig å påvirke vitale funksjoner som respirasjon, blodsirkulasjon osv. Det er få av dem, og de er like for alle mennesker. Å finne en etnisk viktig vital funksjon er neppe mulig.
Og for det andre, hvem er amerikanerne? De har ikke en titulær nasjon. Alle blandet seg der, inkludert russere, kinesere, angelsaksere. Derfor, hvis det opprettes et våpen mot russerne, vil noen av amerikanerne falle inn under dens handling, siden de enten er russere eller deres etterkommere.
Derfor, jo større spredning av mennesker rundt planeten, blandingen av genomer, jo mindre sannsynlig vil det være å lage et genetisk våpen. Det er ulønnsomt. Det er enklere, billigere og mer effektivt å "bløte" på en annen måte.
"SP": - Likevel utfører Pentagon slike studier, og i dag innrømmet representanten dette direkte ...
Jeg så på vilkårene for anbudet. Denne flyvåpenbasen ba om RNA-prøver. Men RNA er subgenetisk materiale. Det er et mellomledd mellom det menneskelige genomet og den funksjonelle tilstanden til cellen. RNA gjenspeiler spesialiseringen til hver enkelt celle på et eller annet tidspunkt. Det vil si genomet - DNA i hver celle i kroppen er alltid det samme. Og RNA i hver celle er forskjellig, fordi det er et portrett av hvordan genomet fungerer hvert minutt. Derfor vil en RNA-prøve tatt fra stortåen være forskjellig fra RNA fra et hvilket som helst annet organ. Amerikanerne har ikke angitt hvor nøyaktig de trenger RNA.
Det andre de ba om var leddvæske fra leddkapselen. Dessuten ble det i tilbudsvilkårene fastsatt at væsken kunne tas fra en annen giver, ikke fra den som RNA ble tatt fra. Det vil si at amerikanerne ikke trenger noen forbindelse mellom RNA og leddvæske. Det ser ut som de tok prøver for forskjellige studier, ikke relatert til hverandre.
"SP": - Dette økte imidlertid ikke klarheten ...
Kanskje de hvitvasket penger der. Eller elevene lærte å skrive anbud.
Men det tidligere medlemmet av FNs kommisjon for biologiske våpen, Igor Nikulin, er ikke i tvil om de aggressive planene til Pentagon.
Selvfølgelig er dette et forsøk fra amerikanerne på å drive forskning for militære formål. Pentagon er ikke en veldedig eller humanitær organisasjon designet for å gagne menneskeheten. Snarere tvert imot. Her er det mulig å bruke virus som vil virke selektivt.
Slike virus eksisterer fortsatt i dag. For eksempel Ebola, Lassa, Marburg. De hemorragiske feberne som de forårsaker virker hovedsakelig på negroide rase, fugleinfluensa - på mongoloid, SARS på indoeuropeere.
"SP": - I konkurransebetingelsene, av en eller annen grunn, er det gjort unntak for Ukraina ...
I Ukraina ble det amerikanske programmet gjennomført for 5-7 år siden. Og nå har de bare epidemier. Enten meslinger, eller røde hunder, eller tuberkulose, eller stivkrampe, eller kolera osv. Og så tilbyr amerikanerne dem vaksiner mot dette. Veldig komfortabelt.
I løpet av de siste ti årene har USA brukt titalls milliarder dollar, satt opp mer enn fire hundre laboratorier rundt om i verden, der nye typer biologiske våpen og vaksiner utvikles. Rundt førti laboratorier er lokalisert i land tidligere USSR. Disse er Ukraina, Moldova, Georgia, Armenia, Aserbajdsjan, Usbekistan, Kirgisistan og Kasakhstan. Slik er det kontrollerte kaoset.
"SP": - Men våre russiske genetikere er skeptiske til den sannsynlige utviklingen av våpen rettet mot en eller annen etnisk gruppe ...
Hva annet har de igjen å gjøre? Mange av dem er på tilskudd. Hvis jeg var på stipend, ville jeg også vært stille. Men siden amerikanerne ikke vil tilby meg noen tilskudd, kan jeg snakke fritt om disse temaene.
Vurder nyhetene
Partnernyheter: Av natur er den genetiske koden til alle mennesker ordnet på en slik måte at hver har 23 par kromosomer, som lagrer all den arvelige informasjonen som er arvet fra begge foreldrene.
Dannelsen av kromosomer skjer på tidspunktet for meiose, når hver tilfeldig tar omtrent halvparten fra mors kromosom og halvparten fra morskromosomet, hvilke spesifikke gener som vil arves fra mor og hvilke fra far er. ikke kjent, alt avgjøres ved en tilfeldighet.
Bare ett mannlig kromosom, Y, deltar ikke i dette lotteriet; det overføres fullstendig fra far til sønn som en stafettstokk. Jeg vil presisere at kvinner ikke har dette Y-kromosomet i det hele tatt.
I hver påfølgende generasjon forekommer mutasjoner i visse regioner av Y-kromosomet, kalt loci, som vil bli overført til alle påfølgende generasjoner. maskulin.
Det var takket være disse mutasjonene at det ble mulig å rekonstruere slekten. Det er bare rundt 400 loci på Y-kromosomet, men bare rundt hundre brukes til sammenlignende haplotypeanalyse og slektsrekonstruksjon.
I de såkalte lociene, eller de kalles også STR-markører, er det fra 7 til 42 tandem-repetisjoner, helhetsbilde som er unikt for hver enkelt. Etter et visst antall generasjoner oppstår mutasjoner og antall tandem-repetisjoner endres opp eller ned, og dermed vil man se på fellestreet at jo flere mutasjoner, jo eldre er felles stamfar for haplotypegruppen.
Haplogruppene selv bærer ikke genetisk informasjon, pga genetisk informasjon er lokalisert i autosomer - de første 22 parene av kromosomer. Du kan se fordelingen av genetiske komponenter i Europa. Haplogrupper er bare markører for svunne dager, ved begynnelsen av dannelsen av moderne folk.
Hvilke haplogrupper er mest vanlige blant russere?
folkeslag |
Menneskelig |
||||||||
|---|---|---|---|---|---|---|---|---|---|
østlige, vestlige og sørlige slaver. |
|||||||||
| russere(Nord) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | |
| russere(senter) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | |
| russere(sør) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 3 | |
| russere (Alle Flotte russere) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| hviterussere | 574 | 52 | 10 | 3 | 16 | 10 | 3 | Russere, slaver, indoeuropeere og haplogruppene R1a, R1b, N1c, I1 og I2I antikken, for rundt 8-9 tusen år siden, var det en språklig gruppe som la grunnlaget for den indoeuropeiske språkfamilien (i det første stadiet mest sannsynlig er dette haplogruppene R1a og R1b). Den indoeuropeiske familien inkluderer slike språklige grupper som indo-iranere (Sør-Asia), slaver og baltere ( Øst-Europa), keltere (Vest-Europa), tyskere (sentral, Nord-Europa).Kanskje de også hadde felles genetiske forfedre, som for rundt 7 tusen år siden, som et resultat av migrasjoner, havnet i forskjellige deler av Eurasia, noen dro sørover og østover (R1a-Z93), og la grunnlaget for de indo-iranske folkene og språk (som i stor grad deltok i etnogenesen til de turkiske folkene), og en del forble på Europas territorium og la grunnlaget for dannelsen av mange europeiske folk (R1b-L51), inkludert slaverne og russere spesielt (Rla-Z283, Rlb-L51). På forskjellige stadier av dannelsen, allerede i antikken, var det kryss av migrasjonsstrømmer, noe som forårsaket tilstedeværelsen av et stort antall haplogrupper i alle europeiske etniske grupper. De slaviske språkene dukket opp fra den en gang forente gruppen av balto-slaviske språk (antagelig den arkeologiske kulturen til den sene Corded Ware). Ifølge beregningene til språkforskeren Starostin skjedde dette for rundt 3,3 årtusener siden. Periode fra det 5. århundre f.Kr til det 4.-5. århundre e.Kr kan betraktes som betinget protoslavisk, tk. Balterne og slaverne hadde allerede delt seg, men det var ingen slaver selv ennå, de vil dukke opp litt senere, på 4-600-tallet e.Kr. I det innledende stadiet av dannelsen av slaverne var sannsynligvis omtrent 80% haplogruppene R1a-Z280 og I2a-M423. I det innledende stadiet av dannelsen av balterne var sannsynligvis omtrent 80 % haplogrupper N1c-L1025 og R1a-Z92. Innflytelsen og skjæringspunktet mellom migrasjonene til balterne og slaverne var helt fra begynnelsen, fordi denne inndelingen i mange henseender er vilkårlig, og gjenspeiler generelt bare hovedtrenden, uten detaljer. De iranske språkene er indoeuropeiske, og dateringen deres er som følger - de eldste, fra det andre årtusen f.Kr. til det 4. århundre f.Kr., det midterste - fra det 4. århundre f.Kr. til 900-tallet e.Kr., og en ny - fra 900-tallet e.Kr. Inntil nå. Det vil si at de eldste iranske språkene dukker opp etter avgangen til en del av stammene som snakket indoeuropeiske språk fra Sentral-Asia til India og Iran. Deres viktigste haplogrupper var sannsynligvis R1a-Z93, J2a, G2a3. Den vestiranske språkgruppen dukket opp senere, rundt det 5. århundre f.Kr. Dermed ble indo-arierne, kelterne, tyskerne og slaverne i akademisk vitenskap indoeuropeere, dette begrepet er det mest passende for et så stort og mangfoldig gruppe. Dette er helt riktig. I det genetiske aspektet er heterogeniteten til indoeuropeerne slående både i Y-haplogrupper og i autosomer. Indo-iranerne preges i større grad av den vestasiatiske genetiske påvirkningen av BMAC. I følge de indiske vedaene var det indo-arierne som kom til India (Sør-Asia) fra nord (fra Sentral-Asia), og det var deres salmer og legender som dannet grunnlaget for de indiske vedaene. Og la oss fortsette videre, la oss berøre lingvistikk, fordi dette russiske språket (og dets beslektede baltiske språk, for eksempel litauisk som en del av det en gang eksisterende balto-slaviske språksamfunnet) er relativt nær sanskrit sammen med keltiske, germanske og andre språk av den store indoeuropeiske familien. Men i den genetiske planen var indo-arierne allerede i større grad vestasiater, da de nærmet seg India, ble også vedoid-innflytelsen intensivert. Så det ble klart det haplogruppe R1a i DNA-slektsforskning er dette en vanlig haplogruppe for en del av slaverne, en del av tyrkerne og en del av indo-arierne (fordi det naturlig nok var representanter for andre haplogrupper i deres miljø), en del haplogruppe R1a1 under migrasjoner langs den russiske sletten ble de en del av de finsk-ugriske folkene, for eksempel mordovierne (Erzya og Moksha). En del av stammene (for haplogruppe R1a1 dette er en undergruppe av Z93) under migrasjoner brakte de dette indoeuropeiske språket til India og Iran for rundt 3500 år siden, det vil si i midten av det 2. årtusen f.Kr. I India, ved arbeidet til den store Panini, ble det omgjort til sanskrit i midten av det 1. årtusen f.Kr., og i Persia-Iran ble de ariske språkene grunnlaget for en gruppe iranske språk, hvorav de eldste dateres tilbake til det 2. årtusen f.Kr. Disse dataene er bekreftet: DNA slektsforskning og lingvistikk er korrelert her. Stor del haplogrupper R1a1-Z93 selv i antikken sluttet de seg til de tyrkiske etniske gruppene og markerer i dag tyrkernes migrasjon på mange måter, noe som ikke er overraskende med tanke på antikken haplogruppe R1a1 mens representantene haplogrupper R1a1-Z280 var en del av de finsk-ugriske stammene, men under bosettingen av de slaviske kolonistene ble mange av dem assimilert av slaverne, men selv nå blant mange folk, for eksempel, er Erzya fortsatt den dominerende haplogruppen R1a1-Z280. Alle disse nye dataene var i stand til å gi oss DNA slektsforskning, spesielt de omtrentlige datoene for migrasjonene til haplogruppebærere på territoriet til den moderne russiske sletten og Sentral-Asia i forhistorisk tid. Så forskere til alle slaver, keltere, tyskere, etc. ga navnet på indoeuropeerne, noe som er sant fra et språkvitenskapelig synspunkt. Hvor kom disse indoeuropeerne fra? Faktisk var det indoeuropeiske språk lenge før migrasjonene til India og Iran, over hele den russiske sletten og så langt som til Balkan i sør, og så langt som til Pyreneene i vest. Senere ble språket spredt til Sør-Asia – både til Iran og India. Men i genetiske termer er korrelasjonene mye mindre. "Det eneste berettigede og aksepterte på det nåværende tidspunkt i vitenskapen er bruken av begrepet "ariere" bare i forhold til stammene og folkene som snakket de indo-iranske språkene." Så i hvilken retning gikk den indoeuropeiske strømmen - mot vest, til Europa, eller omvendt, mot øst? I følge noen estimater er den indoeuropeiske språkfamilien rundt 8500 år gammel. Indoeuropeernes forfedres hjem er ennå ikke bestemt, men ifølge en av versjonene kan det være Svartehavsregionen - sørlig eller nordlig. I India, som vi allerede vet, ble det indo-ariske språket introdusert for rundt 3500 år siden, antagelig fra territoriet til Sentral-Asia, og arierne selv var en gruppe med forskjellige genetiske Y-linjer, som R1a1-L657, G2a, J2a, J2b, H, osv. Haplogruppe R1a1 i Vest- og Sør-EuropaAnalyse av 67 markørhaplotyper haplogruppe R1a1 fra alle europeiske land gjorde det mulig å bestemme den omtrentlige migrasjonsveien til R1a1-forfedre i retning Vest-Europa. Og beregninger viste at nesten i hele Europa, fra Island i nord til Hellas i sør, var den felles stamfaren til haplogruppen R1a1 en for omtrent 7000 år siden!Med andre ord, etterkommerne, som et stafettløp, videreførte sine haplotyper til sine egne etterkommere fra generasjon til generasjon, og spredte seg i prosessen med migrasjon fra det samme historiske stedet - som viste seg å være antagelig Ural- eller Svartehavslavlandet . På et moderne kart er dette land hovedsakelig i Øst- og Sentral-Europa - Polen, Hviterussland, Ukraina, Russland. Men rekkevidden av eldre haplotyper av haplogruppen R1a1 fører mot øst - til Sibir. Og levetiden til den første stamfaren, som er indikert av de eldste, mest muterte haplotypene, er 7,5 tusen år siden. På den tiden var det ingen slaver, ingen tyskere, ingen keltere. Sentral- og Øst-EuropaPolen, den felles stamfaren til R1a1 levde for rundt 5000 år siden (hovedsakelig en undergruppe av R1a1-M458 og Z280). For russisk-ukrainsk - 4500 år siden, som praktisk talt sammenfaller innenfor nøyaktigheten av beregninger.Og selv om fire generasjoner ikke er en forskjell for slike termer. I det moderne Polen haplogruppe R1a1 et gjennomsnitt på 56 %, og i enkelte områder opptil 62 %. Resten er stort sett vesteuropeiske haplogruppe R1b(12%), skandinavisk haplogruppe I1(17 %) og Baltikum haplogruppe N1c1 (8%). I Tsjekkia og Slovakia levde en felles proto-slavisk stamfar for 4200 år siden. Bare litt mindre enn russerne og ukrainerne. Det vil si at vi snakker om å bosette oss i territoriene til det moderne Polen, Tsjekkia, Slovakia, Ukraina, Hviterussland, Russland - alt innen bare noen få generasjoner, men for mer enn fire tusen år siden. I arkeologi er en slik nøyaktighet av datering helt utenkelig. I Tsjekkia og Slovakia etterkommere haplogruppe R1a1 ca. 40 %. Resten har stort sett vesteuropeisk R1b(22-28%) skandinavisk I1 og Balkan haplogruppe I2a(kumulativt 18 %) På territoriet til det moderne Ungarn bodde den felles stamfaren R1a1 for 5000 år siden. Det er nå opptil en fjerdedel av etterkommerne av haplogruppen R1a1. Resten har hovedsakelig den vesteuropeiske haplogruppen R1b (20%) og de kombinerte skandinaviske I1 og Balkan I2 (26% totalt) haplogruppene. Gitt at ungarerne snakker språket til den finsk-ugriske språkgruppen, er den vanligste haplogruppen der N1c1 i de gamle ungarske rike gravene til magyarene er restene av menn med haplogruppen hovedsakelig funnet N1c1, som var de første lederne av stammene som deltok i dannelsen av imperiet. I Litauen og Latvia er den felles stamfaren rekonstruert til en dybde på 4800 år. I utgangspunktet er det i dag subclade Z92, Z280 og M458. Den vanligste blant litauere er den baltiske haplogruppen N1c1, som når 47%. Generelt er Litauen og Latvia preget av den sør-baltiske underkladen L1025 av haplogruppen N1c1. Generelt er situasjonen klar. Jeg vil bare legge til at i europeiske land - Island, Nederland, Danmark, Sveits, Belgia, Litauen, Frankrike, Italia, Romania, Albania, Montenegro, Slovenia, Kroatia, Spania, Hellas, Bulgaria, Moldova - levde den felles stamfaren 5000- For 5500 år siden er det umulig å være mer presis. Dette er en felles stamfar haplogruppe R1a for alle landene som er oppført. Den felles europeiske stamfaren, så å si, uten å regne Balkan-regionen vist ovenfor, det mulige forfedrehjemmet til indoeuropeerne for rundt 7500 år siden. Andel av transportører haplogruppe R1a1 i følgende land varierer, fra 4 % i Holland og Italia, 9 % i Albania, 8–11 % i Hellas (opptil 14 % i Thessaloniki), 12–15 % i Bulgaria og Hercegovina, 14–17 % i Danmark og Serbia, 15-25 % i Bosnia og Makedonia, 3 % i Sveits, 20 % i Romania og Ungarn, 23 % på Island, 22-39 % i Moldova, 29-34 % i Kroatia, 30-37 % i Slovenia (16 % på Balkan som helhet), og på samme tid - 32-37 % i Estland, 34-38 % i Litauen, 41 % i Latvia, 40 % i Hviterussland, 45-54 % i Ukraina. I det østeuropeiske Russland haplogruppe R1a, som jeg allerede har nevnt, et gjennomsnitt på 47%, på grunn av den høye andelen av Baltikum haplogruppe N1c1 i nord og nordvest i Russland, men i sør og i sentrum av Russland, når andelen av forskjellige undergrupper av haplogruppen R1a 55%. Tyrkere og haplogruppe R1a1Haplotyper av forfedre er forskjellige overalt, forskjellige regioner er preget av sine egne underklader. Folkene i Altai og andre tyrkere har også høye prosentandeler av haplogruppen R1a1, bashkirene når 40% av subkladen Z2123. Dette er en barnelinje fra Z93 og kan kalles typisk turkisk og ikke relatert til migrasjonene til indo-iranerne.I dag et stort antall haplogruppe R1a1å være i Sayano-Altai-regionen, blant den turkiske befolkningen i Sentral-Asia. Blant kirgiserne, når 63%. Du kan ikke kalle dem russere eller iranere. Det viser seg å nevne alle haplogruppe R1a1 et enkelt navn - grov overdrivelse, i det minste, men mest - uvitenhet. Haplogrupper er ikke etniske grupper; den språklige og etniske tilhørigheten til transportøren er ikke registrert på dem. Haplogrupper har heller ingen direkte relasjon til gener. Türkene er hovedsakelig preget av ulike underklader av Z93, men i Volga-regionen er det også R1a1-Z280, muligens overført til Volga Türks fra Volga-finnene. Haplogruppe R1a1-Z93 er også typisk for arabere i moderat frekvens, for levitter - en undergruppe av Ashkenazi-jøder (undergruppen CTS6 ble bekreftet i sistnevnte). Denne linjen deltok allerede i de tidligste stadiene i etnogenesen til disse folkene. Territorium for første distribusjon haplogruppe R1a1 i Europa - dette er sannsynligvis territoriet til Øst-Europa og muligens Svartehavets lavland. Før det, sannsynligvis i Asia, muligens Sør-Asia eller Nord-Kina. Kaukasiske R1a1-haplotyperArmenia. Alder til den felles stamfaren til haplogruppen R1a1- For 6500 år siden. I utgangspunktet også en subclade av R1a1-Z93, selv om det også er R1a1-Z282.Lilleasia, Anatolske halvøy. Et historisk veiskille mellom Midtøsten, Europa og Asia. Det var den første eller andre kandidaten for det "indoeuropeiske hjemlandet". Imidlertid bodde den felles stamfaren til haplogruppen R1a1 der for rundt 6500 år siden. Det er klart at dette forfedres hjem, etter haplotypene å dømme, praktisk talt kan være i Anatolia, eller at de opprinnelige indoeuropeerne var bærere haplogruppe R1b. Men det er stor sannsynlighet for lav representasjon av individer fra Tyrkia i den generelle haplotypedatabasen. Så, både armenere og anatolere - de har alle samme stamfar, eller forfedrene er veldig nære i tid, innen flere generasjoner - dette er en undergruppe av Z93 og Z282 *. Det skal bemerkes at 4500 år før den felles stamfaren til R1a1-Z93 haplogruppen i Anatolia er i god overensstemmelse med tidspunktet for hetittenes opptreden i Lilleasia i siste kvartal av det 3. årtusen f.Kr., selv om mange R1a1-Z93 linjer kunne ha dukket opp der etter migrasjonene av de turkiske folkene til halvøya allerede i vår tid. Alexey Zorrin *** | |
Bare ett mannlig kromosom, Y, deltar ikke i dette lotteriet, det overføres fullstendig fra far til sønn som en stafettpinnen. Jeg vil presisere at kvinner ikke har dette Y-kromosomet i det hele tatt.
I hver påfølgende generasjon forekommer mutasjoner i visse regioner av Y-kromosomet, kalt loci, som vil bli overført til alle påfølgende generasjoner etter mannlig kjønn. Det var takket være disse mutasjonene at det ble mulig å rekonstruere slekten. Det er bare rundt 1000 loci på Y-kromosomet, men bare litt mer enn hundre brukes til komparativ analyse av haplotyper og rekonstruksjon av slekter.
I de såkalte lociene, eller de kalles også STR-markører, er det fra 7 til 42 tandem-repetisjoner, hvis generelle mønster er unikt for hver person. Etter et visst antall generasjoner oppstår mutasjoner og antall tandem-repetisjoner endres opp eller ned, og dermed vil man se på fellestreet at jo flere mutasjoner, jo eldre er felles stamfar for haplotypegruppen.
Haplogruppene selv bærer ikke genetisk informasjon, pga genetisk informasjon er lokalisert i autosomer - de første 22 parene av kromosomer. Du kan se fordelingen av genetiske komponenter i Europa. Haplogrupper er bare markører for svunne dager, ved begynnelsen av dannelsen av moderne folk.
Hvilke haplogrupper er mest vanlige blant russere?
| folkeslag | antall, Menneskelig | R1a1, | R1b1, | I1, | I2, | N1c1, | E1b1b1, | J2, | G2a, |
|---|---|---|---|---|---|---|---|---|---|
| østlige, vestlige og sørlige slaver. | |||||||||
| russere(Nord) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | 1 |
| russere(senter) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | 1 |
| russere(sør) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 4 | 3 |
| russere (Alle Flotte russere) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| hviterussere | 574 | 52 | 10 | 3 | 16 | 10 | 3 | 2 | 2 |
| ukrainere | 93 | 54 | 2 | 5 | 16 | 8 | 8 | 6 | 3 |
| russere(sammen med ukrainere og hviterussere) | 1874 | 48 | 7 | 4 | 13 | 16 | 4 | 3 | 3 |
| Poler | 233 | 56 | 16 | 7 | 10 | 8 | 4 | 3 | 2 |
| slovaker | 70 | 47 | 17 | 6 | 11 | 3 | 9 | 4 | 1 |
| tsjekkere | 53 | 38 | 19 | 11 | 12 | 3 | 8 | 6 | 5 |
| slovenere | 70 | 37 | 21 | 12 | 20 | 0 | 7 | 3 | 2 |
| kroatere | 108 | 24 | 10 | 6 | 39 | 1 | 10 | 6 | 2 |
| serbere | 113 | 16 | 11 | 6 | 29 | 1 | 20 | 7 | 1 |
| bulgarere | 89 | 15 | 11 | 5 | 20 | 0 | 21 | 11 | 5 |
| Baltere, finner, tyskere, grekere osv. | |||||||||
| litauere | 164 | 34 | 5 | 5 | 5 | 44 | 1 | 0 | 0 |
| latviere | 113 | 39 | 10 | 4 | 3 | 42 | 0 | 0 | 0 |
| Finner (øst) | 306 | 6 | 3 | 19 | 0 | 71 | 0 | 0 | 0 |
| Finner (vest) | 230 | 9 | 5 | 40 | 0 | 41 | 0 | 0 | 0 |
| svensker | 160 | 16 | 24 | 36 | 3 | 11 | 3 | 3 | 1 |
| tyskere | 98 | 8 | 48 | 25 | 0 | 1 | 5 | 4 | 3 |
| tyskere (bayerere) | 80 | 15 | 48 | 16 | 4 | 0 | 8 | 6 | 5 |
| Engelsk | 172 | 5 | 67 | 14 | 6 | 0.1 | 3 | 3 | 1 |
| irsk | 257 | 1 | 81 | 6 | 5 | 0 | 2 | 1 | 1 |
| italienere | 99 | 2 | 44 | 3 | 4 | 0 | 13 | 18 | 8 |
| rumenere | 45 | 20 | 18 | 2 | 18 | 0 | 7 | 13 | 7 |
| ossetere | 359 | 1 | 7 | 0 | 0 | 1 | 16 | 67 | |
| armenere | 112 | 2 | 26 | 0 | 4 | 0 | 6 | 20 | 10 |
| grekere | 116 | 4 | 14 | 3 | 10 | 0 | 21 | 23 | 5 |
| tyrkere | 103 | 7 | 17 | 1 | 5 | 4 | 10 | 24 | 12 |
Spesielt bemerkelsesverdig er de 4 vanligste haplogruppene blant russere:
R1a1 47,0 %, N1c1 20,0 %, I2 10,6 %, I1 6,2 %
snakker for å si det enkelt: genetisk sammensetning russere langs de rette mannlige linjene til Y-kromosomet er som følger:
østeuropeere - 47 %
Baltikum - 20 %
Og to haplogrupper av originale europeere siden paleolitikum
Skandinaver - 6 %
Balkan - 11 %
Navnene er betingede og gitt i samsvar med de territorielle maksimumene europeisk underklader for haplogruppene R1a1, N1c1, I1 og I2. Det grunnleggende poenget er at det ikke var noen etterkommere av mongolene etter det to hundre år lange tatar-mongolske åket. Enten igjen, men et svært lite antall direkte genetiske arvinger fra slike forhold. Med disse ordene ønsker jeg ikke å stille spørsmål historiske kilder om mongolene i Rus', men bare for å ta hensyn til den antatte genetiske påvirkningen fra mongol-tatarene på russerne - den er ikke der, eller den er ubetydelig. Forresten, det er også et stort antall bærere i genomet til de bulgarske tatarene haprogrupper R1a1(ca. 30%) og N1c1(ca. 20%), men de er stort sett ikke-europeiske.
En annen viktig poeng, sørlige russere skiller seg ikke fra ukrainere innenfor feilmarginen, og nordrussere, som har en av de dominerende haplogruppene R1a1, har også en høyere prosentandel av haplogruppen N1c1. Men prosentandelen av N1c1-haplotyper er i gjennomsnitt 20 % hos russere.
Keisere. Nicholas 2
Den første kjente stamfaren til storhertughuset i Oldenburg var Egilmar, greve av Lerigau (d. 1108), nevnt i annalene for 1091.
Nicholas II viste seg å være bæreren av haplogruppen R1b1a2- en representant for den vesteuropeiske linjen, fra Holstein-Gottorp-dynastiet. Dette tyske dynastiet er preget av terminalsnippen U106, som er mest vanlig i nordvest-Europa på bosettingssteder for de germanske stammene. Det er ikke akkurat typisk russiske folk DNA-markør, men dens tilstedeværelse blant russere kan også være assosiert med tidlige kontakter mellom tyskere og slaver.
naturlige fyrster. Rurikovichi
Vladimir Monomakh og hans etterkommere, referert til som "Monomachis" tilhører haplogruppen N1c1-L550, som er utbredt i den sør-baltiske regionen (subclade L1025) og i Fennoscandia (subclade Y7795, Y9454, Y17113, Y17415, Y4338). Rurik-dynastiet er preget av terminalsnippen Y10931.
Noen av dem som historikere kaller Olgoviches (oppkalt etter Oleg Svyatoslavich - hovedrivalen til Vladimir Monomakh i den føydale kampen - og, som alle kilder forsikrer, hans fetter) er ikke relatert til Rurikovichs fra Monomashich-familien (i en direkte mannlig linje). Dette er etterkommerne til Yuri Tarussky
Russere, slaver, indoeuropeere og haplogruppene R1a, R1b, N1c, I1 og I2
I antikken, for rundt 8-9 årtusener siden, var det en språklig gruppe som la grunnlaget for den indoeuropeiske språkfamilien (i det innledende stadiet er dette mest sannsynlig haplogruppene R1a og R1b). Den indoeuropeiske familien inkluderer slike språklige grupper som indo-iranere (Sør-Asia), slaver og baltere (Øst-Europa), keltere (Vest-Europa), tyskere (Sentral-, Nord-Europa). Kanskje de også hadde felles genetiske forfedre, som for rundt 7 tusen år siden, som et resultat av migrasjoner, havnet i forskjellige deler av Eurasia, noen dro sørover og østover (R1a-Z93), og la grunnlaget for de indo-iranske folkene og språk (som i stor grad deltok i etnogenesen til de turkiske folkene), og en del forble på Europas territorium og la grunnlaget for dannelsen av mange europeiske folk (R1b-L51), inkludert slaverne og russere spesielt (Rla-Z283, Rlb-L51). På forskjellige stadier av dannelsen, allerede i antikken, var det kryss av migrasjonsstrømmer, noe som forårsaket tilstedeværelsen av et stort antall haplogrupper i alle europeiske etniske grupper.De slaviske språkene dukket opp fra den en gang forente gruppen av balto-slaviske språk (antagelig den arkeologiske kulturen til den sene Corded Ware). Ifølge beregningene til språkforskeren Starostin skjedde dette for rundt 3,3 årtusener siden. Periode fra det 5. århundre f.Kr til det 4.-5. århundre e.Kr kan betraktes som betinget protoslavisk, tk. Balterne og slaverne hadde allerede delt seg, men det var ingen slaver selv ennå, de vil dukke opp litt senere, på 4-600-tallet e.Kr. I det innledende stadiet av dannelsen av slaverne var sannsynligvis omtrent 80% haplogruppene R1a-Z280 og I2a-M423. I det innledende stadiet av dannelsen av balterne var sannsynligvis omtrent 80 % haplogrupper N1c-L1025 og R1a-Z92. Innflytelsen og skjæringspunktet mellom migrasjonene til balterne og slaverne var helt fra begynnelsen, fordi denne inndelingen i mange henseender er vilkårlig, og gjenspeiler generelt bare hovedtrenden, uten detaljer.
De iranske språkene er indoeuropeiske, og dateringen deres er som følger - de eldste, fra det andre årtusen f.Kr. til det 4. århundre f.Kr., det midterste - fra det 4. århundre f.Kr. til 900-tallet e.Kr., og en ny - fra 900-tallet e.Kr. Inntil nå. Det vil si at de eldste iranske språkene dukker opp etter avgangen til en del av stammene som snakket indoeuropeiske språk fra Sentral-Asia til India og Iran. Deres viktigste haplogrupper var sannsynligvis R1a-Z93, J2a, G2a3. Den vestiranske språkgruppen dukket opp senere, rundt det 5. århundre f.Kr.
Dermed ble indo-arierne, kelterne, tyskerne og slaverne i akademisk vitenskap indoeuropeere, dette begrepet er det mest passende for en så stor og mangfoldig gruppe. Dette er helt riktig. I det genetiske aspektet er heterogeniteten til indoeuropeerne slående både i Y-haplogrupper og i autosomer. Indo-iranerne preges i større grad av den vestasiatiske genetiske påvirkningen av BMAC.
I følge de indiske vedaene var det indo-arierne som kom til India (Sør-Asia) fra nord (fra Sentral-Asia), og det var deres salmer og legender som dannet grunnlaget for de indiske vedaene. Og la oss fortsette videre, la oss berøre lingvistikk, fordi dette russiske språket (og dets beslektede baltiske språk, for eksempel litauisk som en del av det en gang eksisterende balto-slaviske språksamfunnet) er relativt nær sanskrit sammen med keltiske, germanske og andre språk av den store indoeuropeiske familien. Men i den genetiske planen var indo-arierne allerede i større grad vestasiater, da de nærmet seg India, ble også vedoid-innflytelsen intensivert.
Så det ble klart det haplogruppe R1a i DNA-slektsforskning er dette en vanlig haplogruppe for en del av slaverne, en del av tyrkerne og en del av indo-arierne (fordi det naturlig nok var representanter for andre haplogrupper i deres miljø), en del haplogruppe R1a1 under migrasjoner langs den russiske sletten ble de en del av de finsk-ugriske folkene, for eksempel mordovierne (Erzya og Moksha). En del av stammene (for haplogruppe R1a1 dette er en undergruppe av Z93) under migrasjoner brakte de dette indoeuropeiske språket til India og Iran for rundt 3500 år siden, det vil si i midten av det 2. årtusen f.Kr. I India, ved arbeidet til den store Panini, ble det omgjort til sanskrit i midten av det 1. årtusen f.Kr., og i Persia-Iran ble de ariske språkene grunnlaget for en gruppe iranske språk, hvorav de eldste dateres tilbake til det 2. årtusen f.Kr. Disse dataene er bekreftet: DNA slektsforskning og lingvistikk er korrelert her.
Stor del haplogrupper R1a1-Z93 selv i antikken sluttet de seg til de tyrkiske etniske gruppene og markerer i dag tyrkernes migrasjon på mange måter, noe som ikke er overraskende med tanke på antikken haplogruppe R1a1 mens representantene haplogrupper R1a1-Z280 var en del av de finsk-ugriske stammene, men under bosettingen av de slaviske kolonistene ble mange av dem assimilert av slaverne, men selv nå blant mange folk, for eksempel, er Erzya fortsatt den dominerende haplogruppen R1a1-Z280.
Alle disse nye dataene var i stand til å gi oss DNA slektsforskning, spesielt de omtrentlige datoene for migrasjonene til haplogruppebærere på territoriet til den moderne russiske sletten og Sentral-Asia i forhistorisk tid.
Så forskere til alle slaver, keltere, tyskere, etc. ga navnet på indoeuropeerne, noe som er sant fra et språkvitenskapelig synspunkt.
Hvor kom disse indoeuropeerne fra? Faktisk var det indoeuropeiske språk lenge før migrasjonene til India og Iran, over hele den russiske sletten og så langt som til Balkan i sør, og så langt som til Pyreneene i vest. Senere ble språket spredt til Sør-Asia – både til Iran og India. Men i genetiske termer er korrelasjonene mye mindre.
"Det eneste berettigede og aksepterte på det nåværende tidspunkt i vitenskapen er bruken av begrepet "ariere" bare i forhold til stammene og folkene som snakket de indo-iranske språkene."
Så i hvilken retning gikk den indoeuropeiske strømmen - mot vest, til Europa, eller omvendt, mot øst? I følge noen estimater er den indoeuropeiske språkfamilien rundt 8500 år gammel. Indoeuropeernes forfedres hjem er ennå ikke bestemt, men ifølge en av versjonene kan det være Svartehavsregionen - sørlig eller nordlig. I India, som vi allerede vet, ble det indo-ariske språket introdusert for rundt 3500 år siden, antagelig fra territoriet til Sentral-Asia, og arierne selv var en gruppe med forskjellige genetiske Y-linjer, som R1a1-L657, G2a, J2a, J2b, H, osv.
Haplogruppe R1a1 i Vest- og Sør-Europa
Analyse av 67 markørhaplotyper haplogruppe R1a1 fra alle europeiske land gjorde det mulig å bestemme den omtrentlige migrasjonsveien til R1a1-forfedre i retning Vest-Europa. Og beregninger viste at nesten i hele Europa, fra Island i nord til Hellas i sør, var den felles stamfaren til haplogruppen R1a1 en for omtrent 7000 år siden! Med andre ord, etterkommerne, som et stafettløp, videreførte sine haplotyper til sine egne etterkommere fra generasjon til generasjon, og spredte seg i prosessen med migrasjon fra det samme historiske stedet - som viste seg å være antagelig Ural- eller Svartehavslavlandet . På et moderne kart er dette land hovedsakelig i Øst- og Sentral-Europa - Polen, Hviterussland, Ukraina, Russland. Men rekkevidden av eldre haplotyper av haplogruppen R1a1 fører mot øst - til Sibir. Og levetiden til den første stamfaren, som er indikert av de eldste, mest muterte haplotypene, er 7,5 tusen år siden. På den tiden var det ingen slaver, ingen tyskere, ingen keltere.Ulempen med metoden
Hvis du gjorde testen, og den gledet deg veldig, så skynder jeg meg å hente inn min tjæreøse. Ja, Y-kromosomet overføres fra far til sønn praktisk talt uendret, men faktisk genetisk nyttig informasjon ikke i den, i andre kromosompar er det mye flere gener.
Og disse andre 22 stokkes på en veldig tilfeldig måte, uten spor av slik blanding igjen på Y.
Forestill deg. Angelsaksiske sjømenn fanget negerstaten. Kvinner tas ikke med på slike turer, og man må etablere kontakt med lokalbefolkningen. Hva er mulighetene?
1) Angelsakserne har barn fra svarte kvinner, men nasjonaliteten overføres kun til gutter. I dette tilfellet vil Y-kromosomet overføres til det europeiske, men andelen av faktisk signifikante europeiske gener vil avta. Den første generasjonen vil være halvsvarte og det tidligere "aristokratiet" i et slikt tilfelle vil raskt oppløses, selv om Y vil være fra dette etnisk gruppe. Det vil bare ikke gi mye mening. Kanskje skjedde noe lignende med finnene og indianerne. Yakutene og finnene har den høyeste prosentandelen av N1c1-haplogruppen som er karakteristisk for dem, men genetisk sett er dette helt forskjellige folkeslag med forskjellige undergrupper av N1c1-haplogruppen med sin egen unike historie, atskilt for mer enn 6 årtusener siden. Og omvendt, indianere - med en høy prosentandel haplogruppe R1a1 genetisk har de svært lite til felles med de europeiske representantene for denne haplogruppen, tk. også ulike underklader med sin egen historie, skilt ut for mer enn 6 årtusener siden.
2) Indo-ariere er fornøyde kastesystem. Den første generasjonen vil også være semi-negro, men hvis aristokratiet bare blander seg i hverandre, vil prosentandelen av original genetikk flyte rundt 50%. Men i praksis vil ekteskap hovedsakelig være med lokale kvinner, og enda mer vil det være umulig å få den opprinnelige genpoolen til erobrerne. Og dette skjedde i jordens historie. øvre kaster Indere fra 20 % til 72 % har haplogruppe R1a1(gjennomsnittlig 43%), men genetisk har de svært lite til felles med europeiske eller turkiske representanter for samme haplogruppe R1a1, og igjen årsaken er de ulike underkladdene med sin egen spesielle historie.
En lignende situasjon skjedde sannsynligvis i Kamerun, et sentralafrikansk land hvor Y er opptil 95 % vanlig. haplogruppe R1b-V88, men samtidig blant en typisk antropologisk afrikansk negerbefolkning.
Det kan konkluderes med at tilstedeværelsen av en markør og en haplogruppe er en viktig betingelse for å fastslå nasjonalitet, men ikke tilstrekkelig. For å bestemme den nasjonale territorielle opprinnelsen til en person, har Family Tree DNA en autosomal test kalt Family Finder
Alexey Zorrin 
Forskere har nylig kommet i nærheten av å tyde den menneskelige genetiske koden. På mange måter gjorde dette det mulig å ta et nytt blikk på historien til den russiske etnoen, som viste seg å være mer eldgammel og ikke så homogen som tidligere antatt.
I dypet av århundrer
Det menneskelige genomet er en variabel ting. I løpet av menneskehetens utvikling har dens haplogrupper gjennomgått mutasjoner mer enn én gang. I dag har forskere allerede lært å bestemme det omtrentlige tidspunktet da denne eller den mutasjonen oppsto. Så, amerikanske genetikere fant at en av disse mutasjonene skjedde for rundt 4500 år siden på den sentralrussiske sletten. Det ble født en gutt som hadde et annet sett med nukleotider enn faren - han ble tildelt den genetiske klassifiseringen R1a1, som oppsto i stedet for den faderlige R1a.
Denne mutasjonen, i motsetning til mange andre, viste seg å være levedyktig. R1a1-slekten overlevde ikke bare, men slo seg også ned i en betydelig del av det eurasiske kontinentet. For tiden er omtrent 70 % av den mannlige befolkningen i Russland, Hviterussland og Ukraina bærere av R1a1-haplogruppen, og i gamle russiske byer når dette tallet 80 %. Dermed fungerer R1a1 som en slags markør for den russiske etniske gruppen. Det viser seg at blodet til en eldgammel gutt som levde i den sene neolitiske epoken flyter i venene til de fleste menn i det moderne Russland.
Omtrent 500 år etter fødselen av R1a1-haplogruppen, spredte migrasjonsstrømmene til dens representanter seg mot øst - utover Ural, til sør - til Hindustan og til vest - til det moderne territoriet. europeiske land. Det faktum at innbyggerne på den sentralrussiske sletten gikk langt utenfor sin opprinnelige rekkevidde bekreftes også av arkeologer. Analyse av beinrestene etter begravelser i Altai i det 1. årtusen f.Kr. e. viste at det i tillegg til mongoloidene også bodde uttalte kaukasiere der.
Det er ingen tatar
I en av utgavene av den populærvitenskapelige publikasjonen The American Journal of Human Genetics ble det publisert en artikkel om forskningen til det russisk-estiske teamet av forskere i genpoolen til det russiske folket. Funnene til forskerne var ganske uventede. For det første: den russiske etnoen er heterogen i sin genetiske natur. Den ene delen av russerne som bor i de sentrale og sørlige regionene av landet er nær de slaviske nabofolkene, den andre delen - nord i Russland - er genetisk nært beslektet med de finsk-ugriske folkene.
Den neste konklusjonen er mer interessant. Forskere har ikke klart å oppdage det beryktede asiatiske elementet i det russiske genomet. Det er ikke noe tatarisk-mongolsk sett med gener i noen merkbar mengde i noen av de russiske populasjonene. Det viser seg at det stabile uttrykket "Scratch a Russian - you will find a Tatar" er feil.
Professor Oleg Balanovsky, leder for laboratoriet for genomisk geografi ved Institute of General Genetics ved det russiske vitenskapsakademiet, anser den russiske genpoolen som "nesten fullstendig europeisk", og kaller dens forskjeller fra den sentralasiatiske "virkelig store", som hvis de er to forskjellige verdener.
Akademiker Konstantin Skryabin, leder for den genomiske retningen ved Kurchatov-instituttet, er enig med Balanovsky. Han sier følgende: "Vi fant ikke merkbare tatariske introduksjoner i det russiske genomet, noe som tilbakeviser teorier om den destruktive innflytelsen fra det mongolske åket." I tillegg er sibirere, ifølge forskeren, genetisk identiske med de gamle troende - de har det samme "russiske genomet".
Forskerne trekker også frem en liten forskjell i genotypen mellom russerne på den ene siden og de slaviske nabofolkene – ukrainere, hviterussere og polakker – på den andre. Forskjellen mellom de sørlige og vestlige slaverne fra innbyggerne i det russiske nord er mer uttalt.
Spesielle markører
Ifølge antropolog Vasily Deryabin har den russiske genotypen også sine egne klare fysiologiske markører. En av dem er overvekten av lyse nyanser av øyne på russere: grå, blå, gråblå, blå. Vi har 45 prosent av dem Vest-Europa mindre - omtrent 35 prosent. Mange blant russere og lyshårede. Ifølge antropologer er russere med naturlig svart hårfarge ikke mer enn 5 prosent. I Vest-Europa er sjansen for å møte en svarthåret 45 %.
I motsetning til det mange tror, er det ikke så mange snub-nese-mennesker blant russere - ca 7%, i ca 75% av tilfellene er nesen rett. Også blant russere finnes ikke epicanthus - en fold som er typisk for representanter for mongoloide folk i den indre øyekroken.
Den russiske etniske gruppen er preget av overvekt av I og II blodgrupper, blant jøder er for eksempel gruppe IV mer vanlig. Biokjemiske studier viste også at i blodet til russere, så vel som andre europeiske folk, er det et spesielt gen PH-c, men det er fraværende hos mongoloidene.
Nordlendingene er nærmere
Forskningsinstitutt for molekylær genetikk ved det russiske vitenskapsakademiet og Institutt for antropologi. D.N. Anuchin Moscow State University gjennomførte en dybdestudie av genpoolen til det russiske folket, der en forskjell i genotypen mellom russerne og våre nordlige naboer, finnene, ble etablert - det utgjorde tretti konvensjonelle enheter. Men de genetiske forskjellene mellom den russiske etniske gruppen og de finsk-ugriske folkene (Mordovians, Mari, Veps, Karelians, Komi-Zyryans, Izhors), som tradisjonelt bodde nord i landet vårt, tilsvarer bare tre enheter.
Forskere snakker ikke bare om russernes genetiske enhet med de finsk-ugriske folkene, men om deres felles opprinnelse. Dessuten er den spesifikke strukturen til Y-kromosomene til disse etniske gruppene stort sett identisk med folkene i Hindustan. Men dette er ikke overraskende, gitt retningen for bosettingen av de genetiske forfedrene til det russiske folket.
Menneskelige haplogrupper overføres gjennom direkte mannlige og kvinnelige linjer. Men for genetikken til både menn og kvinner er informasjonen som er lagret i autosomene til DNA ansvarlig. Autosomer er de første 22 parene av menneskelige kromosomer som overføres fra begge foreldrene etter å ha krysset over, en prosess med rekombinasjon. Dermed overføres omtrent halvparten av den genetiske informasjonen fra far og mor til etterkommere.I denne studien mer enn 80 000 autosomale klipp er brukt, fiducial poeng er svært en høy oppløsning, som gjør det mulig å fange opp selv relativt små påvirkninger på genetisk nivå i hoveddelen av menneskene. Sammenlignende analysedata er hentet fra en åpen studie av V.Verenich, spesialist i komparativ analyse genetiske komponenter. Selve de genetiske kalkulatorene er på GedMatch-tjenesten, og lar hvem som helst finne ut sin komparative posisjon på den genetiske grafen. For å gjøre dette er det nok å ha resultatene av en autosomal test fra FTDNA, eller 23andMe. Kart leveres på slutten av studien. geografisk fordeling og frekvensmaksima for store autosomale komponenter fra MDLP World-22-prosjektet.
Grafene nedenfor viser hovedkomponentene og deres gjennomsnittlige prosentandel for hver av populasjonene. På én linje - prosentvis fordeling for én populasjon. Hver inndeling (vertikal strek) tilsvarer 10 %, og navnene på de autosomale komponentene er i samme rekkefølge fra venstre til høyre som i forklaringen fra topp til bunn. Jo mer lik den prosentvise sammensetningen av vanlig genetikk i forskjellige folkeslag, jo mer lik figuren i grafen ovenfor ser ut. Så la oss komme i gang...
Genetikk til tyskere, litauere, russere, svensker, finner osv.
Denne grafen viser de viktigste genetiske komponentene for europeiske folk og på linje med nedgangen i den østeuropeiske komponenten (nordøsteuropeisk) i ulike populasjoner. Som du kan se, er alle europeiske folk ganske forskjellige i genetiske termer, og med genetiske komponenter i sitt sett av samme opprinnelse, er de likevel i svært forskjellige prosenter. For alle slaver og baltere generelt er en av de mest betydningsfulle denne komponenten i Øst-Europa, som er på sitt maksimum blant litauere og hviterussere. Sannsynligvis fra tiden til den arkeologiske "Corded Ware-kulturen" var territoriet til disse landene opprinnelsessenteret for denne komponenten. Den er representert med mer enn 80 % blant litauere, og bare 20 % blant italienere.Den lilla fargen indikerer Atlanto-Middelhavskomponenten, og den øker når den beveger seg fra nordøst til sørvest. Så blant finnene når den et gjennomsnitt på 15 %, og blant italienerne 40 %. Resten av komponentene er mindre uttalt.
Genetikk til russiske ukrainere hviterussere

Denne grafen viser østslaverne - Russere, hviterussere, ukrainere. Oppmerksomheten rettes mot likheten mellom de genetiske mønstrene til de tre listede folkene, og innenfor feilmarginen skiller de seg svært lite - ukrainere og sør-russere har en liten økning i den vestasiatiske komponenten, og nord-russere har en liten økning i en av de sibirske komponentene, betinget kalt samojedisk, og en økning av komponentene i mesolitikum i Europa til omtrent 10%, som ifølge sistnevnte indikator bringer dem nærmere den tysktalende befolkningen i Skandinavia - svenskene.
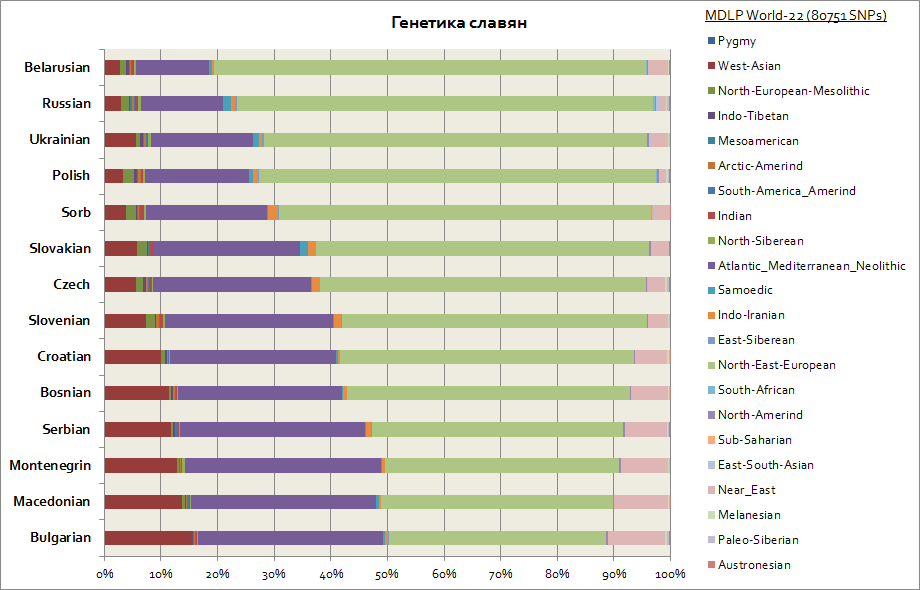
Dette diagrammet viser alle slaverne, inkludert de vestlige - polakker og tsjekkere, så vel som de sørlige - serbere, bulgarere, makedonere, etc.
Alle slaver har 2 hovedkomponenter, disse er østeuropeiske og atlantiske middelhavskomponenter. Den første er på maksimum for hviterussere, og den andre for alle sørslavere - serbere, makedonere, bulgarere. Den østeuropeiske komponenten er mer primær i opprinnelse blant slaverne, og Atlanto-Middelhavskomponenten er mer ervervet etter hvert som slaverne migrerte til Balkan. Vest-ukrainere og slovakker har en liten økning i den samojediske komponenten i forhold til nabolandet Slaviske folk- Hviterussere, tsjekkere, polakker; dette er sannsynligvis det genetiske sporet etter middelalderens migrasjoner av hunnerne og ugrierne til Sentral-Europa.
Genetikk til slaver, russere og tatarer, tyskere, kaukasiere, jøder, etc.
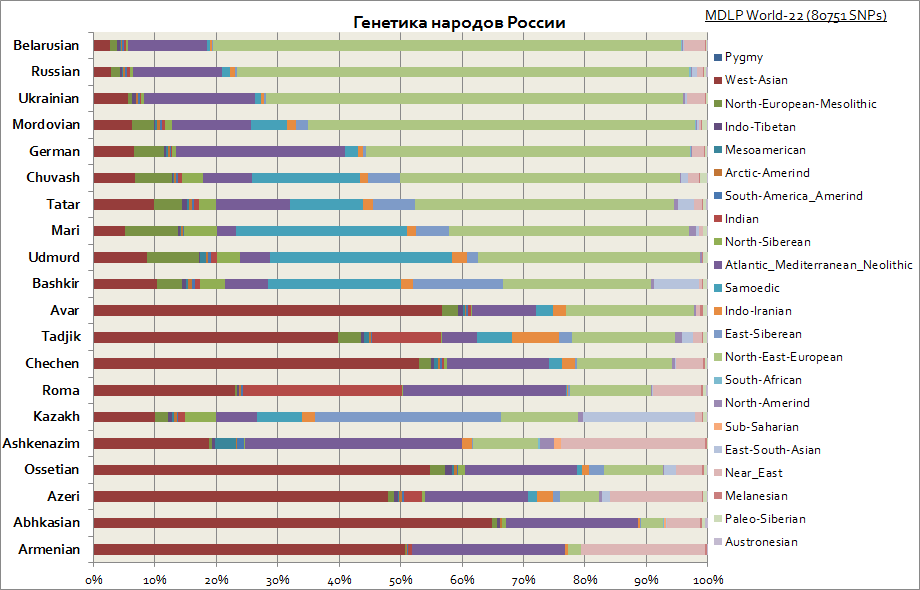
Denne grafen viser de forskjellige opprinnelsene blant folkene i Russland. Som man kan se, blant slaverne, er hovedkomponenten den østeuropeiske, mens blant folkene i Volga-regionen øker andelen av de sibirske komponentene. Mens for kaukasiere er den vestasiatiske komponenten, Middelhavet og Midtøsten de mest karakteristiske.
Genetikk til finnene, ugriere, udmurtere, ungarere, samer, etc.
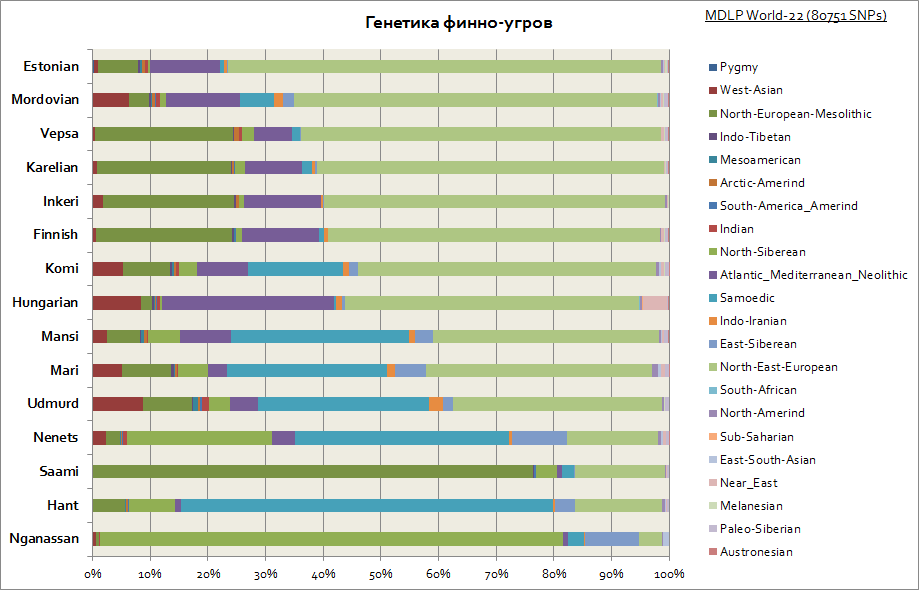
Som man kan se er finnene, vepsianerne og karelerne preget av lignende genetisk opprinnelse med slaverne. De har også den største østeuropeiske komponenten, og avtar nærmere Ural- og Volga-regionen, med en økning i de sibirske komponentene i denne regionen. Alle de finsk-ugriske folkene har også en betydelig del av Europas mesolittiske alder, som når nesten 80 % blant samene og er assosiert med de pre-indoeuropeiske og pre-neolittiske befolkningene i Europa. For ungarerne som helhet er et sett av disse genetiske komponentene karakteristiske, som for andre populasjoner i Karpatene og Sentral-Europa.
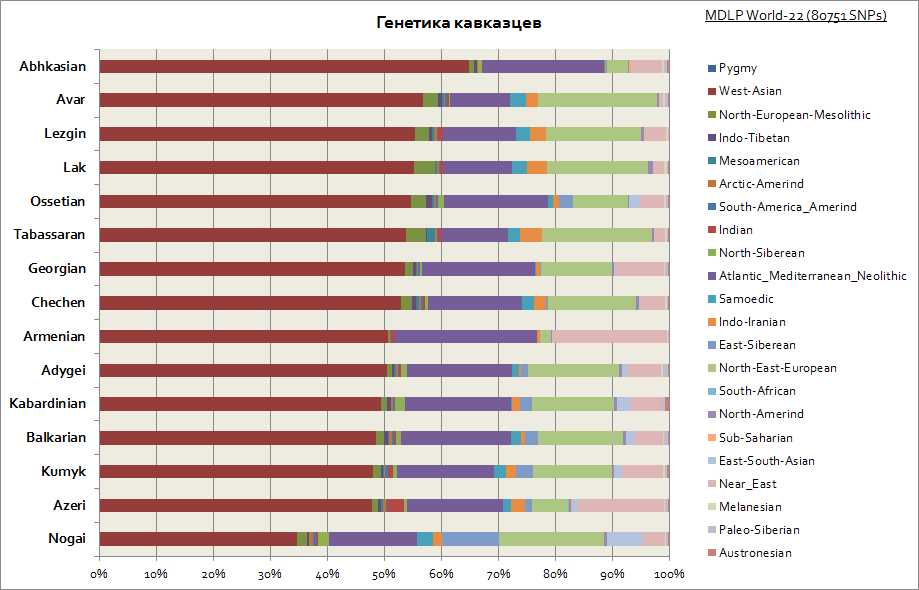
Som man kan se, er hele Kaukasus preget av en relativt lik genetisk opprinnelse - dette er en stor andel av den vestasiatiske komponenten og Middelhavet. Bare Nogais skiller seg litt ut – de har en økt andel sibirske komponenter.

Som man kan se i Ashkenazim og Sephardim, er det en høy frekvens av de vestasiatiske, Atlanto-Middelhavs- og Midtøsten-komponentene. Samtidig har Ashkenazim en liten økning i den sibirske komponenten, noe som sannsynligvis skyldes Khazar-arven og en økning på opptil 30% av den østeuropeiske komponenten, som ifølge denne indikatoren bringer dem nærmere land i Sør-Europa.
Bare etiopiske jøder og indiske jøder er spesielt slått ut av deres "selskap". De førstnevnte har en høy andel av Afrika sør for Sahara (opptil 40 %), mens sistnevnte har en andel av den sørasiatiske genetiske komponenten, betinget kalt indisk (opptil 50 %).
Genetikk til tatarer, bashkirer, aserbajdsjanere, tjuvasjer, etc.
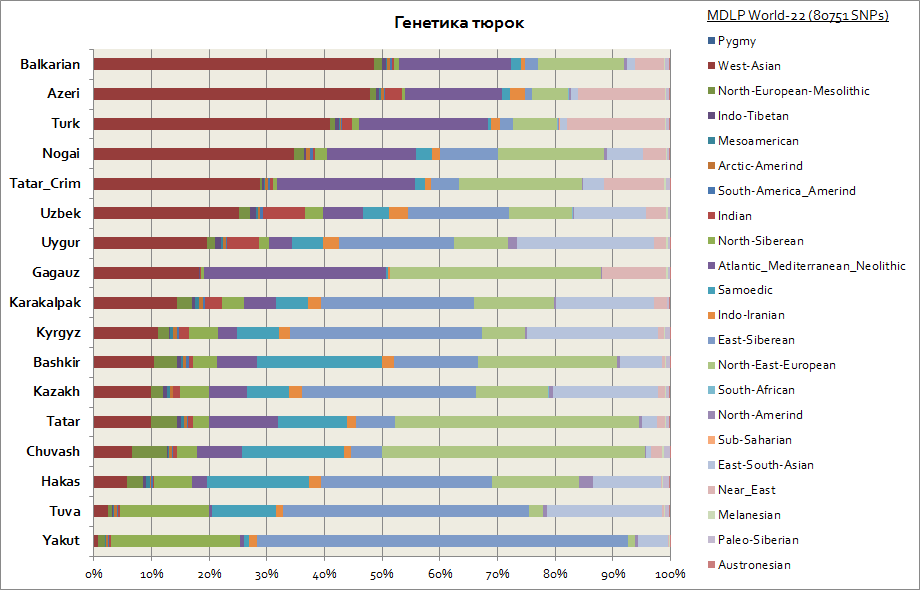
Tyrkerne i genetiske termer viste seg å være en av de mest heterogene etniske gruppene, fordi deres genetiske komponenter er betydelig forskjellige. Så, gitt at det primære hjemlandet til tyrkerne er Sibir, så har slike folk som yakutene, tuvanene, khakassene beholdt den østsibirske autosomale komponenten i den største prosentandelen, som når fra 30 til 65% i dem. Denne genetiske komponenten er også den viktigste blant kirgiserne og kasakherne. De resterende komponentene bringer tyrkerne nærmere folkene fra bostedsregionene. Så for yakutene og tuvanene er dette de nordsibirske og samojed-komponentene. Totalt disse 3 sibirske komponentene blant yakutene utgjør de opptil 90 %, blant tuvanene opptil 70 %, med en økning til 20 % av den øst-sør-asiatiske komponenten, som i større grad er assosiert med migrasjonsstrømmer av befolkningen øst Asia. For basjkirene er andelen av de 3 sibirske komponentene opptil 45 %, og den sørøstasiatiske komponenten er opptil 10 %. Tatarer har dataene til 3 sibirske genetiske komponenter i gjennomsnitt fra 25 til 50%. Samtidig er andelen komponenter som er karakteristiske for den kaukasoide befolkningen blant bashkirene opptil 45%, og blant tatarene i gjennomsnitt fra 50 til 70%. Genetikken til aserbajdsjanere og tyrkere skiller seg praktisk talt ikke innenfor feilmarginen; de, i likhet med andre folkeslag i Kaukasus og Transkaukasia, har en betydelig tilstedeværelse av den vestasiatiske komponenten (når 50%) og Atlanto-Middelhavskomponenten (opp til 20 % i gjennomsnitt). Andelen av 3 sibirske komponenter er representert av aserbajdsjanere, tyrkere og balkarer - på nivået 3-7%.
Konklusjon
Genetikken til folk har ikke en direkte og signifikant korrelasjon med fordelingen av språkfamilier, eller med prosentandelen av uniparentale markører - Y-DNA og mt-DNA haplogrupper, representert i en bestemt populasjon. Den største korrelasjonen kan spores etter det territorialgeografiske prinsippet. Dermed andelen av de sibirske komponentene, som generelt er karakteristiske for Mongoloid rase avtar gradvis fra øst til vest, og andelen komponenter som er karakteristiske for den kaukasoide rasen øker tilsvarende. I grenseområdene langs linjen fra nord i Ural til Sentral Asia omtrent likt forhold. I regionene øst for Baikalsjøen er de genetiske komponentene som er karakteristiske for den store kaukasiske rasen praktisk talt ikke representert, mens samtidig, i regionene vest for Pechora-Volga-linjen, forsvinner de sibirske komponentene som er karakteristiske for den store mongoloide rasen. .Spredningen av den østeuropeiske genetiske komponenten til Sibir skjedde i stor grad allerede i bronsealderen (kulturene i Andronov-sirkelen), selv om individuelle topper i det ytterste øst av Sibir blant tsjuktsjene allerede kan være assosiert med russiske migrasjoner i 17. århundre.
Andelen av komponenten sør for Sahara, karakteristisk for den negroide rasen, er fordelt over hele Afrika - opp til det sørlige Middelhavet og den nordlige grensen til det afrikanske kontinentet, og når et maksimum i sin ekvatoriale del, og finnes praktisk talt aldri utenfor den; en lys bakgrunn er fordelt på den arabiske halvøy og den sørlige delen av det iranske platået.
Geografi av genetiske komponenter
Alexey Zorrin
Prosjekt



